
Abstract
Ursolic acid is a lipophilic pentacyclic triterpenoid found in many fruits and herbs and is used in several herbal folk medicines for diabetes. In this study, we evaluated the effects of apple pomace extract (APE; ursolic acid content, 183 mg/g) on skeletal muscle atrophy. To examine APE therapeutic potential in muscle atrophy, we investigated APE effects on the expression of biomarkers associated with muscle atrophy and hypertrophy. We found that APE inhibited atrophy, while inducing hypertrophy in C2C12 myotubes by decreasing the expression of atrophy-related genes and increasing the expression of hypertrophy-associated genes. The in vivo experiments using mice fed a diet with or without APE showed that APE intake increased skeletal muscle mass, as well as grip strength and exercise capacity. In addition, APE significantly improved endurance in the mice, as evidenced by increased exhaustive running time and muscle weight, and reduced the expression of the genes involved in the development of muscle atrophy. APE also decreased the concentration of serum lactate and lactate dehydrogenase, inorganic phosphate, and creatinine, the indicators of accumulated fatigue and exercise-induced stress. These results suggest that APE may be useful as an ergogenic functional food or dietary supplement.
Introduction
S
Muscle atrophy is characterized by a reduction in protein content due to inhibition of protein synthesis and induction of protein degradation, which results in a decrease in muscle fiber size. 5 Maintenance of muscle mass and strength has a significant impact on the quality of life. However, to date, there is no effective therapy specifically focused on the prevention of muscle weakness or fatigue.
Ursolic (3β-3-hydroxyurs-12-en-28-ioc) acid is a lipophilic pentacyclic triterpenoid found in many fruits, such as apples, where it contributes to the composition of the waxy coats and some medicinal plants used for folk herbal medicines to treat diabetes. 6 –9 Recent studies have shown that ursolic acid may exert beneficial effects in various conditions, including Alzheimer's disease, 10 cancer, 11 –13 diabetes, 14,15 and muscle atrophy. 16
Fruit peels have also been shown to contain higher amounts of phenolics than the edible fleshy parts. Thus, apple peel has twice the amount of total phenolics compared to the peeled fruits (3300 mg/100 g dry weight), 17,18 while the lyophilizate of apple pomace contains only about 118 mg/g. 19 The composition of apple peel, in particular its waxy coat, has been extensively studied because of its relevance to the prevention of fruit damage caused by insects and other pathogenic organisms. Ursolic acid is the main component of apple cuticular wax. 20 Although thousands of tons of waste apple pomace are generated each year in the production of apple juice in Korea, the pomace is either discarded or used for animal feed. We hypothesized that ursolic acid could be isolated from apple pomace and used as a valuable food supplement to prevent or treat muscle atrophy.
In this study, we investigated the effect of apple pomace extract (APE) intake on strengthening of skeletal muscles weakened by injury or fatigue during exercise. To discover the underlying molecular mechanisms, we analyzed gene expression in muscle tissue and biochemical parameters in the serum of mice fed an APE-containing diet.
Materials and Methods
Preparation of APE powder
Apple pomace was collected from the region of Gyeongsangbuk-do, Republic of Korea. After washing, dried apple pomace was extracted using 95% ethanol (dried apple pomace:ethanol = 1:10) for 16 h at 25°C; the extract was filtered and concentrated using an evaporator, followed by freeze-drying with maltodextrin as an excipient. Powdered APE was stored at room temperature until use.
Cell culture and treatment
Mouse myoblast cell line C2C12 was purchased from ATCC (Manassas, VA, USA) and maintained in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum and 1% antibiotic–antimycotic (all ingredients from Gibco Life Technologies, Grand Island, NY, USA) at 37°C in an atmosphere of 95% air/5% carbon dioxide (CO2; v/v) in a humidified incubator. When C2C12 cells reached 90% confluence, the medium was replaced with the differentiation medium, that is, DMEM supplemented with 2% horse serum (Gibco) to induce myotube formation. The medium was changed daily until the cells were fully differentiated. C2C12 myotubes were treated with creatine and APE for 18 h 4–5 days after differentiation.
RNA preparation and real-time polymerase chain reaction
Total cellular RNA (or total muscle RNA from 50 mg of mouse hindlimb skeletal muscle) was isolated using the QIAGEN RNA Prep Kit (QIAGEN, Valencia, CA, USA) according to the manufacturer's instructions. Two milligrams of RNA from each sample was reverse transcribed using MuLV reverse transcriptase, 1 mM dNTP, and 0.5 μg/μL oligo(dT)12–18. The resulting cDNA was used as a template for real-time polymerase chain reaction (RT-PCR) to analyze the expressions of AMP-activated protein kinase alpha 1 catalytic subunit (AMPK), sirtuin 1 (SIRT1), proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), insulin-like growth factor 1 (IGF-1), protein kinase B (PKB, also known as Akt), mammalian target of rapamycin (mTOR), ribosomal S6 protein kinase (S6K-alpha-1), E3 ubiquitin-protein ligase TRIM63 (MuRF1), and F-box protein 32 (Atrogin-1) according to the manufacturer's protocol under the following conditions: 95°C for 10 min, followed by 40 cycles at 95°C for 15 sec, and 60°C for 1 min. GAPDH was used as an internal standard. PCR primers were purchased from the Applied Biosystems Corporation (Foster City, CA, USA). The amplified products were analyzed using the QuantStudio 6 RT-PCR program.
Quantitative phosphoprotein assays
AMPK and LKB1 phosphorylation was analyzed in C2C12 cells pretreated with creatine, ursolic acid (both from Sigma-Aldrich, St Louis, MO, USA), and APE for 24 h after differentiation. AMPK phosphorylation was quantified using the PathScan phospho-AMPKα ELISA Kit (Cell Signaling Technology, Beverly, MA, USA) following the manufacturer's instructions; 50 μg of protein from each sample was used in this assay. Phosphorylated LKB1 (Ser428) was quantified using the Phospho-STK11/LKB1 Cell-Based Phosphorylation ELISA Kit (LSBio, Seattle, WA, USA) following the manufacturer's instructions.
Animals and diets
Four-week-old male C57BL/6NHsd mice were purchased from Harlan Laboratories (Houston, TX, USA). Mice were individually housed in separate cages in the animal room maintained at 23 ± 2°C and 55 ± 10% relative humidity with a 12-h light/12-h dark cycle; animals were provided food and water ad libitum. During the 12-week treatment, the exercise control group (n = 10) was fed an AIN-93G diet, and the test groups were fed AIN-93G diets supplemented with APE (n = 10) or creatine (n = 10) as a positive control, respectively. All animal experiments were approved by the Ethics Review Committee of Korea Yakult Company Limited R&D Center, Republic of Korea (KYIACUC-14-01-0002-Y).
Treadmill running test and grip strength measurement
A treadmill running test was performed using a variable speed belt treadmill enclosed in a Plexiglas chamber with a stimulating device consisting of a shock grid attached to the rear of the belt (JD-A-09, Jeong Do B&P, Seoul, Korea). The shock grid was set to deliver 0.2 mA current, which caused an uncomfortable shock but did not physically harm or injure the animals. Mice were acclimatized using a habituation protocol for the running test described previously. 21 An exhaustive running test started at 10 m/min for 10 min with a 5% slope. The speed was then gradually increased from 15 to 20 m/min and maintained constant until exhaustion. When the mice reached exhaustion defined by the inability to run for 10 sec, the electric shock was discontinued and the animals were sacrificed by drawing blood under deep CO2 anesthesia. Forelimb grip strength was determined using a grip strength meter (Bioseb, Vitrolles Cedex, France) equipped with a pull bar. Each mouse was subjected to five consecutive tests to obtain the peak value.
Blood analysis
Blood samples were collected from the heart of sacrificed animals, centrifuged at 1000 g for 15 min at 4°C, and plasma was stored at −80°C until assayed. Plasma lactate and lactate dehydrogenase (LDH), aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), and creatinine were determined using an automated analyzer (Hitachi 7020, Hitachi, Japan), according to the manufacturer's instructions. Plasma inorganic phosphate (iPO) concentration was determined using the QuantiChrom Phosphate Assay Kit (BioAssay Systems, Hayward, CA, USA) according to the manufacturer's instructions.
High-performance liquid chromatography
High-performance liquid chromatography was conducted on an Agilent 1200 Series Rapid Resolution System (Palo Alto, CA, USA), equipped with a quaternary pump with a vacuum degasser, a thermostatted column compartment, an autosampler, and a diode array detector. Separation was performed on a Capcell Pak UG120 C18 column (4.6 ×250 mm; particle size, 5 μm; Shiseido, Tokyo, Japan) end-capped to minimize the effects of unreacted silanol. The isocratic elution system consisted of aqueous 0.1% phosphoric acid in water and acetonitrile (15:85, v/v); the mobile phase was degassed by filtering through a 0.20-μm membrane filter (Millipore; Billerica, MA, USA) and applied at the flow rate of 0.8 mL/min and operating temperature of 35°C. Elution was monitored by UV absorbance at 210 nm.
Statistical analysis
The data are expressed as the mean ± SD of at least three independent experiments performed in triplicate. Significant differences between the groups were determined using the unpaired Student's t-test and considered statistically significant at the P value < .05.
Results
APE effect on gene expression and LKB1/AMPK phosphorylation in C2C12 myotubes
To investigate the effect of APE on mitochondrial biogenesis in the skeletal muscle, we examined AMPK activation by assessing its mRNA level in C2C12 cells. As shown in Figure 1a, the APE-treated C2C12 myotubes exhibited higher expression levels of AMPK, SIRT1, and PGC-1α in APE dose-dependent manners, compared to the untreated control cells. A previous study has shown that skeletal muscle hypertrophy can be mediated by the induction of IGF-1, a protein growth factor known to promote the accumulation of skeletal muscle mass. 22 IGF-1 acts in part by stimulating the phosphatidylinositol-3 kinase (PI3K)/Akt pathway, resulting in the downstream activation of transcription factors, which induce biosynthesis of the target proteins. 2,23 In addition to stimulating protein expression, IGF-1 inhibits the activation of signaling pathways related to skeletal muscle atrophy. A distinct set of genes, including Atrogin-1, 24 is downregulated by IGF-1-induced hypertrophy, whereas MuRF1 is significantly upregulated in atrophy. 25 Therefore, we examined APE effects on the levels of the IGF-1, Akt, mTOR, and S6K1 genes and found that APE significantly upregulated their mRNA expressions in the skeletal muscle (Fig. 1b). We also tested whether APE increased muscle mass by inhibiting atrophy-associated gene expression in the skeletal muscle. Indeed, APE reduced mRNA levels of Atrogin-1 and MuRF1, suggesting that APE may have a role in the reduction of muscle atrophy (Fig. 1c). To examine a potential role of AMPK activation in APE effects, we treated myotubes with creatine, ursolic acid, and APE. As shown in Figure 1d, the treatment with 10 μg/mL ursolic acid and 50 μg/mL APE increased the phosphorylation of AMPK and LKB1. Given that APE contains ∼183 mg/g of ursolic acid, 50 μg/mL APE should have an estimated amount of 10 μg/mL ursolic acid, which may account for its stimulation of AMPK and LKB1 phosphorylation in C2C12 myotubes.
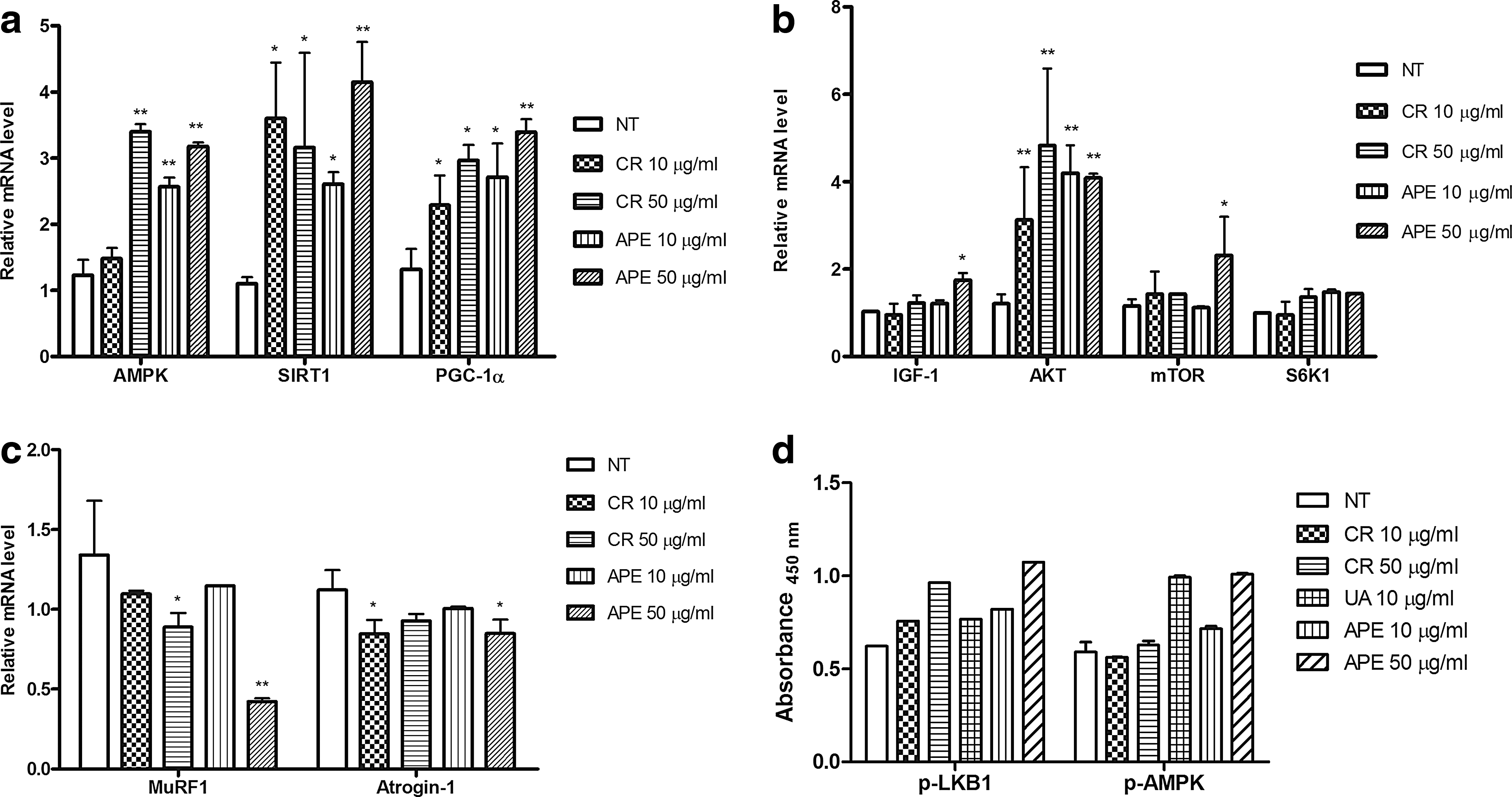
APE induced the expression of AMPK and IGF-1/Akt pathway-related genes in skeletal myotubes in vitro. APE upregulated mRNA levels of AMPK associated with mitochondrial activity
APE increases endurance and reduces fatigue of mice during treadmill exercise
Since APE increased the expression of genes related to mitochondrial biogenesis and muscle hypertrophy and inhibited that of genes associated with muscle atrophy, we examined whether APE could improve mouse exercise capacity. There were no significant differences in body weight and food intake between the control group and creatine- or APE-fed animals during the 12-week feeding period (Fig. 2a). At the end of the treatment, we performed the treadmill running test, after which mice were sacrificed, and blood samples were collected and analyzed. The treadmill endurance test showed that running distance and time-to-exhaustion in the APE-treated groups were increased compared to the creatine-treated and control groups (Fig. 2b, 2c). Serum parameters related to fatigue were significantly higher in the exercised control group compared to the normal (nonexercised) control group (P < .01). Blood analysis indicated that after the treadmill running test, serum levels of lactate, LDH, iPO, AST, ALT, ALP, and creatinine in the APE-treated groups were significantly lower compared to the exercised control group (Table 1). These results suggest that APE could effectively lower serum levels of fatigue indicators in mice.

Effects of APE-supplemented diet on total exercise time and running distance. Mice were provided ad libitum access to control diet supplemented or not with creatine or APE for 12 weeks and their body weight (g) was measured every week. Weekly weight gain rate (%) was calculated relative to the weight at the start of the experiment
The data are presented as the mean ± SD.
P < .01 vs. normal (nonexercise group); a P < .05, b P < .01 vs. control.
ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; iPO, plasma inorganic phosphate; LDH, lactate dehydrogenase.
APE increases skeletal muscle strength and induces muscle hypertrophy
To further investigate the effect of APE intake on exercise capacity in the mouse model, we measured grip strength and analyzed the weight of hindlimb skeletal muscle in mice fed an APE-containing diet.
As shown in Figure 3a, grip strength in the control, creatine-75, APE-75, APE-150, and APE-300 groups was 3.43 ± 0.52, 4.09 ± 0.34, 4.74 ± 0.29, 4.89 ± 0.67, and 4.80 ±0.51 g, respectively, demonstrating significantly increased values for the APE- and creatine-fed animals compared to the control group. Importantly, APE increased mouse grip strength in a dose-dependent manner (P < .01). We also measured hindlimb skeletal muscle weight for each mouse and found that APE significantly increased muscle weight in a dose-dependent manner (Fig. 3b, P < .01). To investigate the molecular mechanisms underlying APE effects in mice, the expression levels of MuRF1, Atrogin-1, and IGF-1 were determined by RT-PCR (Fig. 4). MuRF1 and Atrogin-1 expressions were significantly lower in the muscle tissue of the APE-treated animals compared to the control mice (P < .01; Fig. 4). However, IGF-1 mRNA expression in skeletal muscles was slightly higher in the APE-fed mice compared to control, although the difference was not statistically significant. Overall, these data suggest that APE intake can improve grip strength through induction of muscle hypertrophy.
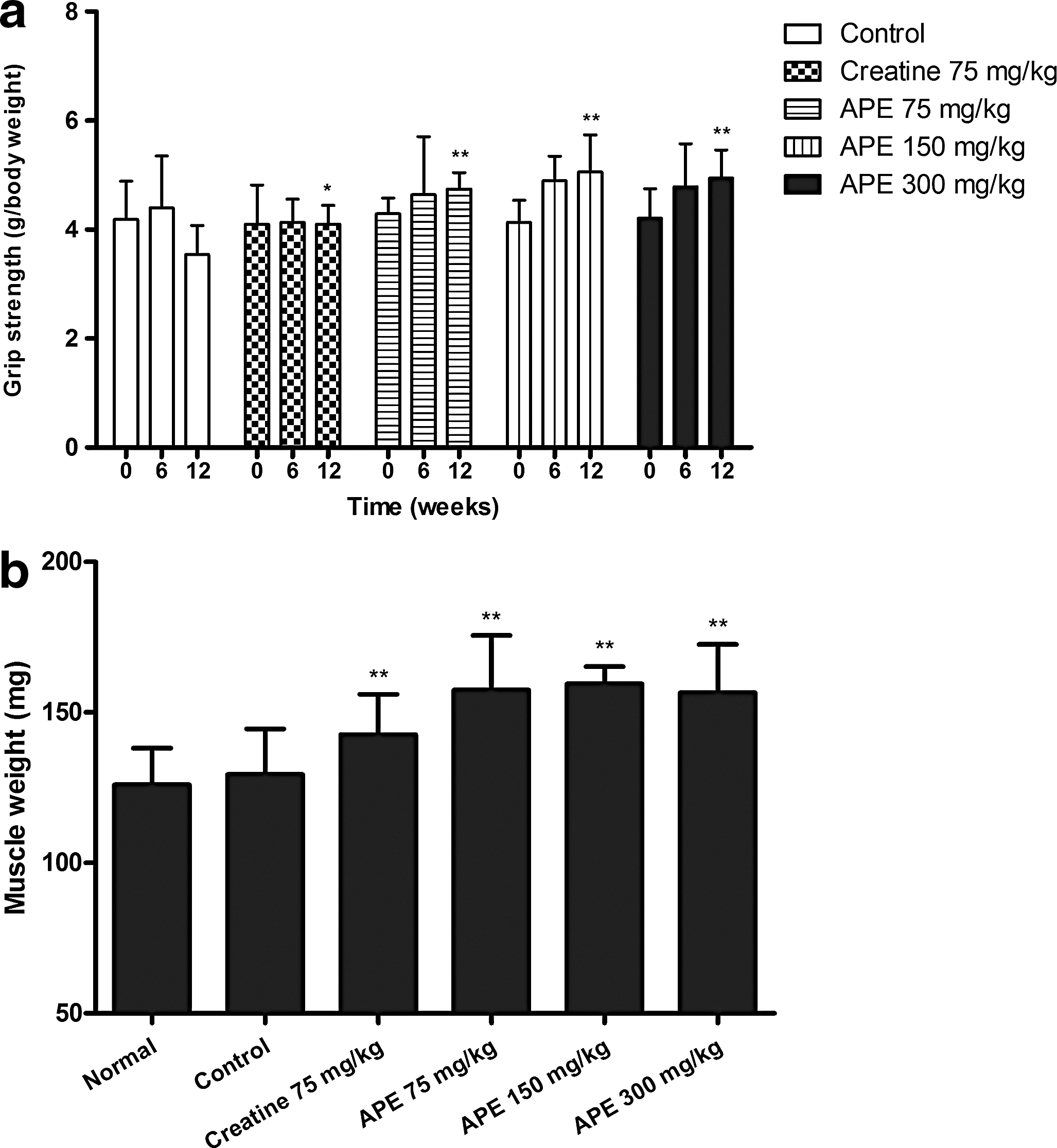
Effects of APE-supplemented diet on grip strength and muscle weight. Mice were provided ad libitum access to control diet supplemented or not with creatine or APE for 12 weeks before grip strength was measured and tissues were harvested. Forelimb grip strength was determined using a grip strength meter equipped with a triangular pull bar. Each mouse was subjected to five consecutive tests to obtain the peak value

APE-supplemented diet downregulated the expression of MuRF1 and Atrogin-1 related to skeletal muscle atrophy and upregulated IGF-1 related to muscle growth. Mice were provided ad libitum access to control diet supplemented or not with creatine or APE for 12 weeks; then, skeletal muscles were harvested and analyzed for the expression of indicated genes by RT-PCR. The data are shown as the mean ± SD of three independent experiments; asterisks indicate significant differences: *P < .05 and **P < .01 vs. the control group. RT-PCR, real-time polymerase chain reaction.
Discussion
Skeletal muscle atrophy is a physiological outcome of disuse, aging, and disease and is characterized by a decrease in protein content and fiber size because of reduced protein synthesis and increased degradation. 5 Recent studies have revealed that ursolic acid induces insulin/IGF-1 signaling in the skeletal muscle, leading to Akt activation, muscle hypertrophy, and reduced adiposity and blood glucose normalization. 16 Another study has found that ursolic acid increases the Akt activity in the muscle of high-fat fed mice. Ursolic acid-mediated increase of Akt activity in the skeletal muscle has been associated with muscle hypertrophy, increased energy expenditure, and reductions in total body weight, white fat, glucose intolerance, and hepatic steatosis. 26
In addition, it has recently been reported that AMPK activation can improve mice endurance in the absence of exercise training. 27 Under the endurance exercise condition, the skeletal muscle suffers a number of changes, such as decrease in glucose consumption, transition from glucose to fatty acids as a main energy source, increase in mitochondrial biogenesis, and fiber-type switching. 28 AMPK activation has been shown to act as a switch to ATP-producing pathways to enhance endurance, whereas PGC-1α functions as the master regulator in mitochondrial biogenesis and skeletal muscle adaptation. 29 –31
In this study, we aimed to determine the effect of ursolic acid-containing APE on muscle strength and physical activity and on the expression of biomarkers associated with muscle weakness and fatigue. We found that APE induced the expression of genes in the AMPK and IGF-1/Akt/mTOR pathways related to the mitochondrial activity in mice, as well as the levels of hypertrophy-associated genes in cultured myotubes, at the same time reducing mRNA of atrophy-associated genes. We also found that treatment with 10 μg/mL ursolic acid and 50 μg/mL APE increased the phosphorylation of AMPK and LKB1. Such expression pattern is expected to result in the increase of skeletal muscle weight and improved physical capacity in exercise. Indeed, in the treadmill exercise experiment, the APE-fed mice demonstrated increased running time and decreased fatigue biomarkers in serum.
The levels of serum lactate, LDH, creatinine, and iPO are known to serve as indicators of accumulated fatigue and exercise-induced stress. 32,33 Blood lactate is the product of carbohydrate conversion under anaerobic conditions, that is, glycolysis, which is the main energy source for short-term intensive exercise. Since accumulation of blood lactate causes fatigue during physical exercise, rapid lactate removal is beneficial for relieving fatigue. 34
After the treadmill running test, the level of serum lactate was significantly lower in the APE-fed mice compared to the control group (Table 1). Increased serum LDH indicates tissue damage in the liver and muscle, suggesting that high cortisol levels might induce muscle atrophy. APE intake effectively inhibited LDH increase in mice (Table 1), further supporting the antifatigue effect of APE. Serum levels of AST, ALT, and ALP are important for the assessment of liver damage. 35 Our results indicate that APE intake could decrease serum AST, ALT, and ALP, suggesting that an APE-containing diet can prevent fatigue by improving liver functions.
We also analyzed the functional effects of APE administration (75, 150, and 300 mg/kg) in mice by assessing grip strength, muscle weight, and expression of the genes associated with atrophy of skeletal muscles. Our results showed that grip strength and muscle weight were significantly enhanced in the APE-fed groups, whereas the genes related to muscle atrophy were inhibited by the administration of 150 and 300 mg/kg APE (Fig. 4). These results indicate that APE is likely to exert positive effects on muscle strength and weight through inhibition of atrophy-associated gene expression in the skeletal muscle.
In conclusion, our data suggest that APE may exert beneficial effects on exercise endurance by downregulating factors related to fatigue and enhancing muscle strength and weight through inhibition of atrophy-associated genes in the skeletal muscle. Such effects may, in part, be related to amelioration of skeletal muscle injury. Further studies are required to elucidate the mechanisms underlying APE antifatigue effects. In addition, clinical trials with human participants may be considered.
Footnotes
Acknowledgment
This research was supported by the High Value-added Food Technology Development Program, Ministry of Agriculture, Food and Rural Affairs (113033-02-2-SB010).
Author Disclosure Statement
No competing financial interests exist.