
Abstract
Food formulation with bioactive ingredients is a potential strategy to promote satiety and weight management. Whey proteins are high in leucine and are shown to decrease hunger ratings and increase satiety hormone levels; cocoa polyphenolics moderate glucose levels and slow digestion. This study examined the effects of cocoa and whey proteins on lipid and glucose metabolism and satiety in vitro and in a clinical trial. In vitro, 3T3-L1 preadipocytes were treated with 0.5–100 μg/mL cocoa polyphenolic extract (CPE) and/or 1–15 mM leucine (Leu) and assayed for lipid accumulation and leptin production. In vivo, a 6-week clinical trial consisted of nine panelists (age: 22.6 ± 1.7; BMI: 22.3 ± 2.1) consuming chocolate-protein beverages once per week, including placebo, whey protein isolate (WPI), low polyphenolic cocoa (LP), high polyphenolic cocoa (HP), LP-WPI, and HP-WPI. Measurements included blood glucose and adiponectin levels, and hunger ratings at baseline and 0.5–4.0 h following beverage consumption. At levels of 50 and 100 μg/mL, CPE significantly inhibited preadipocyte lipid accumulation by 35% and 50%, respectively, and by 22% and 36% when combined with 15 mM Leu. Leu treatment increased adipocyte leptin production by 26–37%. In the clinical trial, all beverages significantly moderated blood glucose levels 30 min postconsumption. WPI beverages elicited lowest peak glucose levels and HP levels were significantly lower than LP. The WPI and HP beverage treatments significantly increased adiponectin levels, but elicited no significant changes in hunger ratings. These trends suggest that combinations of WPI and cocoa polyphenols may improve markers of metabolic syndrome and satiety.
Introduction
F
Nutrient metabolism and weight gain are influenced by a variety of protein and peptide hormones and biomarkers. Of particular interest are the adipocyte-derived proteins leptin and adiponectin. Leptin increases satiety, or the feeling of fullness between meals, and impacts long-term weight management. 7 Adiponectin regulates lipid and glucose metabolism and attenuates insulin resistance and development of Type II diabetes. 7 Research suggests that diets consisting of 30% or greater calories from protein, and more specifically whey protein, are most effective in inducing satiety and reducing food intake. 1 When compared to animal and plant proteins, whey protein has been demonstrated to increase ratings of fullness, satiety hormone levels, leptin levels, and to decrease ad libitum intake. 8 –10 The satiety-inducing benefits of whey proteins are attributed to their high branched-chain amino acid content, specifically leucine. 3 In vitro, in vivo, and in clinical trials, leucine treatments are shown to increase muscle synthesis, increase plasma leptin levels, decrease plasma lipid levels, and improve body composition. 11 –13 Research also suggests that meals containing leucine or whey protein decrease plasma glucose, increase production of insulin and adiponectin, and may attenuate insulin resistance. 3,4,14 –17
Polyphenolic compounds found in cocoa, fruits, and tea have also been shown to increase satiety, insulin sensitivity, and lipid metabolism. 18,19 In preadipocytes, 100 or 200 μg/mL cocoa polyphenol extract inhibited adipogenesis by 30% and 72%, respectively, 20 while 15–30 μg/mL (-)-catechin increased adiponectin production by over 200%. 21 Cocoa tannins, catechins, and procyanidins effectively slowed digestion in vitro by binding digestive enzymes. 22,23 In mouse and human clinical studies, flavanol-rich cocoa or chocolate consumption decreased fat accumulation and fatty acid synthesis, 20 increased insulin sensitivity, 24 and lowered short-term blood glucose and free fatty acid levels. 18,25 Epicatechin consumption increased insulin levels and decreased diabetes rates by 50% in mice whereas dark chocolate consumption decreased hunger ratings and ad libitum intake by 17% and increased satiety more than milk chocolate. 26,27
Previous investigations have shown the individual benefits of polyphenolics and whey protein. However, the combination of these two popular ingredients could provide an affordable, enjoyable, and effective approach to increase satiety and aid in weight loss or weight maintenance. The present study is the first to examine the combined health effects of cocoa polyphenolics and whey proteins and aimed to evaluate the effects of these ingredients upon lipid and glucose metabolism and markers of satiety.
Materials and Methods
Preparation of cocoa polyphenolic extract and flavanol analysis
Extraction of cocoa polyphenols was performed as by Adamson et al. 28 Briefly, cocoa powder (Hershey's, Hershey, PA, USA) was defatted and phenolics were extracted with 70% acetone (Sigma-Aldrich, St. Louis, MO, USA). Supernatant was filter sterilized and stored at −20°C. Flavanol content of cocoa polyphenolic extract (CPE) and raw cocoa was characterized by The Hershey Company: flavanol monomers were determined through LC/APCI-MS, 29 flavanol dimers through decamer polymers by LC/MS, 30,31 and total flavanols using the colorimetric DMAC (4-dimethylaminocinnamaldehyde) method as previously described. 32 Tables 1 and 2 provide the flavanol profiles of CPE and cocoa powders, respectively.
Analyses performed by The Hershey Company.
Analyses performed by The Hershey Company.
Determined through LC/APCI-MS and LC/MS analysis.
Determined through 4-dimethylaminocinnamaldehyde (DMAC) analysis.
Cell culture
3T3-L1 preadipocytes were purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured at 37°C and 5% CO2 in standard culture medium of Dulbecco's modified Eagle's medium fortified with 10% fetal bovine serum (Sigma-Aldrich). Cells were seeded into 96-well plates at a density of 50,000 cells per well and grown to confluence. Cells were differentiated as by Phrakonkham et al. 33 Briefly, cell differentiation was induced with 0.5 mM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), 0.25 μM dexamethasone (Sigma-Aldrich), and 170 nM insulin (Santa Cruz Biotechnology, Dallas, TX, USA) treatment for 48 h (day 0–2) followed by 170 nM insulin for 48 h (day 2–4). At day 4, differentiation media were removed and replaced with standard culture medium for 2 days until cells were visually estimated to be 80–90% differentiated. CPE was incorporated into the standard culture medium at 0.5–100 μg/mL before treatment using acetone as a solvent; cells were treated in triplicate with 0.5–100 μg/mL CPE and/or 1–15 mM Leu from days 0 to 2 or for 48 h after complete differentiation (days 6–10). Cell viability was verified by MTT assay as previously reported. 34 Controls included a negative control (NC) of standard culture medium and a negative control plus solvent (NCS) of culture medium supplemented with 100 μg/mL acetone extraction solvent. NCS distinguished between the effects of the CPE solvent and the cocoa polyphenolics. All CPE treatments contained equal concentrations of extraction solvent to mitigate any solvent effects.
Lipid accumulation was quantified using an Oil-Red-O (Sigma-Aldrich) assay as previously reported. 35 Following 48 h of treatment postdifferentiation, growth media was removed and assayed for leptin content with a Mouse Leptin ELISA Kit (Sigma-Aldrich) according to the manufacturer's instructions.
Clinical trial
Experimental design
This trial was a randomized, single blind crossover study comprising of six four-hour sessions. Panelists participated in one session per week for 6 weeks; for each session, panelists fasted overnight and arrived at the laboratory at 7 am. Upon arrival, panelists completed a baseline hunger questionnaire and used a sterile lancet (Fisherbrand, Waltham, MA, USA) to acquire finger-prick blood samples. Panelists determined baseline blood glucose levels using a commercial Nova Max Plus glucose/ketone monitor (Waltham, MA, USA) and provided 200 μL of blood into a blood collection tube (Becton-Dickinson, Franklin Lakes, NJ, USA). Panelists then consumed a randomized 340 g beverage and ranked beverage hedonic liking, provided blood samples, tested blood glucose levels, and ranked hunger levels at 0.5, 1, 2, and 4 h following beverage consumption. Analyses for serum adiponectin levels were performed using human ADIPOQ/Adiponectin ELISA Kits (Sigma-Aldrich) as per the manufacturer's instructions.
Beverage formulation
Beverage nutritional analysis is outlined in Table 3, including a placebo, whey protein isolate (WPI) (Davisco, Inc., Le Sueur, MN, USA), Dutch-process low polyphenolic cocoa (LP) (Hershey), high polyphenolic cocoa (HP) (Hershey), LP + WPI, and HP + WPI. ADM maltodextrin (Decatur, IL, USA) was used as a direct replacement for WPI and ADM Fibersol-2, a resistant maltodextrin, was used to replace cocoa. All cocoa-containing beverages (LP or HP) provided 36 g of cocoa. All beverages provided 130–150 calories.
Proanthocyanidin oligomers represent monomer to decamer polyphenolic polymers (1–10 monomer units).
Analyses performed by The Hershey Company.
CHO, carbohydrates; WPI, whey protein isolate; LP, low polyphenolic cocoa; HP, high polyphenolic cocoa.
Subjects
All procedures were approved through the North Carolina State University Institutional Review Board (FWA00003429). A total of 9 panelists were recruited based upon the following criteria: men and women between 18 and 35 years of age of healthy weights (body mass index between 18 and 25 and/or waist to hip circumference ratio of ≤0.95 for men and ≤0.8 for women), not dieting, no loss or gain of significant weight in the last year, had not recently started or ceased a regular exercise program, no dietary allergies to dairy or cocoa ingredients, and no history of Type I or Type II diabetes. Baseline panelist information is listed in Table 4.
Values are mean ± SD.
Questionnaire
At each time point, panelists recorded blood glucose levels and rated feelings of hunger on a 100 mm horizontal visual analogue scale validated by Flint et al. for single test meal studies. 36 The visual analogue scale was anchored at both ends with qualifying statements: 0 mm with “not at all hungry” and 100 mm with “extremely hungry.”
Statistical analysis
For cell culture, a one-way ANOVA for Tukey's honestly significant differences was applied to make all pairwise comparisons among treatment means. For the clinical trial, panelist sample order was randomized with a block design in JMP (Cary, NC, USA). The experimental design was a randomized complete block split plot in time, where panelists served as complete blocks and beverages as the whole plot blocks, measured repeatedly in time. The linear mixed effects model was fit with all factorial effects: beverage, time, and all possible interactions and random effects for panelist and panelist-by-beverage. Statistical analysis utilized SAS software (Cary, NC, USA) and was performed at an error rate of 0.05.
Results
Cell culture
CPE treatment inhibits preadipocyte differentiation and decreases lipid accumulation in vitro
Figure 1 compares the effects of CPE and combined CPE/Leu treatments upon preadipocyte and adipocyte lipid accumulation. Preadipocyte treatment of 50 or 100 μg/mL CPE significantly decreased cellular lipid accumulation by 35% and 50%, respectively, while CPE + 15 mM Leu reduced lipid accumulation by 22–36%. Preadipocyte treatment with Leu did not significantly affect lipid content (results not shown). Mature adipocyte treatment with 10 and 100 μg/mL CPE significantly decreased cellular lipid accumulation by 19% and 16%, respectively. As shown by Oil-Red-O staining in Figure 2, CPE treatment inhibited preadipocyte lipid accumulation in a dose-dependent manner. MTT assays indicated that the CPE and Leu treatments had no significant effects upon cell viability (Fig. 3).
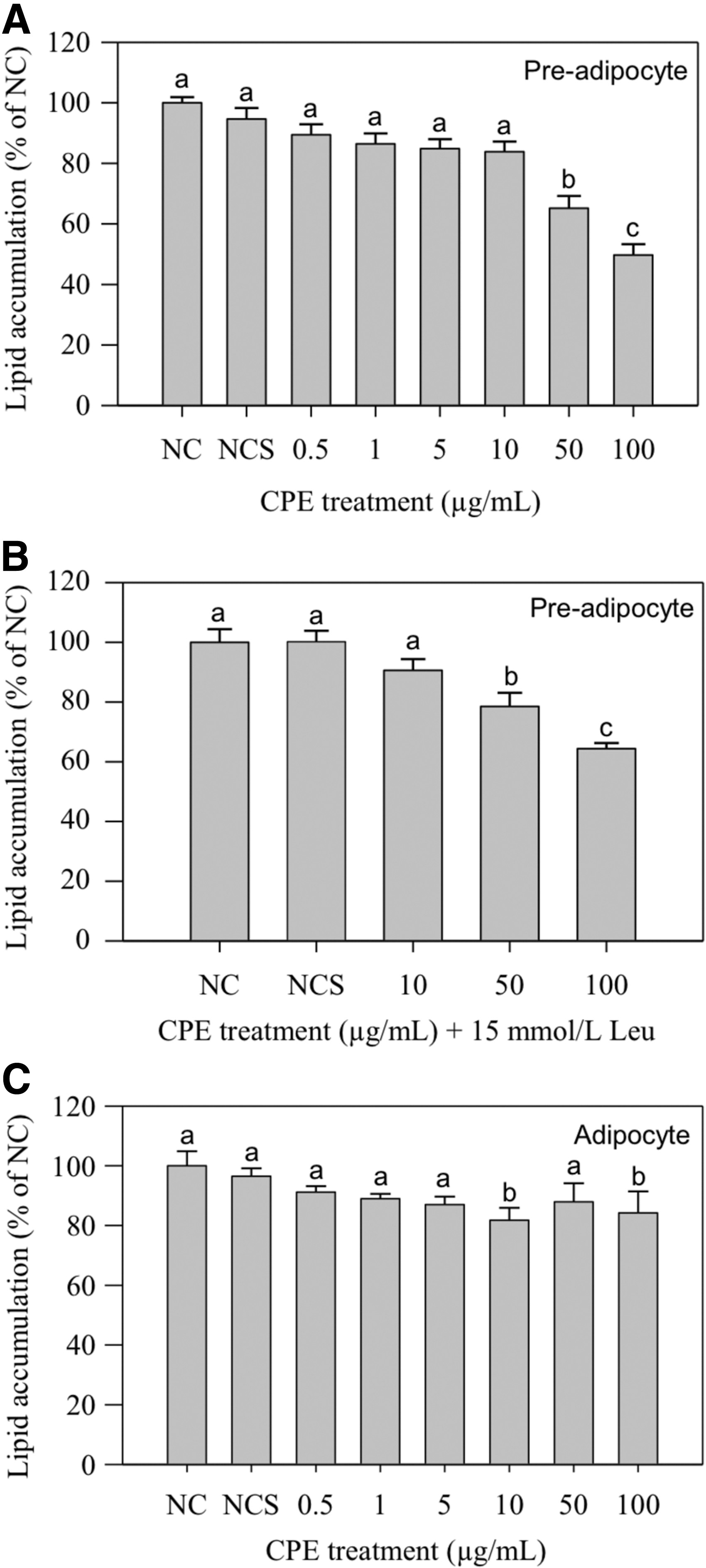
Effects of CPE and Leu treatment on lipid accumulation in 3T3-L1 preadipocytes and adipocytes. Data expressed as percent of negative control in mean ± SE. Preadipocytes were treated for 48 h during differentiation with
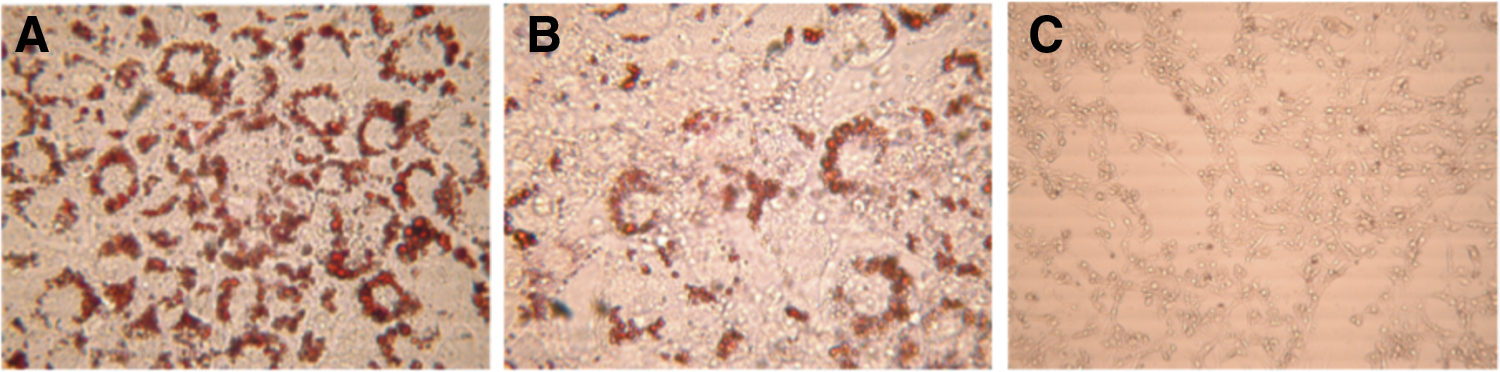
Effects of CPE on 3T3-L1 preadipocyte lipid accumulation following ORO lipid staining in
Cell viability as determined by MTT assay. Data expressed as percent of negative control in mean ± SE. Preadipocytes were treated for 48 h during differentiation with
Leu increases leptin secretion in vitro
Leptin controls included a negative control (NC), negative control + solvent (NCS), and a positive control + solvent (PCS) containing 0.5 μM insulin, which upregulates leptin production. 37,38 Leu treatments of 1–15 mM represented growth media amino acid supplementation of 3.5–50% and corresponded with protein supplementation levels reported in published in vivo and clinical trial studies. 39 –41 Leu treatments of 1–15 mM significantly upregulated leptin secretion by 26–37% (Fig. 4B). CPE treatment of 0.5 μg/mL increased leptin secretion by 25% over NCS; however, this upregulation was not statistically significant (Fig. 4A). Higher CPE treatments significantly decreased leptin production by 78% below that of NC. CPE + Leu treatments significantly decreased leptin production, whereas NCS appeared to upregulate leptin production.
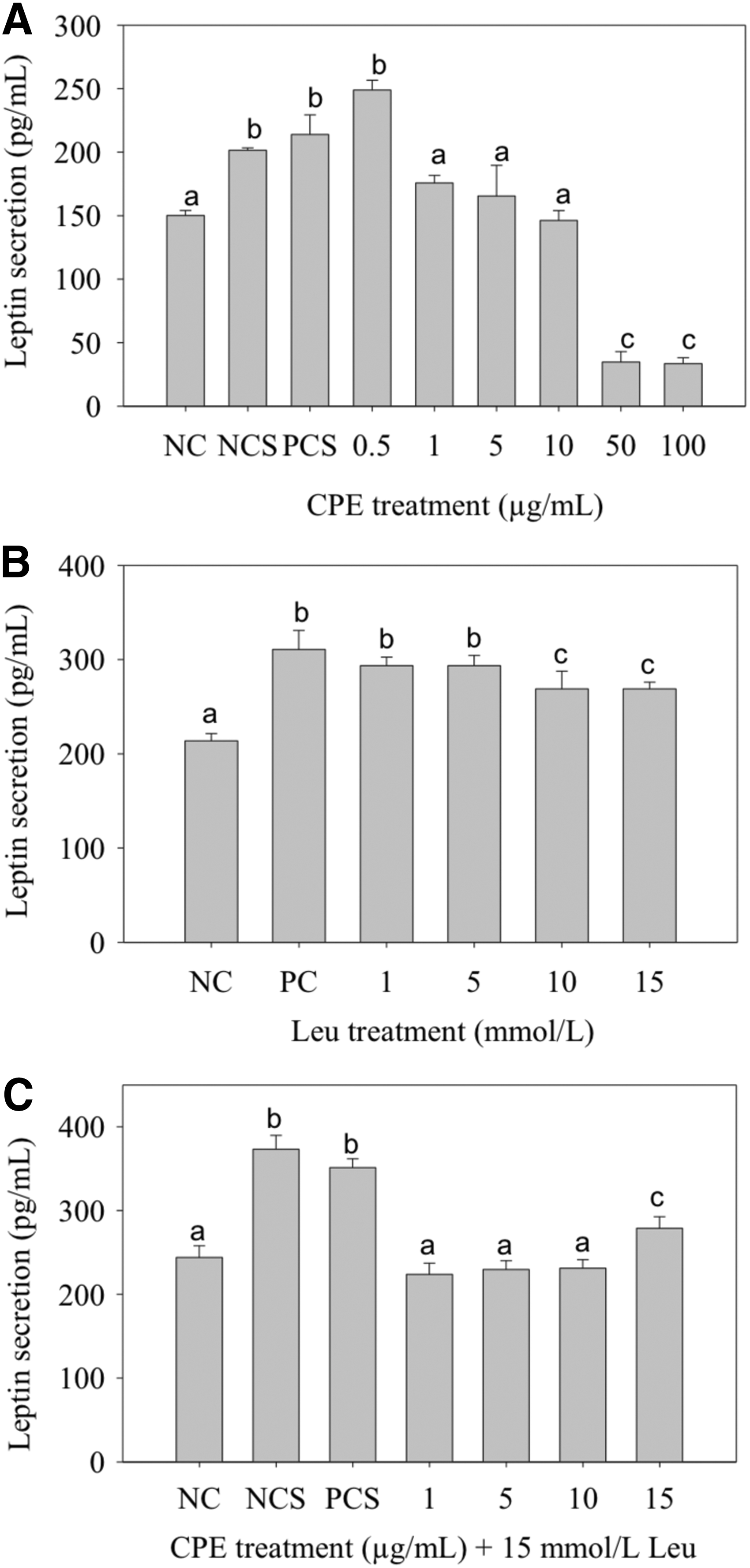
Effects of CPE and Leu treatment on 3T3-L1 adipocyte leptin production. Data expressed as mean ± SE. Adipocytes were treated for 48 h postdifferentiation with
Clinical trial
High polyphenolic cocoa and whey protein moderate blood glucose levels
Figure 5 compares the effects of cocoa and whey protein beverages upon panelist blood glucose levels, hunger ratings, and serum adiponectin levels. The WPI, HP-WPI, and LP-WPI beverages increased blood glucose 8%, 12%, and 16% above baseline levels, respectively, compared to increases of 27%, 39%, and 58% in HP, LP, and placebo beverages, respectively (Fig. 5A). The presence of WPI significantly lowered glucose levels 30 minutes postconsumption. HP significantly decreased the 30-min spike in blood glucose as compared to LP; no significant difference was found between HP-WPI and LP-WPI.

Changes in
Effects of cocoa and whey on panelist hunger ratings
All hunger ratings were significantly decreased relative to baseline at the 30- and 60-min time points (Fig. 5B). However, no significant differences were found in the overall hunger ratings (hours 0–4) among beverage treatments, nor between cocoa or whey variables. Although not significant, the WPI and HP-WPI beverages elicited lower hunger ratings than those of other beverages while the cocoa-containing beverages exhibited the highest hunger ratings 4 h postconsumption.
Cocoa and whey increase serum adiponectin levels
All WPI and cocoa beverages increased serum adiponectin levels above baseline, whereas the placebo lowered levels below baseline at all hours following consumption (Fig. 5C). Adiponectin concentrations reached peak levels between 30 and 60 min after test beverage consumption. The WPI and HP beverages elicited the greatest increases in adiponectin levels and were significantly higher than the placebo at 0.5–2 h and 1 h postconsumption, respectively. However, adiponectin levels between test beverages were not significantly different.
Discussion
The amino acid leucine is found in higher proportions in whey protein and has been shown to increase leptin signaling in vitro and satiety in vivo. 42 –44 Due to the inability of cells to uptake whole proteins, leucine treatments were used as a substitute for whey protein in vitro. Additionally, glucose sensitivity, which was measured in the clinical trial, could not be tested in vitro. The in vitro and in vivo inhibition of lipid accumulation by plant polyphenolics has been reported in the literature. 34,45 –47 Min et al. demonstrated that preadipocyte cocoa polyphenolic treatments of 100 and 200 μg/mL decreased cellular lipids by 30% and 72%, respectively, and decreased fatty acid synthase levels. 20 Epigallocatechin gallate and grape skin extract treatment have shown similar effects. 48 –50 The results of the present study exhibited an additional 20% decrease in lipid accumulation at 100 μg/mL CPE, likely due to differences in cocoa source and extract preparation. Mechanistically, cocoa polyphenolics have been shown to bind directly to the insulin receptor and to reduce downstream signals for cell proliferation, including PPAR-γ, PPAR-α, and C/EBP-α. 20,51 Proteins and polyphenols have been shown to interact through noncovalent hydrophobic binding, 52 which may explain the reduced effectiveness of Leu + CPE treatment. Neutral pH conditions may favor ionic interactions between the unprotonated carboxyl group of Leu and polyphenols, which may then hinder or alter the polyphenolic binding affinity for cellular receptors.
Our results indicated that preadipocytes were more sensitive to the effects of polyphenolic treatment than were mature adipocytes, corresponding with curcumin research conducted by Kim et al. 53 Decreasing both preadipocyte differentiation and adipocyte lipid accumulation may be instrumental to long-term weight maintenance. While CPE treatment had greater effects on preadipocytes, the decreases in lipid accumulation in both cellular states indicate that cocoa polyphenolics influence both mechanisms.
The effects of leucine upon leptin secretion were not as pronounced as those in previous studies. Adipocyte treatment with 5 mM leucine was reported to increase leptin secretion two- to five-fold within 4 h of treatment. 42,43 In vivo, Lynch et al. reported that a high leucine meal increased plasma leptin in rats three-fold, whereas leucine-deficient meals decreased leptin secretion by 40%. 44 The reduced effect observed here may be due to the treatment method: the present study supplemented complete growth media with leucine rather than administering leucine in the absence of most essential nutrients. 12,42 Our approach was designed to prevent cellular nutrient starvation and provided a more realistic representation for higher animal studies. However, this approach may have diminished the observed effects of leucine upon leptin secretion. CPE treatment and leptin secretion were inversely related, indicating that above a threshold cocoa polyphenol concentration, polyphenolics inhibited leptin secretion. As with adipogenesis, this may be due to the antagonistic binding of polyphenols to insulin receptors or to PPAR-γ, which are involved in the upregulation of leptin secretion. 20,54
In the clinical trial, WPI-containing beverages were formulated to contain 10 g of WPI, representing 29% of total calories. This dose provided an excellent source of protein and was designed to mimic supplementary snacks or commercial beverages that provide longer and more satisfying hunger control than other high calorie snack options. 55 The HP beverage also provided three to four times as many cocoa polyphenolics as competitive chocolate beverages. The use of healthy young subjects and a robust placebo (formulated with soluble fiber to replace cocoa) required greater differences between beverages for effects to be statistically significant. Leptin was not measured in this study due to research demonstrating its circadian nature: leptin levels increase very slowly throughout the day and peak up to 6 h following dinner. 56 For this reason, we chose instead to measure adiponectin and hunger ratings in the clinical trial.
Diets consisting of 30% or greater calories from protein are shown to be more effective in inducing satiety and decreasing food intake. 1 Nilsson et al. showed that matched carbohydrate drinks containing branched chain amino acids or whey protein decreased plasma glucose by 44–56% and increased insulin production by 31–60%. 14 Epicatechin and cocoa polyphenolic treatments have been shown to increase insulin sensitivity, glucose transport, and to moderate blood glucose levels in vitro, in vivo, and in clinical trials with diabetic subjects. 18,24,26,57 The variations in sugar content (8.0 g vs. 9.0 g for the lowest versus the highest levels) among the various beverages were a potential confounding factor in this study. While the placebo, which contained the highest sugar content (9.0 g) demonstrated the highest blood glucose response (Fig. 5A), the beverage with the second highest sugar content (8.7 g) produced one of the lowest glucose responses. For this reason, we believe that sugar content was not a major confounding factor. Overall, our results reinforced those of previous studies: WPI and HP significantly decreased spikes in blood glucose and increased adiponectin production.
In conclusion, in vitro and in a clinical trial, this study showed that cocoa polyphenolics alone, and in combination with whey proteins, prevent cellular lipid accumulation and boost lipid metabolism by increasing adiponectin levels. WPI-containing beverages significantly decreased spikes in blood glucose and high polyphenolic cocoa significantly moderated blood glucose levels when compared to low polyphenolic cocoa. While leucine increased satiety signaling in vitro, neither cocoa or whey protein had significant effects upon hunger ratings in the clinical trial. These null results were likely due to the limited size of this pilot study, the low calorie content of the beverage, or the use of a robust control. Further studies with larger sample sizes and higher calorie model foods are necessary to determine the physical and physiological effects of WPI and cocoa upon glucose metabolism and satiety. However, this study indicates that polyphenols may improve health by moderating glucose metabolism and increasing rates of lipid metabolism. Although only a pilot study, the trends observed indicate that combined cocoa and whey protein consumption may be a potential tool in the formulation of low calorie, satiety-inducing foods.
Footnotes
Acknowledgments
This study was supported by the NCSU Department of Food, Bioprocessing, and Nutrition Sciences and a grant (56178) from the Dairy Research Institute. The authors thank The Hershey Company and Davisco Foods International for their generous donation of cocoa powder and whey protein, respectively, for clinical trial beverage formulation. The authors would also like to thank Dr. Jason Osborne for his help in statistical analysis and Avi Aggarwal for her help with clinical trial data entry and analysis.
Author Disclosure Statement
No competing financial interests exist.