
Abstract
Growing concerns about food safety and environmental protection enhanced the need for new and safe plant disease control strategies. The chemical composition of the three essential oils (EOs) extracted from leaves and fruits of Schinus terebinthifolius and leaves of Schinus molle, growing in Tunisia, was studied by GC and GC-MS. In all, 12 compounds were identified. The oils were mainly composed of terpene compounds. α-Pinene, α-phellandrene, and
Introduction
T
Plant essential oils (EOs) could be successfully exploited as a source of alternative substances to synthetic pesticides, usable for controlling various infectious plant pathogens. Moreover, the EO antimicrobial efficacy has extended their use as a natural preservative for foodstuff conservation as well as for prolonging their shelf life. 7,8 The newly discovered EO antimicrobial properties showed the possibility of their use as natural biopesticides, which could be taken into account by the pharmaceutical industry to develop innovative and less risky therapies for some human diseases. 2,9
The cashew family Anacardiaceae Lindl. includes more than 700 species in 82 genera that are primarily distributed pantropically. Some of its genera are also present in temperate areas. Plants in this family are cultivated for their edible fruits and seeds as well as are a source of their medicinal compounds. From the chemical and economical points of view, they produce a good quantity of resins and tannins. Several species of the family belonging to genus Schinus are cultivated also as ornamentals. 10
The most common EOs of Anacardiaceae were extracted from the Schinus molle L. and Schinus terebinthifolius Raddi, native to Brazil and commonly known either as Brazilian pepper tree or as Felfel Aareed. They are indigenous to South and Central America and also grow in tropical and semitropical regions of the United States and Africa as ornamental garden trees. The above-mentioned species are characterized by production of an EO, which is responsible for a peppery flavor. In some places, they have also been used in the perfume industry. 11 The leaves and reddish fruits of S. terebinthifolius are rich in this EO that contains high concentrations of monoterpene and sesquiterpene hydrocarbons, as reported by El-Massry et al. 12
S. terebinthifolius was traditionally used as an antibacterial, antiviral, diuretic, digestive stimulant, tonic, wound healer, anti-inflammatory, and hemostatic as well as a medicament to treat urinary and respiratory infections. 13
El-Massry et al. 12 have studied the antimicrobial activity of different crude extracts, which have been obtained, using dichloromethane or ethanol, from fresh leaves of S. terebinthifolius cultivated in Egypt. The crude extract obtained using the first solvent exhibited higher antimicrobial activity against some fungi and bacteria, such as Staphylococcus aureus Rosenbach, Pseudomonas aeruginosa, Escherichia coli, Aspergillus niger, Aspergillus parasiticus, and Candida albicans, in comparison either to the crude extract obtained by ethanol or to the EO from the same source.
Ghanney and Rhouma 14 reported that crude leaf extracts of S. terebinthifolius, prepared with methanol, ethanol, and hot distilled water, had a clear antibiotic effect against Agrobacterium tumefaciens Smith & Townsend, the causal agent of crown gall disease in tomato plants. It is supposed that some alkaloid and flavonoid compounds in them could be responsible for the antibacterial activity.
dos Santos et al. 10 have chemically characterized the composition of two EOs extracted from the above two Anacardiaceae by capillary GC and GC-MS. Twenty-seven and 29 compounds were identified in EO obtained from S. molle and S. terebinthifolius, respectively. The same researchers showed that sesquiterpene and monoterpene hydrocarbons were present in high percentage in the leaf and fruit EOs obtained from both Schinus species.
Affonso et al. 15 analyzed the chemical composition of the EO isolated from fruits of S. terebinthifolius using chromatographic analysis by GC-MS and quantified the relative percentages of each component. In particular, 22 components, including monoterpene and sesquiterpene, were identified, with a yield of about 2.6% (w/w) of dry fruit weight. They also showed that its major constituents were α-fenchene (20.75%), β-pinene (10.11%), β-myrcene (9.30%), α-phellandrene (14.94%), limonene (20.81%), and isosylvestrene (13.87%). In previous studies, Barbosa et al. 16 reported that α-phellandrene and β-pinene represented, on average, 7.0% and 1.5%, respectively, of the EO isolated from fruits of S. terebinthifolius.
Generally, the major component of EO from leaves, flowers, and/or fruits of S. terebinthifolius resulted to be α-pinene (15.01–51.82%). 17 Barbosa et al. 16 reported the following major components of EO isolated from unripe fruits of S. terebinthifolius: α-cadinol (20.60%), δ-cadinene (15.48%), β-pinene (10.21%), and epi-α-muurolol (9.89%). Finally, Richter et al. 18 indicated α-pinene (16.9%), α-phellandrene (21.1%), β-phellandrene (10.8%), and limonene (23.7%) as the major constituents of EO from fruits of the same Schinus species.
Regarding the biological characterization, S. terebinthifolius EOs, Siddiqui et al. 19 have reported that they have antifungal, healing, and antiallergic effects. In some countries, S. terebinthifolius was used in folk medicine for treatment of inflammatory and venereal diseases as well as hemostatic, antirheumatic, pain relieving, antidiarrheic, and a remedy for gingivitis and fever. 15
Azoxystrobin is a fungicide acting, through the inhibition of mitochondrial respiration, on spore production and germination as well as mycelial growth. The same active substance has been shown to explicate a phytotoxic effect on some plants, such as certain apple and crab apple varieties. Azoxystrobin is toxic to fish and aquatic organisms. It is moderately persistent in soil, with a half-life of 1–4 weeks. Azoxystrobin may present a leaching risk to groundwater. 20
Due to the limited number of articles on the antimicrobial activity of EOs extracted from plants of genus Schinus, the current study has the goal to cast light on the composition and antimicrobial activities of EOs obtained from leaves and fruits of S. terebinthifolius and leaves of S. molle. The antifungal activity of the studied EOs and their minimum inhibitory concentrations (MIC) have been investigated against Botrytis cinerea Pers. and Colletotrichum acutatum J.H. Simmonds in comparison with the systemic fungicide azoxystrobin. Moreover, their antibacterial activity has been evaluated against three strains of Gram-positive (G+ve) bacteria (Bacillus megaterium de Bary, Bacillus mojavensis Roberts and Clavibacter michiganensis Smith) and four strains of Gram-negative (G−ve) bacteria (E. coli Migula, Xanthomonas campestris Pammel, Pseudomonas savastanoi Janse, and Pseudomonas syringae pv. phaseolicola Van Hall) compared to tetracycline antibiotic.
Materials and Methods
Fungal isolates
The tested phytopathogenic fungi were monoconidic isolates, stored at 4°C as pure cultures, and maintained in the mycotheca of the School of Agricultural, Forestry, Food and Environmental Sciences of the University of Basilicata, Potenza, Italy. The fungal species were cultured on potato dextrose agar (PDA) at 24°C ± 2°C. The micromycetes used were C. acutatum (isolate number 1778 from olive) and B. cinerea (isolate number 1132 from strawberry). Identification of the two studied isolates was reached on the basis of their microscopic morphological features and, successively, with molecular methods based on polymerase chain reaction only for the isolate from olive.
The total nucleic acids were extracted from pure cultures of isolate from olive with a commercial kit (QIAGEN, DNeasy Plant Mini Kit), according to the manufacturer's instructions. The DNA was amplified using the universal primer pair ITS4/ITS5. 21 The amplicons obtained were directly sequenced, and the resulting sequences were compared with those available in the GenBank nucleotide archive using BLAST software. 22
Bacterial isolates
The tested bacterial isolates were three strains of G+ve bacteria (B. megaterium, B. mojavensis and C. michiganensis) and four strains of G−ve bacteria (E. coli, X. campestris, P. savastanoi pv. savastanoi, and P. syringae pv. phaseolicola). All tested bacterial strains were cultivated on agar King B medium (KB), except for E. coli that was cultivated on Luria-Bertani medium (LB). All isolates were then incubated at 30°C ± 2°C for 48 h. All tested bacterial isolates have been previously identified and stored as pure freeze-dried cultures at −20°C in the collection present at the School of Agricultural, Forestry, Food and Environmental Sciences of the University of Basilicata, Potenza, Italy.
Studied EOs
All studied EOs were obtained from leaves and fruits of S. terebinthifolius and leaves of S. molle.
Plant material and extraction of EOs
For the extraction of studied EOs, the above plant organs were collected from the Arid Land Institute of Medenine (ALIM, southeast of Tunisia) and deposited in the Herbarium of the Dryland Farming and Oasian Crop Department of the same institute. Air-dried plant materials (200 g) were placed in a 5-L round-bottom distillation flask and added 3 L of double-distilled water. The EOs were obtained by steam distillation for 3 h using a Clevenger-type apparatus. After removing water traces with anhydrous sodium sulfate, the EOs were stored at 4°C in a clean amber glass bottle until used.
Separation and analysis of oil components
The overall analysis was performed by GC/MS with a gas chromatograph Shimadzu brand (GC 2010 Plus) coupled to a QP 2010 Ultra mass spectrometer. The separation of EO components was achieved by capillary column chromatography on 0.25-μm-thick flash silica RTX-5MS (30 × 0.25 mm) using helium as eluting gas, with a flow rate set of 1.2 mL min−1. Samples (1 μL) were injected in split mode (leakage ratio, 1:50). The device was connected to a computer system managing a mass spectrum library NIST 98 and driven by software to monitor chromatographic analyses. Identification of EO components was made through the comparison of their retention indices with those of standard compounds of the computerized database (NIST 98).
Antifungal activity test
The possible fungicidal inhibitory activity of EOs of S. terebinthifolius and S. molle was determined as follows: preparation of different concentrations of each EO, that is, 250, 500, and 1000 ppm in PDA and 0.2% Tween 20 in case of C. acutatum and 2000, 4000, and 8000 ppm in case of B. cinerea. Then, after that, 14 mL aliquots of each EO and PDA were poured into Petri dishes. After the complete dry off of the agar surface under laminar flow, 0.5 cm in diameter fungal discs of the single above-mentioned mitosporic fungi, cutoff from 96 h of fresh cultures, were singularly inoculated in the center of each Petri dish prepared as above. All plates were incubated at 24°C ± 2°C for 4 days under dark conditions. The diameter of fungal mycelial growth was measured in mm. Petri dishes containing only PDA and PDA + Tween 20 were also inoculated with fungal discs of the same anamorphic fungi and used as a control. Each treatment was carried out in triplicate. The fungitoxicity is expressed as a percentage of growth inhibition (PGI) and calculated according to the formula of Zygadlo et al.
23
(Equation 1), herein reported, in comparison with that of the synthetic fungicide azoxystrobin incorporated at 80 μL 100 mL−1 to PDA nutrient medium:
where GC is the average diameter of fungal colony grown on PDA alone (control), and GT is the average diameter of fungal colony grown on PDA containing each EO.
Determination of MIC (96-well microplate method)
MIC was considered as the fungicidal effect of each EO and is defined as the lowest concentration of each EO that definitely inhibits the fungal growth. MIC was determined on 96-well culture plates by a microdilution method using microorganism suspension at a density of 108 spore mL−1. Exactly 10 mL of liquid suspension was prepared from fresh PDA fungal culture of 10 days old that was incubated at 24°C ± 2°C. Spore formation was ascertained after 9 days of incubation under a light microscope. The fungal suspension was centrifuged at 1100 g for 5 min. Stock solution of each EO was prepared in PDB liquid nutrient medium at 2000, 1800, 1600, and 1400 ppm in the case of C. acutatum and 10000, 9000, 8000, 7000, 6000, and 5000 ppm in the case of B. cinerea. Each hole of the 96-well microplate was then filled up with 200 μL of each single prepared EO suspension and 100 μL of fungal suspension. After that, the 96-well microplate was incubated for 12 days at 24°C ± 2°C, and then, the absorbance was read at λ 630 nm using the ELISA Microplate Reader instrument. All samples were tested in triplicate. Azoxystrobin was used as the reference fungicide. Technical procedures of Lehtinen et al. 24 have been followed with some minor changes.
To verify the fungicidal effect, fungal reculturing from each well of the 96-well microplates has been performed on PDA. The fungistatic effect of each tested EO was determined by monitoring the lowest EO concentration that caused a significant reduction in fungal mycelial growth in comparison with the positive control.
Antibacterial activity test
In these tests, the disc diffusion method of Bhunia Johnson 25 has been used with some modifications as hereafter explained: all tested bacterial strains, that is, B. megaterium, C. michiganensis, X. campestris, B. mojavensis, P. savastanoi, and P. syringae pv. phaseolicola, were cultured on KB, except for E. coli, which was cultured on LB selective substrate. All bacterial cultures were incubated at 30°C ± 2°C for 48 h. Bacterial suspension of each bacterial culture was prepared in sterile distilled water at a concentration of 108 CFU mL−1 (OD = 0.2 nm). A mixture of 0.7% of soft agar and bacterial suspension (9:1, v/v) was prepared, and 4 mL of this suspension was poured into each 10 mL KB Petri dish 90 mm in diameter. Blank discs (6 mm; Oxoid) were placed over KB plate surfaces after complete solidification, and 20 μL aliquots from the EO suspension at the following concentrations, original (TQ), 940, 470, and 235 μg μL−1 and tetracycline at 1600 μg mL−1, were carefully applied over discs. The antibacterial activity of the tested EOs was evaluated by measuring the inhibition zone diameter in mm.
Statistical analysis
Results obtained from the current research were statistically processed and subjected to analysis of variance, followed by Tukey's B and Duncan's post hoc multiple comparison tests, with a probability of P < .05 using SPSS statistical software package version 13.0 (2004) to detect the significance between the different concentrations of studied EOs.
Results and Discussion
Composition of Schinus EOs
The separation and analysis of EO components of the ground fruits and leaves of the two species of Schinus investigated were performed by GC-MS, as described above. The identification and quantification of EO components, achieved using the NIST database, are reported in Table 1.
Identification of fungal isolates
The isolate from strawberry was identified as B. cinerea based on the morphological features. The analysis of DNA sequences obtained from the isolate from olive shows a high similarity (99%—E value 0.0) with those present in the GenBank for C. acutatum (accession numbers KM594093 and JN943492 confirming the morphological identification by microscopic technique).
Antifungal activity test
All studied EOs have significantly reduced all tested fungi, which exhibited a different sensitivity level to each tested EO. Results of these trials showed that the studied EOs were able to reduce the mycelial radial growth of C. acutatum already at low concentrations (Fig. 1A). Higher EO concentrations were needed to hold back the fungal growth of B. cinerea (Fig. 1B).

Antifungal activity of the studied essential oils (EOs) against Colletotrichum acutatum
The results of antifungal activity of S. terebinthifolius EOs are in agreement with those of Gundidza et al., 26 who found that these EOs exhibited antifungal activity against Aspergillus flavus, C. albicans, and A. niger.
The antifungal activity of studied EOs is possibly due to the contents of volatile constituents, especially monoterpene and sesquiterpene fractions. 12,27,28 For antifungal activity, among those terpenes, the tri-terpenemoronic acid is considered the most important. It was isolated, for the first time, from Schinus sp. 28 Actually, moronic acid has been obtained from other medicinal plants, such as Ozoroa mucronata and Rhus javanica, and Brazilian propolis. 28,29
The moronic acid exhibited various biological activities, such as antitumor, 30 anti-AIDS, 31 antidiabetic, 32 and antimicrobial. 12
The major components identified in EO of ripe fruits of S. molle were α-phellandrene, β-pinene, β-phellandrene, limonene, and cyclohexane methanol,4-ethyl-,alpha.,alpha.,4-trimethyl-3-(1-methylethenyl)-,[1R-(1-α,3.α.,4.beta,)]. 33 The antifungal activity of moronic acid is due to the fungal cell membrane damage, with an alteration of their permeability and an inhibition of their respiration. 26,34
Determination of MIC (96-well microplate method)
The effects of the studied EOs on the tested fungi resulted, in this respect, variable. In particular, EO of S. terebinthifolius leaves strongly inhibited both tested fungi, showing the lowest MIC values of 1400 and 6000 ppm against C. acutatum (Fig. 2A) and B. cinerea (Fig. 3A), respectively. EOs of S. terebinthifolius fruits and S. molle leaves showed slightly minor effectiveness, exhibiting MIC values of 1600 ppm against C. acutatum (Fig. 2B,C) and 5000 and 6000 ppm against B. cinerea (Fig. 3B,C), respectively.
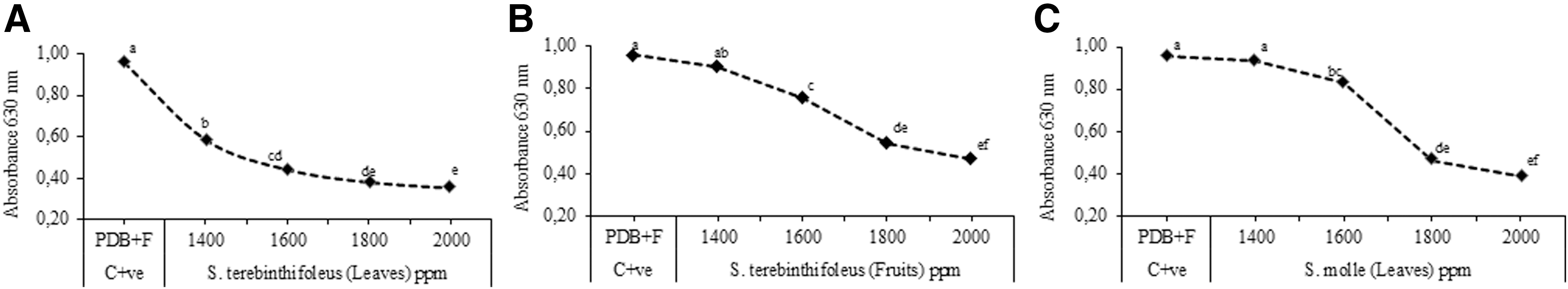
Minimum inhibitory concentration (MIC) of studied EOs (ppm) against C. acutatum (Microplate 96).

MIC of studied EOs (ppm) against B. cinerea (Microplate 96).
The moderate mycelial growth inhibitory activity of studied EOs against B. cinerea has recently also been reported by El-Badawy and Abdelgaleil, 35 who showed that the EO of S. molle has a lower effect on B. cinerea than other tested EOs extracted from Artemisia monosperma, Cupressus macrocarpa, and Pelargonium graveolens. Ibrahim and Al-Naser 33 showed an analogous level of inhibitory effect against the above anamorphic fungus for an n-hexane EO extracted from the same Schinus species.
Antibacterial activity test
The EO extracted from leaves of S. terebinthifolius has the most significant activity against the majority of tested bacterial species, except for P. savastanoi and P. syringae pv. phaseolicola (Table 2), whereas EOs isolated from S. terebinthifolius fruits and S. molle leaves showed a moderate significant antibacterial effect against B. mojavensis and a slightly significant activity against B. megaterium, C. michiganensis, and X. campestris compared to that of S. terebinthifolius leaves (Table 2). Generally, the tested G+ve bacteria were more susceptible to the tested EOs than G−ve ones.
Values are recorded as the mean diameter of inhibition zone (mm) ± standard deviation. Values followed by the different letters in each vertical column are significantly different according to Tukey's B test at P < .05. Data were obtained from three replicates.
TQ, original concentration; E. col, Escherichia coli; B. meg, Bacillus megaterium; C. mich, Clavibacter michiganensis; X. cam, Xanthomonas campestris; B. moj, Bacillus mojavensis; P. sav, Pseudomonas savastanoi, and P. syr, Pseudomonas syringae pv. phaseolicola.
The notable antibacterial activity of EO of S. terebinthifolius leaves has also been reported by Guerra et al. 36 and Gundidza et al. 26 against Yersinia enterocolitica, P. aeruginosa, E. coli, Acinetobacter alcoaceticus, B. subtilis, and Klebsiella pneumoniae.
According to the results of previous research, the antibacterial activities of studied EOs are not only due to their contents of monoterpene and sesquiterpene
28
but also due to the phenolic hydroxyl groups forming hydrogen bonds with the active site of target enzymes.
27
Therefore, the antimicrobial activity of the EO of S. terebinthifolius leaves could be ascribed to the presence of biologically active compounds, such as α-pinene, α-phellandrene,
The low activity observed against E. coli, P. savastanoi, and P. syringae pv. phaseolicola could be due to the presence of lipopolysaccharides in the outer membrane of the G−ve bacteria, which make them inherently resistant to external agents, such as hydrophilic dyes, antibiotics, and detergents. 37 –39
The obtained antibacterial activity results are in agreement with those of Cole et al., 39 who found that EO of S. terebinthifolius fruits was active particularly against tested G+ve bacteria, such as Corynebacterium sp., Bacillus sp., and Nocardia sp., while G−ve species, Enterobacter sp., E. agglomerans, E. coli, and K. oxytoca, showed less sensitivity to the tested EO.
Cole et al. 39 have reported that the differences of antibacterial activity among G−ve and G+ve bacteria are due to the structure of their cell walls. Whereas the G−ve bacteria have a more complex cell wall composed of a thin peptidoglycan layer and an outer membrane containing lipopolysaccharides, the G+ve ones are protected by a wall predominantly composed of one type of macromolecule (peptidoglycan).
Conclusion
The implementation of EO treatments especially for controlling some fungal and bacterial human or plant diseases is very promising nowadays compared to synthetic fungicides and antibiotics. Results from the current study showed that the three tested EOs can be potentially used for controlling the two studied phytopathogenic fungi C. acutatum and B. cinerea in a dose-dependent manner. In particular, all tested EOs were able to control C. acutatum at low concentrations ranging from 250 to 1000 ppm rather than B. cinerea, which has been controlled at higher concentrations ranging from 2000 to 8000 ppm.
In addition, the studied EOs were effective in controlling most of the investigated bacterial species: P. savastanoi and P. syringae pv. phaseolicola showing a complete resistance toward all of them. E. coli exhibited a moderate resistance only toward EO extracted from leaves of S. terebinthifolius and a complete resistance toward the other two tested EOs extracted from fruits of S. terebinthifolius and leaves of S. molle.
In conclusion, the current study validates, in an organized way, that most of the studied plant EOs possess substantial antimicrobial properties against several plant or human pathogenic microorganisms. This can explain their use in folk medicine and pharmaceutical industry for curing several serious diseases. Other trials are recommended in the near future to study the in vivo antimicrobial efficacy of crude EOs against several phyto- and human diseases. Moreover, the isolation and chemical identification of most of their bioactive substance(s) and the evaluation of its effectiveness compared to synthetic fungicides and/or antibiotics would also be advisable. Further studies are needed to determine which of the compounds characterizing our studied EOs are responsible for the observed antimicrobial activity, as well as to understand the exact mechanism of action of the compound(s).
Footnotes
Acknowledgments
We are grateful to Prof. Gian Luigi Rana for the critical revision of the article. We also appreciate the assistance of Prof. Michèle Parisien in revising the English editing. Thanks for the technical assistance from C.T. Michele Palumbo.
Author Disclosure Statement
No competing financial interests exist.