
Abstract
Nonalcoholic fatty liver disease (NAFLD) progresses to nonalcoholic steatohepatitis, ultimately leading to cirrhosis and liver cancer. It is important to prevent this progression during the initial stages of hepatic fatty degeneration. Maltitol is a polyol produced by the hydrogenation of maltose. We investigated the efficacy of maltitol for treating hepatic fatty degeneration in C57BL/6 male mice using a high-fat diet model. Intake of 5.0% maltitol for 8 weeks significantly suppressed weight gain, hepatic fatty degeneration, hyperglycemia, and hypercholesterolemia. With maltitol intake, sterol regulatory element-binding protein 1c (SREBP1c) mRNA expression was significantly decreased, and farnesoid X receptor (FXR), peroxisome proliferator-activated receptor α (PPARα), and hydroxymethylglutaryl-Co reductase expressions were significantly higher in the liver. The increase in SREBP1c and suppression of FXR and PPARα expressions are correlated with NAFLD. Our results suggest that maltitol may prevent steatosis of NAFLD with a high-fat diet.
Introduction
N
Maltitol is a polyol produced by the hydrogenation of maltose 7 and has a low caloric value and low glycemic index. 8,9 Furthermore, maltitol has sweetness and satiety-inducing effects equivalent to sugar and is currently used as a sugar substitute by diabetic patients.
In this study, we investigated the efficacy of supplementing a high-fat diet with maltitol for 8 weeks using an early-stage model of NAFLD.
Materials and Methods
Mice
Eight-week-old C57BL/6 male mice were divided into three groups (n=4). The “normal” group received a normal diet (MF; Oriental Yeast Co., Tokyo, Japan)+distilled water. The “control” group received a high-fat diet (High Fat Diet 32; CLEA Japan, Tokyo, Japan)+distilled water. The “maltitol” group received the high-fat diet+5.0% maltitol (Ueno Fine Chemical Industry, Osaka, Japan). In consideration of the results of preliminary studies, the concentration of maltitol was determined. After 8 weeks of treatment, mice were anaesthetized using isoflurane and sacrificed to collect blood and organ samples by cardiocentesis. The inguinal fat pad was weighed to represent subcutaneous fat tissue weight. The concentration of maltitol was decided by our preliminary study, since no adverse events such as diarrhea were observed by the intake of 5.0% maltitol.
Ethics
The experimental protocol was approved by the Ethics Review Committee for Animal Experimentation of Osaka University Graduate School of Medicine and was carried out in accordance with the approved guidelines.
Hepatic fatty degeneration index
Hepatocytes were stained using hematoxylin and eosin and three images per hepatocyte sample were taken, with the exception of the blood vessel region, using a BZ-9000 fluorescence microscope (Keyence Corporation, Osaka, Japan). The vacuolar degeneration region was then selected using Dynamic Cell Count software (Keyence Corporation) and the glycogen degeneration area was identified. Subsequently, the glycogen degeneration area pixels were counted. The sum of the counted pixels was defined as the hepatic fatty degeneration index.
Measurement of hepatic triglyceride
Liver triglyceride (TG) content was determined as described by Chandler et al. 10 Briefly, 0.5 g of hepatic tissue was homogenized at 4°C in 3.0 mL PBS. Aliquots, 200 μL, of the homogenates were transferred to 15-mL screw-capped glass tubes containing 7.5 mL of a mixture of CHCl3-MeOH (2:1; v/v) and mixed vigorously for 20 sec. One milliliter of ddH2O was then added, and the resulting suspension was mixed vigorously for 15 sec then centrifuged at 800 g for 5 min at room temperature. The chloroform–methanol layer was removed, placed in a 13×100 mm test tube, and evaporated to dryness under nitrogen at 60°C. The lipid residue was resuspended in 200 μL 1% Triton X-100 in absolute EtOH, and the TG concentrations were determined using the Triglyceride E test kit (Wako Pure Chemical Industries, Osaka, Japan).
Plasma analysis
Plasma was isolated by centrifugation at 1000 g for 10 min and analyzed for aspartate aminotransferase (AST), alanine aminotransferase (ALT), total cholesterol, TG (Wako Pure Chemical Industries), glucose (Sanwa Kagaku Kenkyusyo, Kumamoto, Japan), and insulin (Morinaga Institute of Biological Science, Yokohama, Japan).
Real-time polymerase chain reaction
The total RNA was isolated from the liver, and the first-strand cDNA was synthesized using the ReverTra Ace-α First Strand cDNA Synthesis Kit (Toyobo, Osaka, Japan). Polymerase chain reaction amplification was performed using a LightCycler system (Roche Diagnostics, Tokyo, Japan). Primer sequences were as follows: sterol regulatory element-binding protein 1c (SREBP1c), forward: 5′-GGAGCCATGGATTGCACATT-3′, reverse: 5′-GCTTCCAGAGAGGAGGCCAG-3′; farnesoid X receptor (FXR), forward: 5′-TGTGAGGGCTGCAAAGGTTT-3′, reverse: 5′-ACATCCCCATCTCTCTGCAC-3′; peroxisome proliferator-activated receptor α (PPARα), forward: 5′-TGGGGATGAAGAGGGCTGAG-3′, reverse: 5′-GGGGACTGCCGTTGTCTGT-3′; hydroxymethylglutaryl-Co reductase (HMG-CoAR), forward: 5′-AAGCCGTCATTCCAGCCAAGGTG-3′, reverse: 5′-AATGTTTGCTGCGTGGGCGTTG-3′; fatty acid synthase (FAS): 5′-TCCTGGAACGAGAACACGATCT-3′, reverse: 5′-GAGACGTGTCACTCCTGGACTTG-3′; stearoryl CoA desaturase (SCD-1): 5′-GATAGAGCAAGTCCCCGTTG-3′, reverse: 5′-CCTGCATTAACCCCCTTCAC-3′; and β-actin, forward: 5′-TACCACAGGCATTGTGATGG-3′, reverse: 5′-TTTGATGTCACGCACGATTT-3′. SYBR Green dye (LightCycler DNA Master SYBR Green I; Roche Diagnostics) was used to detect cDNAs. Relative quantification of the signals was performed by normalizing their signals relative to β-actin signals.
Statistical analysis
The results are represented as the mean±SD. The comparison between groups was made by ANOVA, and the significance was analyzed by the Dunnett's test. Differences of P<.05 were considered statistically significant.
Results
Body and organ weight
The body weight of the maltitol group was significantly lower compared with the control group (Fig. 1a). The maltitol group tended to have a lower liver weight than the control group (Fig. 1b). No significant differences were observed between the two groups with respect to the weight of subcutaneous fat and epididymal fat (Fig. 1b). Diarrhea is one of the adverse events of maltitol. 11,12 However, 5.0% maltitol did not induce diarrhea as was confirmed in our preliminary study. Moreover, no differences were observed in fecal TG or in energy intake between the control and maltitol groups (data not shown).

The control group had a significantly higher weight gain than the normal group, but the 5.0% maltitol group had significantly lower weight gain than the control group
Hepatic fatty degeneration
The maltitol group showed significantly lower pixel counts in the region of hepatic vacuolar degeneration and hepatic TG concentration compared to the control group, suggesting that the intake of maltitol suppressed the hepatic fatty degeneration (Fig. 1c).
Plasma analysis
Plasma glucose, cholesterol, AST, and insulin levels were significantly lower in the maltitol group than in the control group. Moreover, the maltitol group tended to have lower ALT levels than the control group (Fig. 2). No differences were observed in the TG levels between the two groups (data not shown).
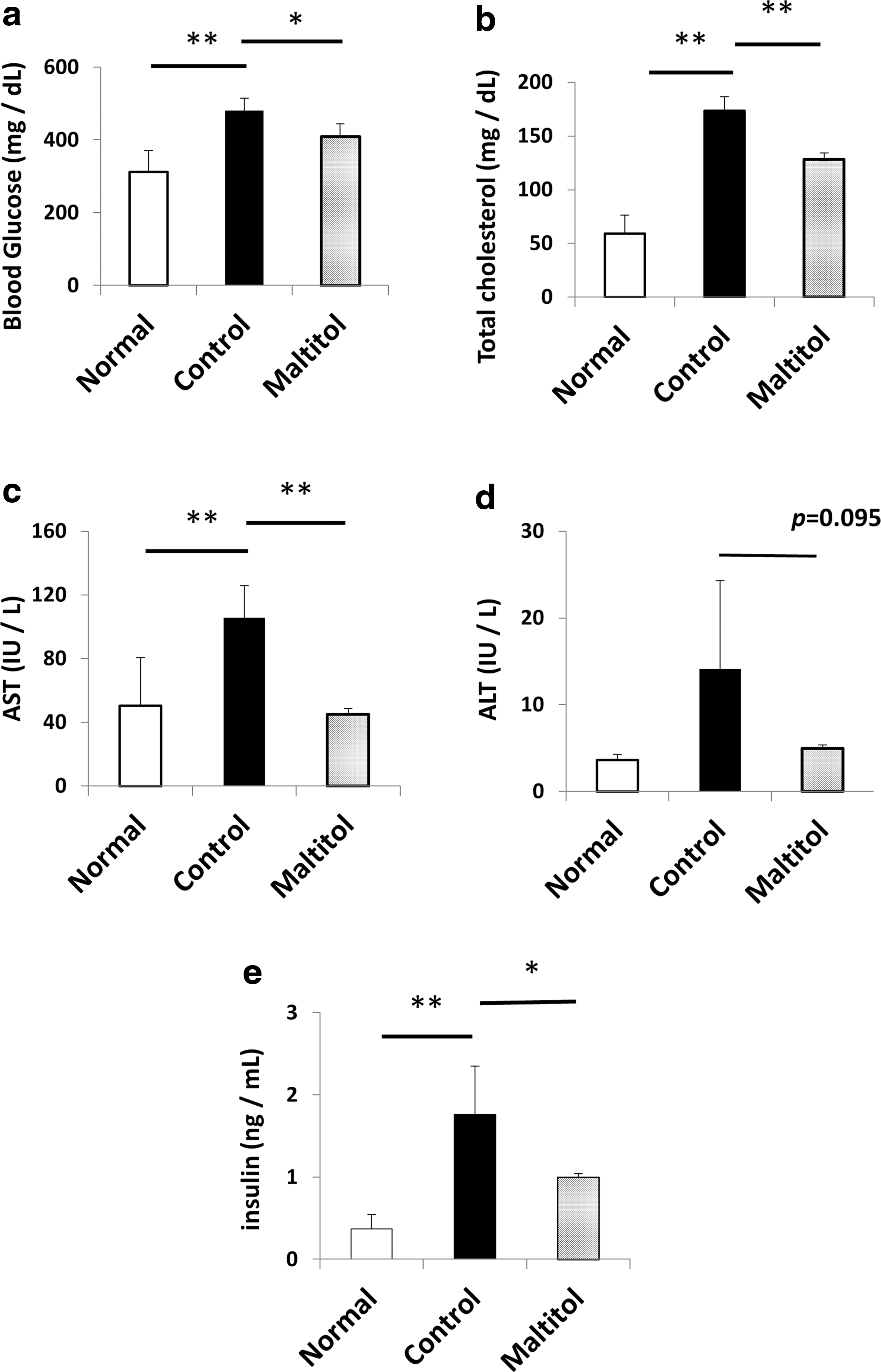
Plasma analysis. Glucose
The expression of fatty liver related genes in liver
SREBP1c is a transcription factor controlling the expression of genes involved in the metabolism of fatty acids and TG in the liver. 13 In the present study, the hepatic SREBP1c expression was significantly higher in the control group than in the normal and maltitol groups (Fig. 3a). FXR is a nuclear receptor activated by the bile acids, chenodeoxycholic acid, cholic acid, deoxycholic acid, and lithocholic acid, 14 –16 regulating the production and excretion of bile acids and protecting hepatocytes from bile acid toxicity. It has been reported that the SREBP1c expression is downregulated by the activation of FXR. 17 In this study, the FXR expression in the maltitol group was significantly higher than in the control group (Fig. 3b). PPARα regulates the proteins and enzymes involved in the metabolism of fatty acids in the liver 18 ; the SREBP1c expression in the liver is directly regulated by PPARα. 19 Also, the expression PPARα was enhanced by FXR. 20 Similar to FXR, maltitol increased the expression of PPARα, whereas the normal and control groups did not undergo such an increase (Fig. 3c). In addition, the expression of HMG-CoAR was significantly higher in the maltitol group than in the control group (Fig. 3d). There were no differences in the expression of FAS and SCD-1 among these groups (Fig. 3e, f).

SREBP1c, FXR, and PPARα mRNA expression.
Discussion
The rate of weight gain and the glucose and insulin levels in the maltitol group were significantly lower than in the control group, whereas the total energy intake did not differ between the groups. Most ingested maltitol is fermented by intestinal microbes, 11 and maltitol intake increases bifidobacteria and lactobacilli in healthy human subjects. 12 A recent study revealed that bifidobacteria and lactobacilli can protect against diet-induced obesity. 21,22 Moreover, intestinal bacterial flora converts maltitol into short-chain fatty acids (SCFAs), such as acetic acid, propionic acid, and butyric acid, by fermentation in the intestine. 12 SCFA administration also reportedly prevents diet-induced obesity and improves insulin sensitivity. 23,24 Our results suggest that a change in intestinal flora and production of SCFAs may contribute to a suppressive effect on weight gain, which can be a cause of low blood glucose and insulin levels.
FXR plays an important role in insulin resistance and glucose tolerance. 25 –28 The activation of FXR can decrease the expression of gluconeogenesis enzymes, such as phosphoenolpyruvate carboxykinase and glucose-6-phosphatase, and promote bile acid excretion. 28 Our results suggest that the activation of FXR by maltitol may also contribute to the suppression of blood glucose increases.
Many indigestible components such as dietary fiber and β-glucan reportedly suppress circulating cholesterol levels by absorbing bile acid in the intestine. 29,30 Propionic acid produced by intestinal bacterial flora, in addition to indigestibility, may suppress cholesterol synthesis in the liver 31,32 leading to decreased circulating cholesterol levels. FXR regulates the metabolism of bile acid and cholesterol—FXR knockout mice become hypercholesterolemic. 33 –35 It has also been reported that soluble fiber activated FXR in a high-fat diet model, 36 suggesting that indigestible and soluble fiber, like maltitol, absorb bile acids in the intestine and decrease circulating bile acid levels, leading to the activation of hepatic FXR and increasing circulating bile acid levels.
The hepatic fatty degeneration index, hepatic TG concentration, and the AST plasma levels, an index of hepatocyte damage, were significantly lower in the maltitol group than in the control group, suggesting that maltitol inhibited the advance of hepatic fatty liver degeneration. Through maltitol intake, the expression of SREBP1c was strongly suppressed. Under normal metabolic conditions, SREBP1c activation increases the storage of nutrients. However, under excessive nutrient intake conditions, its elevated expression can lead to obesity-associated disease and lipotoxicity. 13,37 Our results suggest that maltitol suppresses fatty liver degeneration through downregulation of SREBP1c. In addition to decreasing the SREBP1c expression, an FXR agonist has been shown to modulate hepatic monocyte activity and improve NAFLD through an increase in anti-inflammatory macrophages and IL10. 38 PPARα agonist improved hepatic fatty degeneration by enhancing fatty acid β oxidation enzymes such as CPT-1 and acyl Co A oxidase. 39,40 In the present study, the control group showed a significantly higher SREBP1c expression than the normal group, whereas FXR expression did not differ between the control and normal groups. It is possible that the activation of FXR and PPARα may contribute to the suppression of hepatic fatty degeneration independent of SREBP1c or other factors related to the SREBP1c expression. 41 Furthermore, the acetic acid produced as a result of maltitol intake may have also downregulated SREBP1c in the liver. 42 The increase of expression of FXR and PPARα mRNA in the maltitol group may have contributed to the suppression of fatty liver degeneration and is a likely mechanism for the suppression of SREBP1c expression.
HMG-CoAR is suppressed and degraded by low-density lipoprotein, 43 playing a role in cholesterol synthesis and lipid metabolism. 44 An increased HMG-CoAR expression was observed with the amelioration of fatty liver, 45 suggesting that maltitol may promote lipid metabolism in liver. The enhancement of HMG-CoAR expression might be an endogenous compensation due to promoted excretion of bile acid in the liver. Further examination regarding these mechanisms is needed.
Taken together, we suggested the efficacy of maltitol for a mouse NAFLD model as shown in Figure 4.
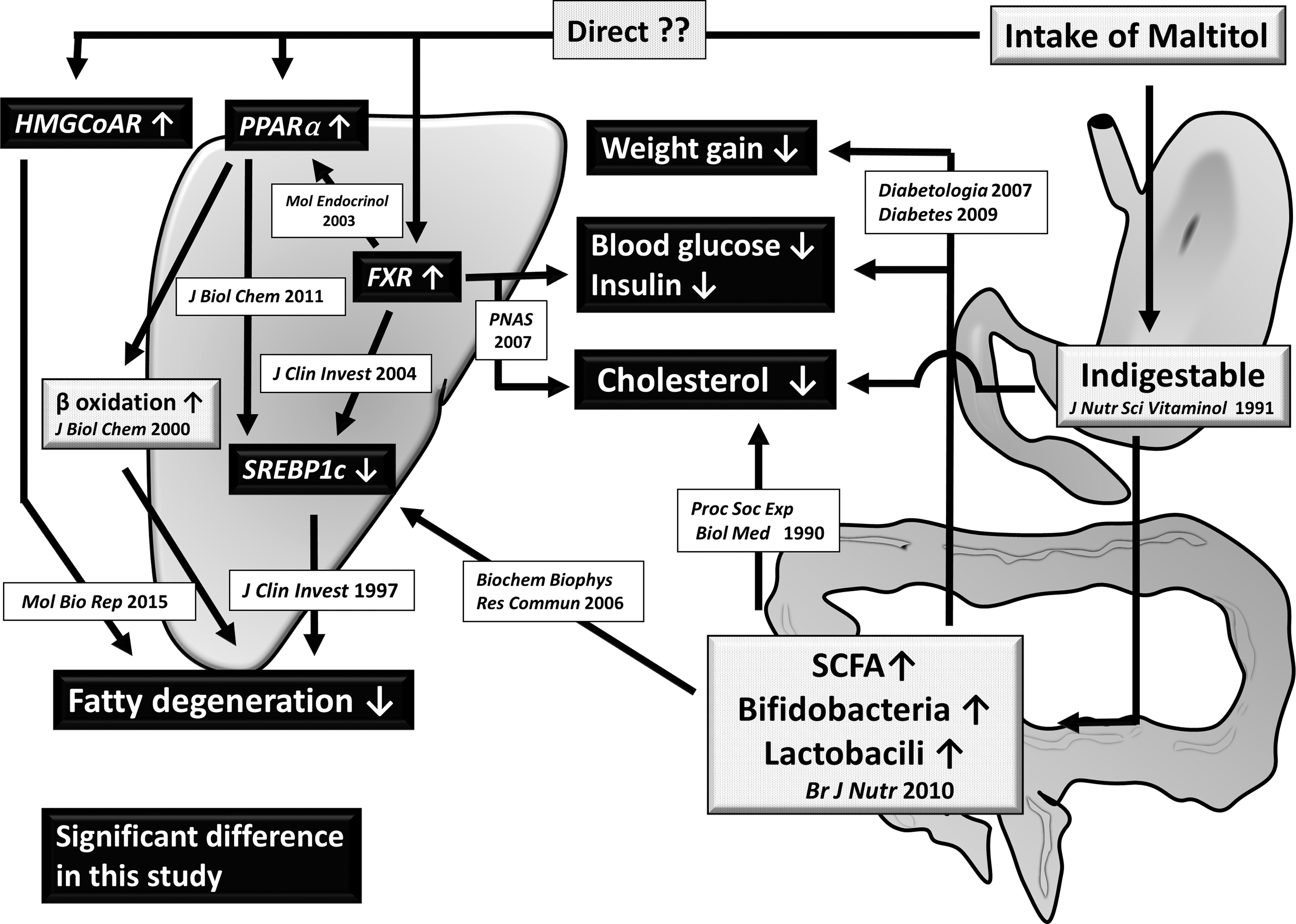
Suggested mechanism for the suppression of hepatic fatty degeneration by maltitol. Gray rectangles indicate previously reported maltitol traits. Black rectangles represent the significant effects of maltitol revealed in the present study. Further study about the enhancement of expression of FXR, PPARα, and HMG-CoAR by maltitol was needed. SCFA, short chain fatty acid.
In conclusion, the present study revealed that supplementation with maltitol prevented obesity, hyperglycemia, hypercholesterolemia, and hepatic fatty degeneration in high-fat diet-fed mice. Thus, maltitol might be useful for treating patients at the initial stages of hepatic fatty degeneration to improve steatohepatitis.
Footnotes
Acknowledgments
We owe our deepest gratitude to Ryosuke Kawaura, Aya Kuretani, Junya Honda and Masaya Kitayama (Ueno Fine Chemicals Industry, Ltd., Osaka, Japan) for their tremendous support.
Author Disclosure Statement
No competing financial interests exist.