
Abstract
Nonalcoholic fatty liver disease, a major cause of abnormal liver function, is often associated with obesity. Arginine (ARG) plays a role in modulating body weight/fat, but limited data exist as to the role of ARG in soy protein's ability to protect from liver steatosis. We investigated the role of native ARG in the soy protein isolate (SPI) in reducing liver steatosis in male obese Zucker rats. Rats (N=48; 6 weeks old) were randomly assigned to one of three diets for 8 or 16 weeks: the casein (CAS) diet as control (0.6% ARG), CAS diet supplemented to contain 1.3% ARG, or an SPI diet containing isoflavones (1.3% ARG). SPI and ARG rats gained significantly more weight (P<.05) than CAS rats after 16 weeks only. The SPI rats had lower liver steatosis scores after 8 and 16 weeks (P<.05 and P<.001, respectively) compared to CAS and ARG rats. SPI rats had lower serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels (P<.05) compared to CAS after 16 weeks, and AST was lower (P<.05) compared to ARG rats. After 16 weeks, the SPI rats had lower (P<.05) serum ALT and AST levels than at 8 weeks. Our results suggest that a longer period of SPI feeding results in lower liver steatosis and serum ALT and AST levels, while the ARG diet had no effect on steatosis or ALT and AST levels. We found that the SPI diet reduced (P<.001) serum tumor necrosis factor-α (TNF-α) compared to CAS and ARG diets after 8 and 16 weeks. The SPI diet significantly reduced (P<.001) interleukin-6 (IL-6) when compared to the CAS diet at 8 weeks, but there was no significant difference at 16 weeks. Based on the findings of our study, the protective effect of SPI in reducing liver steatosis is not modulated by its native arginine content.
Introduction
O
Fatty liver disease can range from the fatty liver alone (liver steatosis) to fatty liver associated with inflammation (steatohepatitis). This condition can occur with the use of alcohol (alcohol-related fatty liver) or in the absence of alcohol (nonalcoholic fatty liver disease [NAFLD]). NAFLD is one of the common causes of liver diseases with a prevalence of up to 34% in the United States, 5,6 but this prevalence increases to more than 50% in the obese population. NAFLD has been associated with obesity, type 2 diabetes, and hyperlipidemia. 5 It is known now that NAFLD patients can develop progressive liver disease leading to end-stage liver disease and hepatocellular carcinoma. 6 In one cohort study, patients with NAFLD had a higher mortality rate compared to the general population. 7
Obesity leads to insulin resistance, and the increase in adipose tissue mass is often accompanied by an increase in inflammation caused by the infiltration of macrophages into adipose tissue. The accumulation of macrophages correlates with the increase in circulating inflammatory markers such as tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6), which also correlate with insulin resistance and metabolic syndrome. 8
Several studies have reported that consumption of soy protein can decrease serum cholesterol and triglycerides and also reduce accumulation of cholesterol and triglycerides in the liver, which can lead to reduction in or absence of liver steatosis. 9 –13 Soy protein is a rich source of bioactive molecules (e.g., isoflavones and arginine [ARG]), which suggests that these components may be integral in soy protein's lipid-modulating abilities. 14,15 Understanding which bioactive molecules are responsible for the potential benefits of soy protein is of importance, as a specific intrinsic lipid-lowering mechanism for soy protein has not been clearly delineated. Recent findings demonstrated that soy protein containing isoflavones reduced liver lipids, which resulted in reduction of cholesterol, and that adding unconjugated soy isoflavones (genistein and daidzein) to a casein (CAS) diet lowered the hepatic concentration of triglycerol compared to the casein diet alone. 16 –18 Several studies have indicated that isoflavones in soy protein are responsible, at least in part, for the reduction of liver steatosis. 17 –19
Arginine is a conditionally essential amino acid and a precursor for nitric oxide synthesis. Several studies have shown that arginine can promote fat oxidation and decrease fat synthesis in the liver. 20,21 In addition, another study using obese diabetic fatty Zucker rats found that dietary supplementation of arginine in drinking water reduced fat mass by enhancing expression of fatty acid and glucose oxidation. 22
Soy protein is rich in arginine, which may account for some of its potential beneficial effects on liver metabolism. We have recently shown that obesity increases liver steatosis, and long-term soy protein isolate (SPI) feeding reduced the liver steatosis in the rat dimethylbenz[α]anthracene (DMBA)-induced mammary tumor model. 23 The objective of this study was to investigate the effects of short- and long-term feeding of an SPI diet (naturally rich in ARG) on reducing liver steatosis in male obese Zucker rats. We hypothesized that the addition of ARG to a casein diet (control diet) at a level equivalent to that in the SPI diet may, in part, protect from the development of fatty liver disease in obese Zucker rats.
Materials and Methods
Experimental design
All animal protocols were approved by the Institutional Animal Care and Use Committee at the University of Arkansas for Medical Sciences.
Forty-eight 6-week-old male Zucker rats (obese fa/fa) were purchased from Harlan Laboratories (Indianapolis, IN, USA). After 1 week of acclimation, rats were randomly assigned to three different dietary groups (n=16/group) and had access ad libitum to water and to one of three diets. Rats were housed one per cage. All rats were weighed twice per week, and the food intake was not monitored. We used a semipurified AIN-93G diet (20% protein) (Harlan Laboratories, Madison, WI, USA) as the base. The base diet was prepared with either CAS or SPI with naturally occurring isoflavones as the dietary protein source, as previously reported. 23 The base diet prepared with CAS had 6 g arginine/kg diet, and the base diet prepared with SPI had 13 g arginine/kg diet. For the ARG diet, another 7 g arginine/kg diet (6+7 g=13 g) was added to match the arginine concentrations to the SPI diet. The diet groups are as follows: CAS (6 g arginine/kg diet, 0.6% ARG), CAS with ARG (13 g arginine/kg diet, 1.3% ARG, equal to levels in SPI diet), or SPI (13 g arginine/kg diet, 1.3% ARG). The isoflavone content of the SPI used was as follows: 1.87 mg isoflavones/g protein (expressed as aglycone equivalents) with 66% as genistein and 29% as daidzein. In each dietary group, rats received their respective diet for either 8 weeks (short term) or 16 weeks (long term), and at the end of each experiment (8 or 16 weeks), eight rats from each dietary group were sacrificed. Livers were weighed, and liver samples were taken for histopathological evaluation as described below. In addition, serum samples were collected to measure the levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), and different adipokines (TNF-α and IL-6).
Liver histology
A board-certified anatomic pathologist (Soheila Korourian) evaluated livers in a blinded protocol. Two 3-mm sections of each lobe of the liver were fixed in 10% buffered formalin for histological examination. Liver sections were fixed in 5% buffered formalin, cut, and stained with hematoxylin and eosin. Liver sections were evaluated for the presence of microvesicular and macrovesicular steatosis. The steatoses were semiquantitated as a score of 1–4 in each case, as previously described. 24
Serum measurements
Serum ALT and AST were measured using the VetScan VS2 profile. The analysis uses an immunoassay based on the measurement of enzyme mass in the serum. The kit uses antibodies directed to rat ALT or AST, which are detected using 20 μL of serum using triplicate measurements. Serum TNF-α and serum IL-6 were measured using ELISA Kits (R&D Systems, Inc., Minneapolis, MN, USA) according to the manufacturer's instructions.
Statistical analyses
Statistical analyses were performed as previously reported. 25,26 Descriptive analysis was performed, and statistics were summarized in graphs. Analyses of variance followed by t-tests were carried out and P-values were reported. Both parametric and nonparametric statistical procedures were used based on the distribution of variables. Statistical significance was set at P<.05 (two sided) for the individual test, and Bonferroni adjustment was applied in the significance level for multiple comparisons. Data preparation and analyses were performed using SAS software (version 9.2; SAS Institute, Inc., Cary, NC, USA).
Results
Body and liver weights
All rats gained weight during the course of the 8- and 16-week experiments. The average body weights (mean±standard deviation) at the midpoint and end of the experiment are shown in Figure 1 and Table 1. There was no significant difference among the body weight of rats fed SPI and ARG compared to the CAS diet at the end of the 8-week feeding. However, the rats fed SPI and ARG gained more weight (P<.05) than CAS rats after 16 weeks. Comparing the short-term (8 weeks) versus long-term (16 weeks) dietary groups, all rats gained significantly (P<.001) higher body weight in their respective diets over a period of time (8 weeks vs. 16 weeks). The liver weights expressed as absolute weight or as a percentage of body weight are shown in Figure 2A and B and Table 1. Obese rats fed the SPI diet had significantly lower liver weight (expressed as absolute liver weight or as a percentage of the body weight), both after 8 and 16 weeks (P<.001), compared to obese CAS- and ARG-fed rats. However, obese CAS- and ARG-fed rats had a significantly higher liver weight and the SPI diet lowered liver weights.
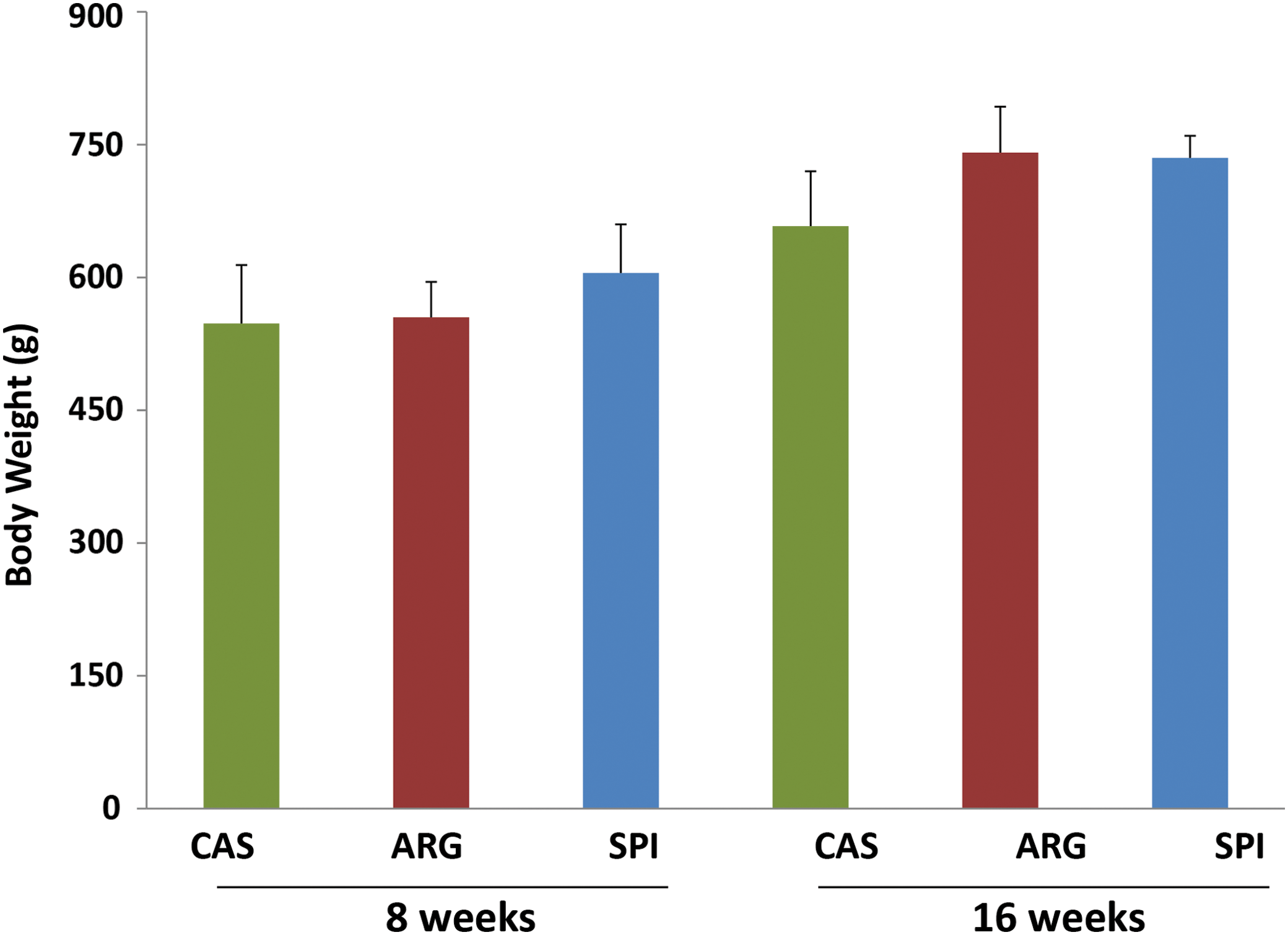
Body weight of obese rats fed casein (CAS), arginine (ARG), and soy protein isolate (SPI) at the end of 8 or 16 weeks of feeding. Data are presented as mean±standard deviation (SD). Color images available online at
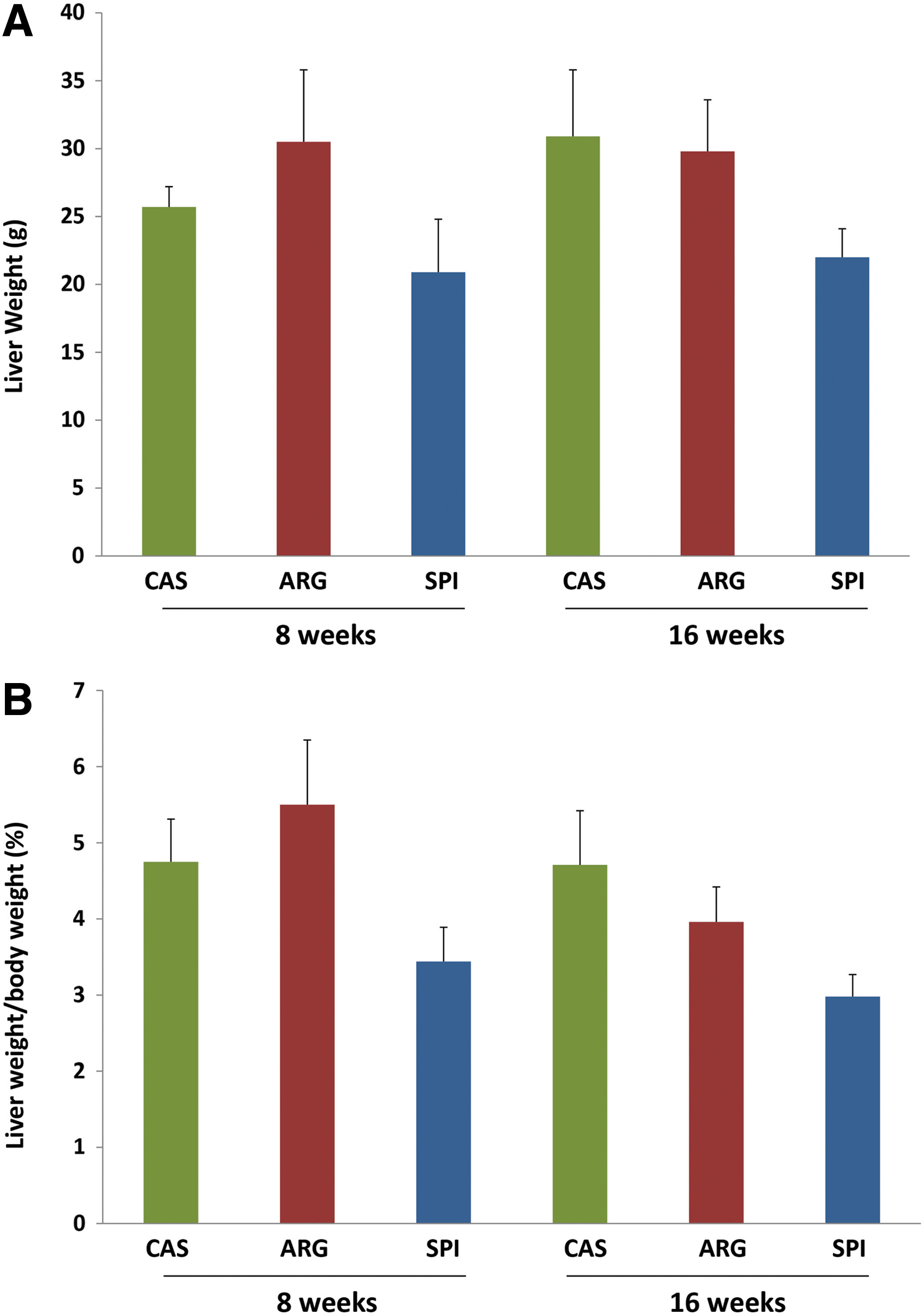
CAS, ARG, and SPI at the end of 8 or 16 weeks of feeding.
ALT, alanine aminotransferase; ARG, arginine; AST, aspartate aminotransferase; CAS, casein; IL-6, interleukin-6; SPI, soy protein isolate; TNF-α, tumor necrosis factor-α.
Liver pathology
Representative photomicrographs of the hepatic parenchyma for CAS, ARG, and SPI diets after 8 and 16 weeks are shown in Figure 3A. The average scores for liver steatosis (mean±standard error) at the end of the 8- and 16-week experiments are shown in Figure 3B and Table 1. At the end of 8 weeks, the SPI-fed rats had a significantly lower (P<.05) liver steatosis score (2.10±1.0) than CAS-fed (3.20±0.5) and ARG-fed (3.40±0.7) rats, and there was no significant difference between CAS- and ARG-fed rats. At end of the 16-week experiment, the SPI-fed rats had a significantly lower (P<.001) liver steatosis score (1.70±0.7) than CAS-fed (3.4±0.9) and ARG-fed (3.10±0.6) rats, and there was no significant difference between CAS- and ARG-fed rats. Our results show that the SPI-fed rats had lower liver steatosis scores after 8 and 16 weeks (P<.05 and P<.001, respectively) compared to CAS- and ARG-fed rats.

Serum ALT and AST
At the end of the 8-week experiment, the SPI-fed rats had lower, not significant, serum ALT (P=.08) and AST (P=.06) compared to ARG-fed rats (Fig. 4A, B and Table 1). However, after 16 weeks of feeding, SPI-fed rats had lower serum ALT and AST levels (P<.05) compared to the CAS-fed group. In addition, the SPI-fed rats had lower AST levels (P<.05) compared to ARG rats. When comparing the longer period of SPI feeding (16 weeks vs. 8 weeks), the rats fed SPI for 16 weeks had lower (P<.05) serum ALT and AST levels than rats fed SPI for 8 weeks.

Serum TNF-α and IL-6
Rats fed the SPI diet had reduced (P<.001) serum TNF-α concentration compared to the CAS and ARG diet after 8 and 16 weeks (Fig. 5A, B and Table 1). In addition, SPI diet consumption significantly reduced (P<.001) the IL-6 concentration compared to the CAS diet at 8 weeks, but not at 16 weeks.

Discussion
Zucker rats (fa/fa) are the best known, most widely used rat model for genetic obesity and have been used to study noninsulin-dependent diabetes mellitus. 17 –19,27,28 Animals homozygous for the fa allele become noticeably obese by 3–5 weeks of age, and by 14 weeks of age, their body composition is more than 40% lipid. 29 The most valuable contribution of the Zucker rat has been its utility as a model of human, early-onset hyperplastic–hypertrophic obesity. 30 –34 In addition, this rat model develops hepatic steatosis due to dysregulated metabolic gene expression in the liver, as is seen in humans, 28 making it suitable for studying fatty liver disease. 35
Our current study findings as well as those of our colleagues demonstrated that SPI consumption reduced liver steatosis in the obese Zucker rat model with 16-week (long term) SPI consumption, 23,36 but little is known about the inhibitory effect of SPI consumption on liver steatosis in the more early stages (e.g., 8 weeks). As soy protein is rich in arginine, we anticipated that the potential beneficial metabolic effects of soy protein-derived arginine 37 –40 may play a role in soy protein's protective effect to reduce fatty liver. Therefore, we examined the effects of short- and long-term consumption of SPI and ARG diets on reducing liver steatosis in the present study.
The major objectives of this study were to investigate the effects of short- and long-term ARG and SPI feeding of male obese Zucker rats on (1) liver steatosis, (2) liver damage markers, such as serum ALT and AST, and (3) serum inflammation markers TNF-α and IL-6.
In the present study, we found that SPI-fed rats had lower liver steatosis after 8 and 16 weeks (P<.05 and P<.001) compared to CAS- and ARG-fed rats. In addition, SPI-fed rats had lower serum ALT and AST levels (P<.05) compared to CAS after 16 weeks, suggesting less impaired liver function in the SPI-fed group. Our results also showed that the SPI diet reduced markers of inflammation, namely, serum TNF-α (P<.001) compared to the CAS and ARG diet after 8 and 16 weeks. Consumption of the SPI diet also resulted in lower IL-6 compared to the CAS diet in 8 weeks (P<.001), but not after 16 weeks. This is the first report to examine the levels of arginine present endogenously in the SPI diet or free arginine added to a CAS diet on liver steatosis. Our results suggest the important roles of arginine in the SPI diet on body weight, as both SPI and ARG long-term feedings significantly increased body weight to the same extent relative to the control CAS-fed group (Fig. 1). This observation suggests that the arginine levels in the SPI diet may be responsible for the body weight increase, since it has been shown that arginine, a conditionally essential amino acid, has a positive body weight gain effect. 38 –40
From our study, SPI but not ARG feeding significantly reduced the liver steatosis score (Fig. 3A, B). The inhibitory effect of SPI feeding on liver steatosis is consistent with a previously reported study using soy protein-fed obese Zucker rats, which showed that SPI feeding resulted in increased body weight gain, but lower liver steatosis. 32 Other similar studies have reported using long periods of SPI feeding, which lowered the steatosis, but we investigated a shorter period (8 weeks) of SPI feeding that also resulted in lower liver steatosis. Previous studies from our laboratory in the rat DMBA-induced mammary tumor model demonstrated that obesity increases liver steatosis and that long-term soy protein feeding also reduced the liver steatosis in that model. 23 In the present study, we found that arginine alone added to a casein diet, as the level found in the SPI diet did not reduce liver steatosis. Despite the observation that the SPI-fed rats had higher body weights, we found that liver weights, as absolute percentages of body weight, were lower compared to ARG- and CAS-fed rats. As SPI is an isoflavone-rich diet, these observations are consistent with the fact that isoflavones prevent fatty liver disease through the regulation of peroxisome proliferator-activated receptors, carbohydrate responsive element-binding protein, and Wnt signaling to control fatty acid β-oxidation, lipid synthesis, and oxidative stress. 41,42 Other studies have shown that feeding single-cell protein (SCP) with low methionine:glycine and lysine:arginine ratios to obese Zucker rats demonstrates hypercholesterolemic effects. 16,19 In a more recent study, the same investigators demonstrated that SCP feeding reduced hepatic steatosis and lowered plasma transaminases levels. They concluded that increased β-oxidation of fatty acids after SCP feeding may have contributed to the reduction in liver steatosis combined with the change in fatty acid composition, which directed fatty acids toward oxidation rather than esterification to triacylglycerols. 43 In addition, SCP feeding resulted in a reduced hepatic clearance of circulating very-low-density lipoprotein, which may also have contributed to decreased liver fat accumulation. 43
To further understand the functional roles of ARG and SPI feeding, we examined their effects on key markers of liver function. Elevated AST or ALT levels are always present in NAFLD. It has been shown that obesity increases the serum ALT and AST levels. 44 In the present study, we have shown that long-term SPI feeding reduced both serum ALT and AST levels, which demonstrates a potential role for the SPI diet in effectively reducing liver steatosis. Gudbrandsen reported that adding soy isoflavones (genistein and daidzein) to the casein diet lowered the hepatic concentration of triglycerol and decreased plasma AST and ALT levels compared to a casein diet alone. 18 These data suggest that isoflavones may be responsible for the decreased liver enzyme concentrations.
TNF-α and IL-6 are proinflammatory cytokines and stimulate hepatic collagen-producing stellate cells during steatohepatitis. 45 The current study demonstrates for the first time that SPI feeding greatly reduced proinflammatory cytokine TNF-α (short and long term) and IL-6 (short term) compared to CAS or ARG feeding in the Zucker fatty rat model. These results provide a new line of evidence that the reduction of proinflammatory cytokines (e.g., TNF-α and IL-6) by SPI feeding, at least in part, is associated with a reduction in liver steatosis. Interestingly, we also found that long-term ARG feeding significantly decreased the level of AST, but not the levels of ALT, TNF-α, and IL-6. This finding is important because it indicates that arginine in SPI or ARG may have some effect on maintaining normal liver function (decreased AST), but arginine feeding alone could not reduce liver steatosis.
In summary, we demonstrated that short- and long-term soy protein consumption can decrease liver steatosis and markers of liver damage and can reduce the expression level of inflammatory cytokines (e.g., TNF-α and IL-6) in the obese Zucker rat model. The protective effect of SPI to reduce liver steatosis does not appear to be due to its relatively enriched arginine content.
Footnotes
Acknowledgments
This study was supported by a grant from the Arkansas Biosciences Institute. The authors wish to thank Phaedra Yount, John Gregan, and Heather Johnston for valuable assistance in preparation of this article. DuPont Nutrition & Health provided the soy protein isolate used to prepare the diets.
Authors' Contributions
R.H. applied for and received funding for this project, designed and supervised the study, and participated in collection of liver and serum data and writing of this article. H.Z. participated in experiments, interpretation of study results, and in writing of this article. S.K. provided expertise in classifying and analyzing all of the liver pathology, interpreting study results, and writing of this article. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.