
Abstract
Accumulating evidence indicates that overconsumption of ethanol contributes in many ways to the pathogenesis of hepatic injury. Although studies indicate that taurine decreases lipogenesis, oxidative stress, and inflammatory cytokines, the protective effect of taurine against alcohol-induced liver injury is still unclear. To clarify the precise signaling involved in the beneficial effect of taurine on alcohol-induced liver injury, rats were randomly divided into four treatment groups: (1) control (Ctl), (2) alcohol (Alc), (3) Alc+taurine (Tau), and (4) Alc+silymarin (Sil). The Tau and Sil groups had lower lymphocyte infiltration and significantly lower TLR-4/MyD88 and IκB/NFκB compared to the Alc group. The inducible nitric oxide synthase (iNOS), C-reactive protein (CRP), tumor necrosis factors (TNF)-α, interleukin (IL)-6, and IL-1β were also significantly lower in the Tau and Sil groups than in the Alc group. The experimental results indicated that hepatoprotection against alcohol-induced inflammation may be mediated by decreased TLR-4/MyD88 signaling.
Introduction
E
Indeed, both animal and clinical studies indicate that chronic hepatitis induced by alcohol should be classified as an alcoholic liver disease with an immunopathological etiology and should be considered a step in the development of liver cirrhosis, fibrosis, and liver cancer. 5 –7
Antioxidants, that is, free radical scavenging compounds, reportedly ameliorate oxidative stress-related hepatic damage. 8 One well-known antioxidant is silymarin, which is the active ingredient extracted from Silybum marianum (milk thistle). Silymarin has chemoprotective effects against liver injuries such as hepatoxicity induced by carbon tetrachloride (CCl4), liver fibrosis, and ethanol-induced hepatic inflammation. 9 –11 Recent clinical trials also indicate that long-term administration of silymarin significantly increases the survival time of patients with alcohol-induced liver cirrhosis. 12 In fact, silymarin is apparently the complementary alternative medicine most widely used for treating alcohol-induced liver disease in the United States. 13 Accordingly, silymarin is often used as a positive control in hepatoprotective studies.
Taurine (2-amino ethanesulfonic acid) is a major free intracellular amino acid in many mammalian tissues, for example, the brain, retina, myocardium, skeletal muscle, liver, platelets, and leukocytes. 14 Taurine is not considered an essential amino acid since it does not participate in peptide bond formation. 15 Although taurine can be synthesized from methionine and cysteine, the biosynthesis rate is slow in mammals. 16 Therefore, taurine is mainly obtained by consuming eggs, meat, and seafood. 15,17 Recent studies indicate that taurine is an antioxidant with crucial roles in detoxification, membrane stabilization, osmoregulation, neuromodulation, and brain and retinal development. 18
For example, in a rat model of chronic alcohol consumption, rats given taurine supplements at a dosage of 1 g Tau/kg body weight showed that alcohol metabolism was accelerated by upregulation of alcohol dehydrogenase, catalase, and aldehyde dehydrogenase activities, whereas microvesicular steatosis and necrotic cells in the liver were reduced. 19 Our recent study indicated that taurine downregulates lipogenesis, oxidative stress, and inflammatory cytokines in a rat model of chronic alcohol consumption. 20 However, the precise signaling involved in the beneficial effect of taurine on alcohol-induced liver injury is still obscure. Therefore, this study investigated the precise signaling conferred by taurine in alcohol-induced liver injury.
Materials and Methods
Animals
This study was approved by the Institutional Animal Care and Use Committee at Chung Shan Medical University (IACUC Approval No. 1315). Animal welfare and experimental procedures were performed according to the NIH Guide for the Care and Use of Laboratory Animals. The experimental animals were purchased and grouped as described elsewhere. 21 Thirty-two male Wistar rats (body weight: ∼190 g) were purchased from BioLASCO Taiwan Co., Ltd. (Taipei, Taiwan). Rats were housed individually in cages in an animal room at 22°C±2°C with a 12-h light–12-h dark cycle.
Diet
The rats were given chow containing 48.7% (w/w) carbohydrate, 23.9% (w/w) protein, 5.0% (w/w) fat, 5.1% (w/w) fiber, and 7.0% ash (Laboratory Rodent Diet 5001; PMI Nutrition International/Purina Mills LLC., Richmond, IN, USA) with water for 1 week to allow acclimation. The rats were then fed chow and water ad libitum. Chronic alcoholic hepatitis in rats was induced as described elsewhere. 20 Since our previous study indicated that a dosage of 1 g taurine/kg BW and 0.25 g silymarin/kg BW accelerated alcohol clearance in a rat model of chronic alcohol consumption, 20 the rats in the current study were randomly divided into four treatment groups with eight rats each: (1) control group (Ctl): 3 mL of phosphate-buffered saline (PBS) per day (p.o.); (2) alcohol group (Alc): 3 g alcohol/kg BW in 3 mL of distilled water per day (p.o.); (3) 1× taurine group (Tau): 3 g alcohol/kg BW+1 g Tau/kg BW in 3 mL of sterile distilled water per day (p.o.); and (4) 1X silymarin group (Sil): 3 g alcohol/kg BW+0.25 g silymarin/kg BW/day in 3 mL of sterile distilled water per day (p.o.). The experimental period lasted for 6 weeks and then rats were sacrificed by CO2 asphyxiation. The liver tissues were collected and stored at −80°C until use.
Hematoxylin–eosin staining
The liver samples were excised and soaked in formalin and covered with wax. 22 Slides were prepared by deparaffinization and dehydration. They were passed through a series of graded alcohols (100%, 95%, and 75%), 15 min each. The slides were then dyed with hematoxylin. After gently rinsing with water, each slide was soaked with 85% alcohol, 100% alcohol I and II for 15 min each, and finally they were soaked with Xylene I and Xylene II. Photomicrographs were obtained using Zeiss Axiophot microscopes (Carl Zeiss, Inc., Thornwood, NY, USA).
Preparation of liver homogenates
Livers were homogenized with PBS (pH 7.0, containing 0.25 M sucrose) at a 1:9 ratio (liver:PBS, weight:volume) on ice, and the filtrate was collected for further analyses. The protein level in the filtrate was measured according to the procedures specified in the Bio-Rad Protein Assay Kit (Cat No. 500-0006; Bio-Rad Laboratories, Inc., Hercules, CA, USA) with bovine serine albumin as a standard.
Immunoblotting
Protein samples were separated in 10% or 12.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electrophoretically transferred to the nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ, USA) described elsewhere. 21,23 After blocking with 5% nonfat dry milk in PBS, antibodies against tumor necrosis factors (TNF)-α, interleukin (IL)-6, IL-1β, inducible nitric oxide synthase (iNOS), C-reactive protein (CRP), toll-like receptor (TLR)-4, myeloid differentiation factor (MyD)88, extracellular signal-regulated kinases (ERK), phosphorylated-ERK (p-ERK), P38, p-P38, IκB, p-IκB, p-nuclear factor (NF)-κB p65 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and β-actin (Upstates, Charlottesville, VA, USA) were diluted in PBS with 2.5% bovine serum albumin and incubated for 1.5 h with gentle agitation at room temperature. The membranes were washed twice with PBS-Tween for 1 h and a secondary antibody conjugated with horseradish peroxidase (HRP) (Santa Cruz Biotechnology) was added. Pierce's Supersignal West Dura HRP Detection Kit (Pierce Biotechnology, Inc., Rockford, IL, USA) was used to detect antigen–antibody complexes. Quantification of results was performed by densitometry (Appraise; Beckman-Coulter, Brea, CA, USA).
Statistical analyses
All statistical analyses were performed using SPSS 10.0 software (SPSS, Inc., Chicago, IL, USA). Three independent experiments were repeated. Statistical analyses were performed using ANOVA plus posterior multiple comparison test to test the difference. P<.05 was considered statistically significant.
Results
Taurine decreases hepatic lymphocyte infiltration and the expression of hepatic iNOS and CRP proteins in a rat model of chronic alcohol consumption
To clarify the anti-inflammatory effects of taurine in rats with chronic alcohol consumption, this study measured hepatic lymphocyte infiltration and expressions of iNOS and CRP protein in the livers of rats given PBS, alcohol, taurine, and silymarin. Notably, lymphocyte infiltration of the liver was markedly higher in the Alc group compared with the Ctl group (Fig. 1A). In contrast, lymphocyte infiltration in the liver was lower in the Tau and Sil groups than in the Alc group. The iNOS and CRP proteins and hepatic inflammation-associated proteins were significantly higher in the Alc group than in the Ctl group (Fig. 1B). In contrast, the iNOS and CRP protein levels in the liver were significantly lower in the Tau and Sil groups than in the Alc group (Fig. 1B, C).
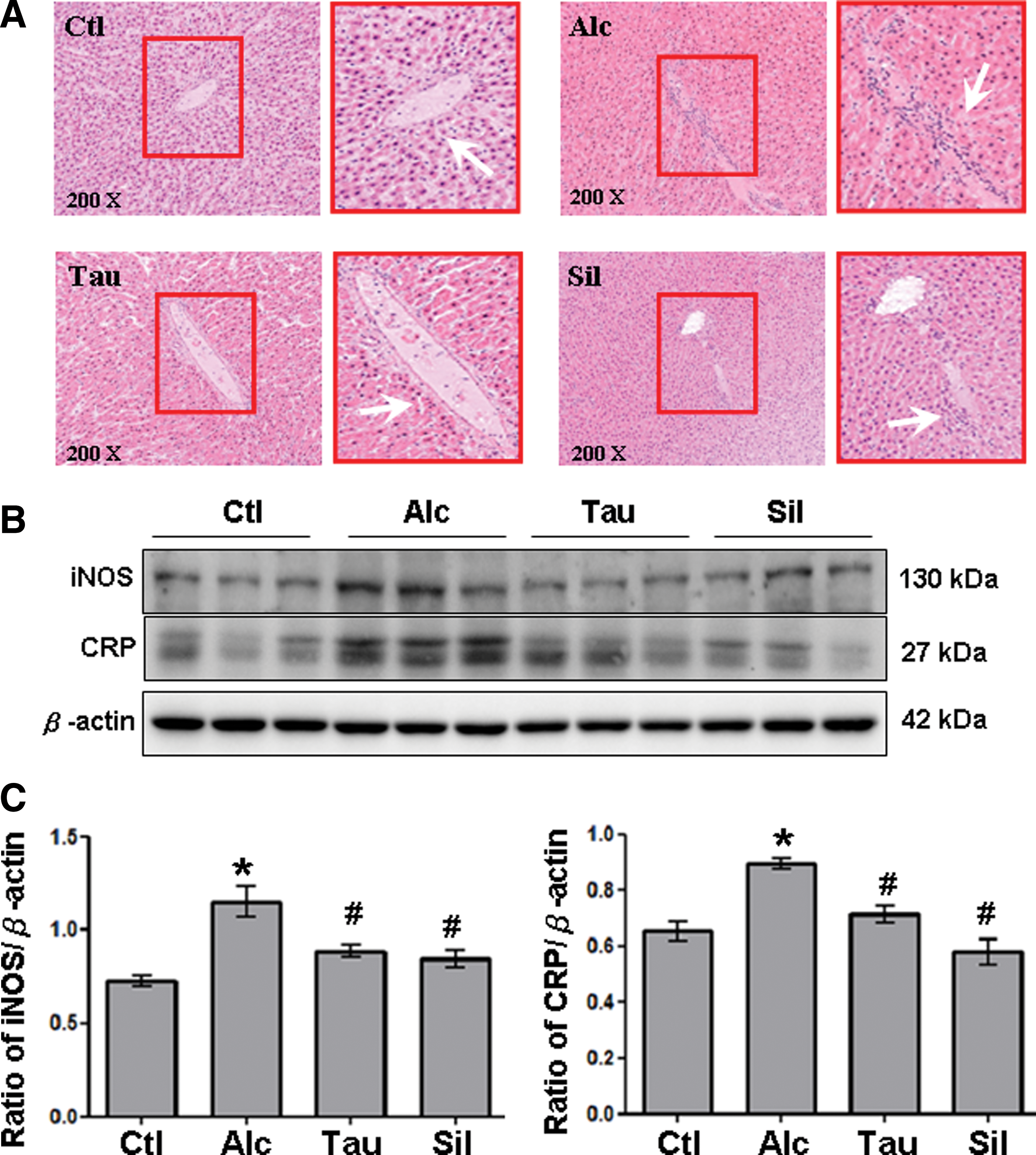
Detection of liver damage indices. Lymphocyte infiltration and expression of inducible nitric oxide synthase (iNOS) and C-reactive protein (CRP).
Taurine decreases the expressions of TLR-4/MyD88 and IκB/NFκB p65 signaling in rats with chronic alcohol consumption
The signaling involved in the protective effect of taurine on alcohol-induced hepatic injury was further investigated by studying the signaling molecules, TLR-4 and IκB/NF-κB p65. Expressions of TLR-4 and its consequent molecule MyD88 in the liver were significantly higher in the Alc group than in the Ctl group (Fig. 2). In contrast, expressions of both TLR-4 and MyD88 in the liver were significantly lower in the Tau and Sil groups than in the Alc group (Fig. 2).

Expression of toll-like receptor (TLR)-4 and myeloid differentiation factor (MyD)88. Liver lysates obtained from different groups of rats (Ctl, Alc, Tau, and Sil) were probed with antibodies against
Experiments were also performed to examine phosphorylated-ERK1/2 (p-ERK1/2) and phosphorylated-P38 (p-P38), the downstream molecules of TLR-4/MyD88. Expressions of p-ERK1/2 and p-P38 in the liver were significantly higher in the Alc group than in the Ctl group (Figs. 3A, B). In contrast, expressions of p-ERK1/2 and p-P38 in the liver were significantly lower in the Tau and Sil groups than in the Alc group (Fig. 3A, B). Moreover, p-IκB and p-NF-κB p65, another downstream signaling of TLR-4/MyD88, were examined. Notably, expressions of p-IκB and p-NFκB p65 in the liver were significantly higher in the Alc group than in the Ctl group (Fig. 4A, B). In contrast, significant decreases in both p- IκB and p-NFκB p65 were observed in the livers of rats from Tau and Sil groups compared to those from the Alc group (Fig. 4, B).

Expression of p-ERK1/2 and p-P38. Liver lysates obtained from different groups of rats (Ctl, Alc, Tau, and Sil) were probed with antibodies against
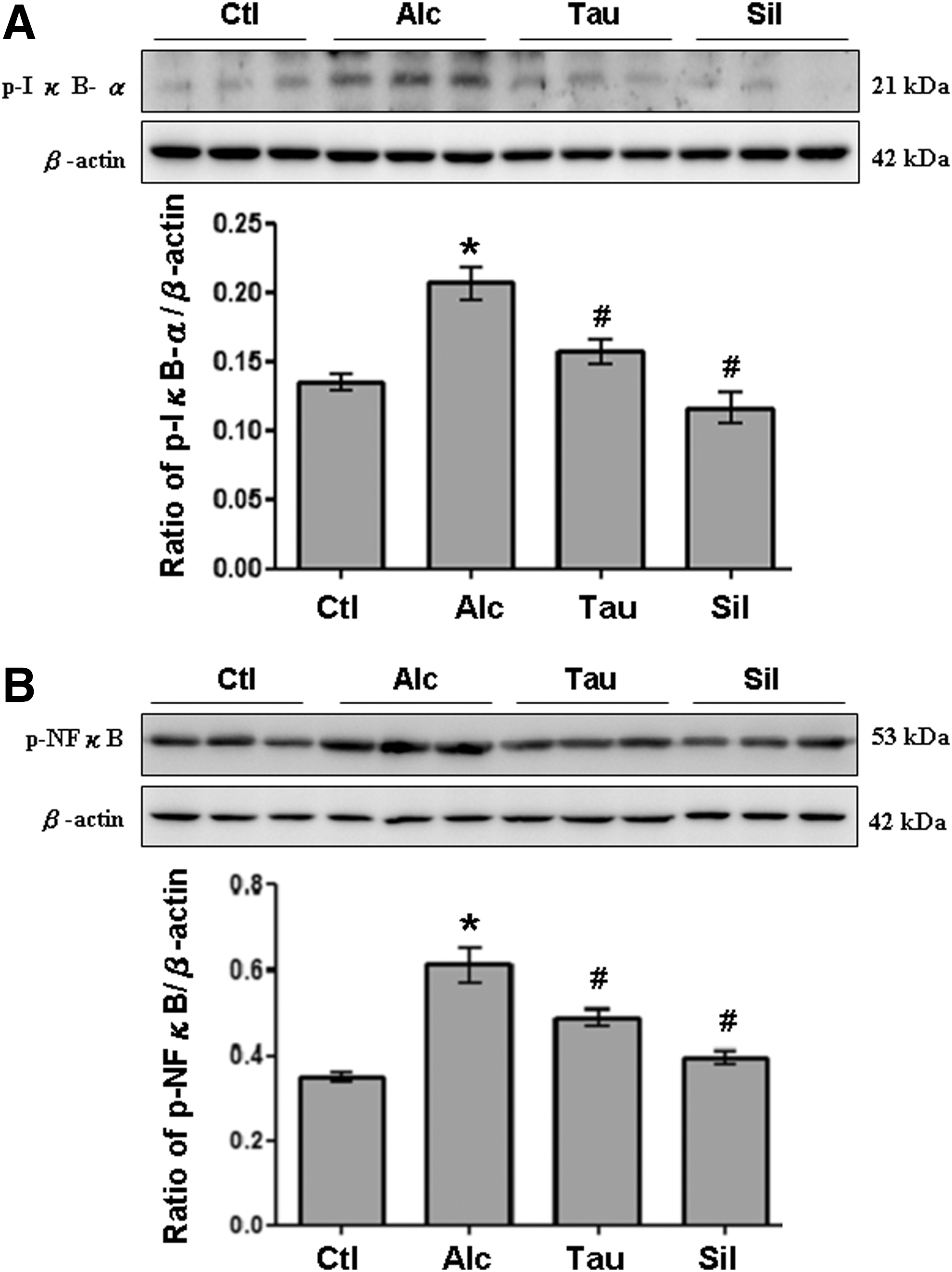
Expression of p-IκB and p-NF-κB. Liver lysates obtained from different groups of rats (Ctl, Alc, Tau, and Sil) were probed with antibodies against
Taurine decreases the expressions of serum and hepatic IL-1β, IL-6, and TNF-α in rats with chronic alcohol consumption
The consequent inflammatory cytokines, TLR-4/MyD88, p-IκB, and p-NF-κB p65 signaling, and IL-1β, IL-6, and TNF-α, were studied to elucidate anti-inflammatory effects of taurine on alcohol-induced liver inflammation. The IL-1β, IL-6, and TNF-α in serum were significantly higher in the Alc group than in the Ctl group (Fig. 5A). Conversely, IL-1β, IL-6, and TNF-α in the serum were significantly lower in the Tau and Sil groups than in the Alc group (Fig. 5). Accordingly, similar findings were observed in rat livers. The IL-1β, IL-6, and TNF-α in the liver were significantly higher in the Alc group than in the Ctl group (Fig. 6A). In contrast, IL-1β, IL-6, and TNF-α in the liver were significantly lower in Tau and Sil groups than in the Alc group (Fig. 6A). Figures 6B–D show the quantification results for IL-1β, IL-6, and TNF-α, respectively.

Expression of serum interleukin (IL)-1β, IL-6, and tumor necrosis factors (TNF)-α. Serum samples obtained from different groups of rats (Ctl, Alc, Tau, and Sil) were probed with antibodies against
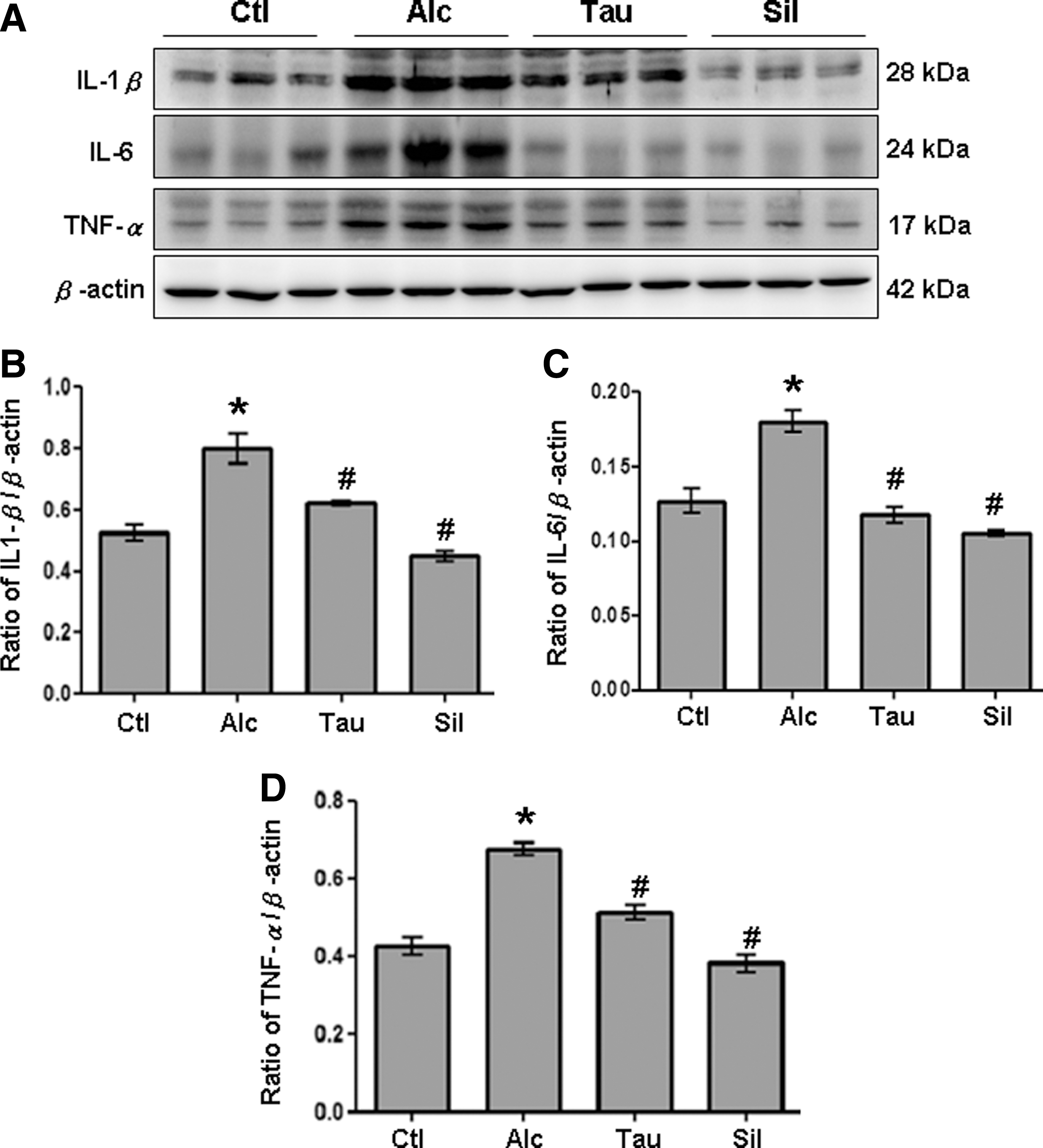
Expression of hepatic IL-1β, IL-6, and TNF-α. Liver lysates obtained from different groups of rats (Ctl, Alc, Tau, and Sil) were probed with antibodies against
Discussion
Studies indicate that natural products are the best way to treat some physiological disorders. Taurine is known to be a vital amino acid with multiple functions, including regulation of the immune responses and protection of biological systems against various injuries. 22,24 –26 Taurine is also known to have potent antioxidant properties and to regulate proinflammatory cytokines. 27 –29 Although taurine has proven effective for treating alcohol-induced hepatic injury, 30 the precise signaling involved in its protective activity is still unclear. This study further showed that by inhibiting TLR-4/MyD88 and IκB/NFκB p65 signaling, taurine suppresses inflammation-associated proteins and cytokines, which alleviates chronic alcohol-induced liver inflammation.
TLR-4 protein encoded by the TLR-4 gene is a member of the TLR family, which plays important roles in pathogen recognition and activation of innate immunity. 31 Activation of innate immunity by TLR-4 signaling is known to be a key process in chronic liver disorders. 32 TLR-4 transduces signals through MyD88, a common signal adaptor molecule. This cascade leads to the activation of NF-κB and results in the production of various proinflammatory cytokines, such as IL-1β, TNF-α, and IL-6. 33 Subsequently, this process induces the expressions of inflammatory markers such as CRP and iNOS. 34,35 CRP, produced by hepatocytes, is an annular pentameric protein, the level of which rises in response to inflammation. 36,37 In addition, iNOS, an isoform of NOS enzyme family, could produce large amounts of NO as an immune defense mechanism. 35 Both CRP and iNOS are known to be induced by various proinflammatory cytokines, including IL-1β, IL-6, and TNF-α, and elevated levels of CRP and iNOS are recognized as markers in alcohol-induced liver injury. 34,35 Consistently, similar results were observed in rats with chronic alcohol consumption in this study. In contrast, taurine was demonstrated to reduce the expression of hepatic inflammatory molecules such as iNOS, CRP, TNF-α, IL-6, and IL-1β and the activation of TLR-4/MyD88 and NF-κB signaling in rats with chronic alcohol consumption. Accordingly, these findings suggest a protective role of taurine on alcohol-induced liver injury by decreasing the hepatic levels of CRP and iNOS through TLR-4/MyD88 and NF-κB signaling along with the decreases in proinflammatory cytokines, including TNF-α, IL-6, and IL-1β. Although these findings provide a possible mechanism for taurine in alleviating alcohol-induced liver inflammation, further studies are still needed to clarify whether taurine alleviates the alcohol-induced liver inflammation by directly acting on these inflammatory molecules.
Overconsumption of alcohol causes various morphological and functional changes in the liver that contribute to various hepatic pathological processes such as steatosis, inflammation, fibrosis, cirrhosis, and carcinoma. 38,39 In terms of morphology, hepatic cirrhosis or even death can result from macrovesicular steatosis, Mallory–Denk bodies (MDB), megamitochondria, occlusive lesions of terminal hepatic venules, and pericellular fibrosis. 40 Studies show that MDBs occur in the cytoplasm of hepatocytes in 65% and 51% of patients with alcoholic hepatitis and alcoholic cirrhosis, respectively. 41 Formation of MDBs is known to increase IL-1β and its subsequent cytokines, for example, TNF-α. Liver biopsies from alcoholic hepatitis patients often reveal IL-1β-forming macrophages. The IL-1β activates expressions of interferon (IFN)-γ and TNF-α receptors. The IFN-γ and TNF-α induce two pathways of MDB formation: one pathway is epigenetic changes resulting in the immunoproteasome-26S proteasome switch; another pathway is the TLR-4- NF-κB-ERK-P38 phosphorylation. 42 By activating the TLR-4 signaling pathway, both factors contribute to inflammation and subsequent injuries. 43 Moreover, NF-κB activation is known to be an essential factor in MDB formation. Experiments in mice have shown that MDBs can be induced in the liver by consumption of diethyl 1,4-dihydro-2,4,6-trimethyl-3,5-pyridinedicarboxylate (DDC). Primary cultures of DDC-treated hepatocytes upregulate IKK, ERK, JNK, and P38 after 3 h of culturing. 42 NF-κB is upregulated by the sixth day of in vitro culture when MDBs form spontaneously and by the 10th week of in vivo DDC feeding. Formation of MDB can also be prevented by specific inhibitors of NF-κB, JNK, or P38. 42,44 Indeed, these findings might indicate a positive correlation between formation of MDBs and activation of TLR-4/MyD88 and NF-κB/ERK/P38 signaling. Although our experimental findings revealed inhibitory effects of taurine on alcohol-induced liver injury by suppressing TLR-4/MyD88 and NF-κB/ERK/P38 signaling, the precise roles of taurine on MDB formation are still unclear and need further investigations.
Although both taurine and silymarin have beneficial effects on alcohol-induced liver injury, a very recent study reported a possible side effect of silymarin, that is, in patients with both type-2 diabetes and liver injury, silymarin induces insulin resistance. 45 This side effect is attributable to the ability of silymarin to disrupt insulin signaling by increasing phosphatase and tensin homolog expression. This study is the first to demonstrate that, when integrated with the protective mechanism of silymarin, taurine alleviates alcohol-induced liver injury by reducing IκB/NF-κB and TLR-4/MyD88 signaling. These experimental results may help researchers develop an alternative remedy for alcohol-induced liver injury and suggest that taurine is a potential ingredient for hepatoprotective health foods.
Footnotes
Acknowledgments
This work was funded by the Chung Shan Medical University and Changhua Christian Hospital cooperative project CSMU-CCH-102-02. Consumptive material for animal experiments was partially supported by the National Science Council grant NSC99-2320-B-040-007-MY3. T.C.H., C.C.C., Y.C.C., and M.L.C. performed the animal study, zymography, hematoxylin and eosin staining, and Western blot. C.J.L., T.C.H., and B.S.T. conceived this study, drafted the manuscript, and performed the statistical analyses. All authors read and approved the final manuscript. Ted Knoy is appreciated for his editorial assistance. The upright optical microscope, chemiluminescence imaging analyzer, and gel digital imaging analyzer from the Instrument Center of Chung Shan Medical University were utilized in this study, which is partly supported by the National Science Council, Ministry of Education, and Chung Shan Medical University.
Author Disclosure Statement
No competing financial interests exist.