
Abstract
A Mucuna pruriens protein concentrate was hydrolyzed with a digestive (pepsin–pancreatin) enzymatic system. The soluble portion of the hydrolysate was fractionated by ultrafiltration and the ultrafiltered peptide fraction (PF) with lower molecular weight was purified by reversed-phase high-performance liquid chromatography. The PF obtained were evaluated by testing the biological activity in vitro. Fractions showed that the ability to inhibit the angiotensin-converting enzyme had IC50 values that ranged from 2.7 to 6.2 μg/mL. Trolox equivalent antioxidant capacity values ranged from 132.20 to 507.43 mM/mg. The inhibition of human platelet aggregation ranged from 1.59% to 11.11%, and the inhibition of cholesterol micellar solubility ranged from 0.24% to 0.47%. Hydrophobicity, size, and amino acid sequence could be factors in determining the biological activity of peptides contained in fractions. This is the first report that M. pruriens peptides act as antihypertensives, antioxidants, and inhibitors for human platelet aggregation and cholesterol micellar solubility in vitro.
Introduction
T
Bioactive peptides are specific protein fragments that have a positive impact on body functions and may ultimately influence health. These peptides can be produced by enzymatic hydrolysis using single or multiple specific or nonspecific proteases because the process requires a shorter time to obtain a similar degree of hydrolysis, as well as better control of the hydrolysis to obtain more consistent molecular-weight profiles and peptide composition. 3 Food proteins from both plant and animal sources have been used to obtain a wide range of peptides with biological activity. 4
Pharmacological inhibition of the renin–angiotensin system (RAS) is a major therapeutic strategy currently used to manage hypertension and reduce the risks of cardiovascular events. In the blood, renin, an aspartyl protease, cleaves liver-derived angiotensinogen to form the decapeptide angiotensin I (Ang I). The angiotensin-converting enzyme (ACE) hydrolyzes inactive Ang I into the vasoconstrictor octapeptide angiotensin II (Ang II). 5 ACE-inhibitory peptides block the first step in the RAS and interrupt the negative feedback effects of Ang II. 3
An excess of Reactive Oxygen Species, both due to excessive production or impaired antioxidant capacities, or both, is harmful and leads to what is known as oxidative stress. Several chemical methods with different mechanisms of action have been developed to measure the antioxidant potential of food proteins and peptides. This is because of the complexity of the oxidative reactions taking place in biological systems. 4
The suppression or inhibition of platelet aggregation is believed to decrease the incidence of the disease. 6 Currently, the primary antiplatelet agents available for clinical use act as metabolic inhibitors of platelet activation. However, these agents have significant adverse effects, particularly hypotension, which may limit their usefulness in patients with cardiovascular instability. 7 Food-derived peptides that inhibit blood platelet aggregation can be useful natural ingredients for the prevention of thrombosis. 8
Dyslipidemia, characterized by the presence of one or more abnormal serum lipid concentration, is a prime risk factor for cardiovascular disease. 3 Some bioactive peptides have hypocholesterolemic activity. It has been suggested that hypocholesterolemic peptides inhibit the micellar solubility of cholesterol, decreasing in vivo cholesterol absorption.
Chromatography is used for isolation and purification of bioactive peptides, and among the many chromatography techniques, high-resolution fractionation by size exclusion chromatography and high-performance liquid chromatography (HPLC) is particularly well-suited to function as the final step in a purification regimen. 9 For the abovementioned reason, the objective of this study was to evaluate the ACE inhibitory, antioxidant, antithrombotic, and hypocholesterolemic activity of peptide fractions (PF) purified by HPLC from Mucuna pruriens protein hydrolysates.
Materials and Methods
Seeds and chemicals
Pods of M. pruriens were collected in Yucatan, Mexico. After thoroughly drying, the pods were thrashed to remove seeds. The seeds, after thorough clearing and removal of broken seeds, foreign materials, and immature seeds, matured and dried seeds were stored in airtight plastic jars at room temperature (25°C). All chemicals were reagent grade or better and purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Protein concentrate
Selected grains were ground in a disk mill (model 4-E Quaker; Mill Straub Co., Philadelphia, PA, USA) and then sifted through 4.76- and 2.38-mm screens to remove the smallest particles before the air classification. Hulls were removed with a fluidizing air bed and the flour resulted was milled in a Cyclotec mill (Tecator, Höganas, Sweden) until passing through a 0.841-mm screen. The protein concentrate of M. pruriens bean was obtained by wet fractionation following Herrera-Chalé et al., 10 with some modifications. Briefly, 5.0 kg M. pruriens bean flour was suspended in 3% sodium bisulfite in a 1:6 (w:v) ratio, pH was adjusted to 8 using 1.0 M NaOH, and the suspension left to soak under constant agitation for 1 h. The suspension was then passed through a 0.177-mm screen to separate the fiber solids from the protein and starch containing liquid portion. Residual solids were washed thrice with 300 mL of 3% sodium bisulfite. The suspension left to sediment for 30 min to recover the starch after separation of the solubilized protein. The pH of the protein solution was adjusted to 4.2 with 1.0 M HCl. The suspension was centrifuged at 1317 g for 20 min (Mistral 3000i; Curtin Matheson Sci., Houston, TX, USA), the precipitate was freeze-dried at −47°C and 13 × 10−3 mbar (FreeZone 4.5; Labconco, Kansas City, MO, USA), pulverized, and stored until required.
Preparation of hydrolysate from M. pruriens protein concentrate
Hydrolysis was done under controlled conditions (temperature, pH, and stirring) in a 1000-mL reaction vessel equipped with a stirrer, thermometer, and pH electrode. The hydrolysis with the sequential pepsin–pancreatin system was done for 90 min: predigestion with pepsin for 45 min followed by incubation with pancreatin for 45 min. Hydrolysis parameters were substrate concentration 4%; enzyme/substrate ratio 1:10; pH 2.0 for pepsin; pH 7.5 for pancreatin; and 37°C hydrolysis temperature. 11 The reaction was stopped by heating to 80°C for 20 min, followed by centrifuging at 1317 g for 20 min (Mistral 3000i; Curtin Matheson Sci.) to remove the insoluble portion. Degree of hydrolysis was calculated by determining free amino groups with o-phthaldialdehyde 12 ; DH = h/htot ×100; where htot is the total number of peptide bonds per protein equivalent, and h is the number of hydrolyzed bonds.
Hydrolysate fractionation
The pepsin–pancreatin hydrolysate (PPH) was fractionated by ultrafiltration 13 with a high-performance ultrafiltration cell (Model 2000; Millipore, Darmstadt, Germany). The supernatant of PPH was collected. Part of it was studied as whole hydrolysate and the rest was fractionated with a high-performance ultrafiltration cell (Model 2000; Millipore). Five fractions were prepared using four molecular-weight cutoff (MWCO) membranes: 1, 3, 5, and 10 kDa. Soluble fractions were prepared by ultrafiltering the hydrolysates through the MWCO membranes beginning with the largest cartridge (10 kDa). The retentate and permeate were collected separately, and the retentate recirculated into the feed until maximum permeate yield was reached, as indicated by a decreased permeate flow rate. The permeate from the 10 kDa membrane was then filtered through the 5 kDa membrane with recirculation until maximum permeate yield was reached. The 5 kDa permeate was then processed with the 3 kDa membrane and the 3 kDa permeate with the 1 kDa membrane. The five ultrafiltered PF from PPH were prepared and designated as >10 kDa (10 kDa retentate); 5–10 kDa (10 kDa permeate–5 kDa retentate); 3–5 kDa (5 kDa permeate–3 kDa retentate); 1–3 kDa (3 kDa permeate–1 kDa retentate); and <1 kDa (1 kDa permeate). Peptide content in ultrafiltered PF was quantified using the method of Lowry et al. 14
Reversed-phase HPLC
Ultrafiltered PF designated as <1 kDa was dissolved in deionized water and injected in a preparative HPLC (Model 1110; Agilent, Santa Clara, California, USA) reverse phase column (C18 Hi-Pore RP-318, 250 × 10 mm column; BIO-RAD, Hercules, California, USA). 9 The injection volume was 100 μL, and the sample concentration was 20 mg/mL. Elution was achieved by a linear gradient of acetonitrile (ACN) in water (0–30% in 50 min) containing 0.1% trifluoroacetic acid at a flow rate of 4 mL/min at 30°C. Elution was monitored at 215 nm, and the fractions obtained were collected for assay of biological activity.
ACE inhibitory activity
Ang I-converting enzyme inhibitory activity in the hydrolysate and the ultrafiltered PF was analyzed following Hayakari and Kondo 15 Hippuryl-L-histidyl-L-leucine (HHL) is hydrolyzed by ACE to yield hippuric acid and histidyl-leucine. This method relies on the colorimetric reaction of hippuric acid with 2,4,6-trichloro-s-triazine (TT) in a 0.5 mL incubation mixture containing 40 μmol potassium phosphate buffer (pH 8.3), 300 μmol sodium chloride, 40 μmol 3% HHL in potassium phosphate buffer (pH 8.3), and 100 mU/mL ACE. The mixture was incubated at 37°C for 45 min and the reaction terminated by adding TT (3% v/v) in dioxane and 3 mL of 0.2 M potassium phosphate buffer (pH 8.3). After centrifuging the reaction mixture at 10,000 g for 10 min, the enzymatic activity was determined in the supernatant by measuring absorbance (A) at 382 nm. All runs were done in triplicate. The ACE inhibitory activity was quantified by a regression analysis of ACE inhibitory activity (%) versus peptide concentration and defined as an IC50 value, that is, the peptide concentration (μg protein/mL) required to produce 50% ACE inhibition under the described conditions. Determinations were made in triplicate.
Trolox equivalent antioxidant capacity
The ABTS•+ radical cation was produced by reacting 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid (ABTS) with potassium persulfate. 16 To prepare the stock solution, ABTS was dissolved at a 2 mM concentration in 50 mL phosphate-buffered saline (PBS) prepared from 4.0908 g NaCl, 0.1347 g KH2PO4, 0.7098 g Na2HPO4, and 0.0749 g KCl dissolved in 500 mL ultrapure water. If pH was lower than 7.4, it was adjusted with NaOH. A 70 mM K2S4O8 solution in ultrapure water was prepared. The ABTS radical cation was produced by reacting 10 mL ABTS stock solution with 40 μL K2S4O8 solution and allowing the mixture to stand in darkness at room temperature for 16–17 h before use. The radical was stable in this form for more than 2 days when stored in darkness at room temperature. Antioxidant compound content in hydrolysates and PF were analyzed by diluting the ABTS•+ solution with PBS to an A of 0.800 ± 0.030 AU at 734 nm. After adding 990 μL of diluted ABTS•+ solution (A 734 nm = 0.800 ± 0.030) to 10 μL of 6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (TROLOX) standard (final concentration 0.5–3.5 mM) in PBS, A was read at room temperature exactly 6 min after initial mixing. All analyses were run in triplicate. The percentage decrease in A at 734 nm was calculated and plotted as a function of the antioxidant concentration of Trolox for the standard reference data. To calculate the trolox equivalent antioxidant capacity (TEAC), the slope of the A inhibition percentage versus antioxidant concentration plot was divided by the slope of the Trolox plot. This produces the TEAC at a specific point in time. Determinations were made in triplicate.
In vitro inhibition of human platelet aggregation
The in vitro inhibition of human platelet aggregation was analyzed following protocols of Miyashita et al. 17 Blood from human volunteers, diluted with 130 mM trisodium citrate (4.5 mL), was centrifuged at 127 g for 15 min at room temperature in a Solbat J-600 centrifuge to produce platelet-rich plasma (PRP) as a supernatant. After removing most of PRP, the residual phase was centrifuged at 1918 g for 15 min at room temperature to obtain platelet-poor plasma (PPP). A platelet adjustment was then immediately performed to the PRP (200 × 103/μL) using a hematic cytometer (Sysmex KX-21). The PRP (450 μL) was incubated with each test compound, which was dissolved in saline (154 mM NaCl at pH 6.4) for 1 min at 37°C. Platelet aggregation was induced by adding 10 μL of an aqueous solution of 10 μm adenosine diphosphate. Aggregation was measured by an aggregometer (Chrono-Log 400) as the increase in the light transmission through PRP. The PPP, which was used as the control, exhibited 100% transmission. The platelet aggregation inhibition percentage was determined by subtracting the platelet aggregation percentage from the PRP with protein hydrolyzed from the platelet aggregation percentage from the PRP in basal conditions.
In vitro cholesterol micellar solubility inhibition
The in vitro micellar solubility of cholesterol was measured according to the methods described by Zhong et al. 18 with some modifications. The in vitro assay of micellar cholesterol solubility, which consists of a micellar solution (7.0 mL) containing 10.0 mM sodium taurocholate, 2.0 mM cholesterol, 5.0 mM oleic acid, 132.0 mM NaCl, 15.0 mM sodium phosphate (pH 7.4), and 1.0 mL of hydrolysate or PF, was prepared by sonication. The mixture was incubated at 37°C for 24 h and ultracentrifuged at 100,000 g for 60 min at 37°C. The supernatant was collected for the determination of total cholesterol content using a COBAS C 111 analyzer (Roche Diagnostics, Indianapolis, IN, USA).
Amino acid composition
Amino acid composition was determined by high-performance liquid chromatography. 19 Samples (4 mg of protein) were treated with 4 mL of HCl 6.0 N, placed in hydrolysis tubes, and gassed with nitrogen at 110°C for 24 h. They were then dried in a rotavapor (Rotavapor R-215; Büchi, Flawil, Switzerland) and suspended in a sodium borate buffer (1.0 M, pH 9.0). Derivatization was performed at 50°C using diethyl ethoxymethylenemalonate. Amino acids were separated using HPLC with a reversed-phase column (300 × 3.9 mm, Nova Pack C18, 4 mm; Waters, Milford, Massachusetts, USA) and a binary gradient system with sodium acetate containing 25 mM (A) 0.02 g/L sodium azide at pH 6.0, and (B) ACN as solvent. The flow rate was 0.9 mL/min and the elution gradient time was 0.0–3.0 min, linear gradient A:B (91:9) to A–B (86:14); time 3.0–13.0 min, elution with A–B (86–14); time 13.0–30.0 min, linear gradient A–B (86:14) to A–B (69:31); time 30.0–35.0 min, elution with A–B (69:31).
Statistical analysis
All results were analyzed using central tendency and dispersion measures. A least significant difference multiple range test was used to determine differences between treatments and biological activities in purified PF. All analyses were done according to Montgomery 20 and processed with the Statgraphics Plus version 5.1 software.
Results and Discussion
Preparation of hydrolysate from M. pruriens protein concentrate
Food-protein-derived bioactive peptides have been extensively investigated for potential application in the management and treatment of some human diseases. These peptide sequences are inactive with the intact protein structure but could exert several pharmacological properties upon release due to the activity of the proteases. The hydrolysate was evaluated for their degree of hydrolysis, as this measure is strongly correlated with protease activity. 21 Extensive hydrolysis of the M. pruriens protein concentrate with the sequential system, pepsin–pancreatin, produced a hydrolysis degree of 40.15%. In general, increasing the degree of hydrolysis increases the number of low-molecular-weight peptides and improves their bioactive properties. Thus, the structure and activity of the peptides released depend largely on the specificity of the proteolytic enzyme used in hydrolysis. 22 Pepsin and pancreatin are aspartic endoproteinases specific to hydrophobic amino acids, with preference for aromatic ones. 23 Digestive proteases have been used to hydrolyze different sources of proteins to obtain hypocholesterolemic, hypolipidemic, and antithrombotic peptides. 24
Hydrolysate fractionation
Ultrafiltration membrane-based separations of enzymatic hydrolysates can remove peptides from nonhydrolyzed proteins and proteolytic enzymes. In addition, ultrafiltration also can be used to perform peptide separation according to their molecular masses and charges. 25 The protein content was determined through filtrates and retentates. The highest protein content was observed in PF >10 kDa (1.813 mg/mL) and the lowest in PF <1 kDa (0.088 mg/mL). Although the hydrolytic activity of the PP enzymatic system was high (40.15%), this produced primarily high-molecular-weight peptides (>10 kDa) and a small amount of low-molecular-weight peptides. Usually low-molecular-weight peptides exhibited higher bioactivity. Thus, the ultrafiltered PF <1 kDa was selected to carry out the purification by reversed-phase high-performance liquid chromatography (RP-HPLC).
RP-HPLC chromatography
The principle of chromatographic fractionation is based on the interaction of the proteins or peptides, with the solid support (stationary phase) and the mobile phase. Fractionation is achieved by using mobile phase gradients, through which proteins or peptides are eluted differentially by changing the organic modifier concentration over time. 26 Ultrafiltered PF <1 kDa was further separated into several peaks by preparative RP-HPLC (Fig. 1).

Elution profile of the <1 kDa ultrafiltration fraction of the M. pruriens protein hydrolysate purified by reversed-phase high-performance liquid chromatography (RP-HPLC). Data are presented as mean of three replicates.
The qualitative analysis of the chromatograms indicated a greater affinity of the components of the ultrafiltered PF for the mobile phase through the column, recording a high elution rate of less than 30 min, eluting the first polar components and the nonpolar afterward. Five fractions (F1–F5) were thus collected throughout the elution time. The highest protein content was observed in F5 (1.495 mg/mL) and the lowest in F2 (0.721 mg/mL). These five fractions were evaluated by testing the biological activity in vitro.
ACE inhibitory activity
The sample concentration required to produce a 50% inhibition of ACE (IC50) was used as an activity indicator. This indicator is expressed as μg protein/mL, with smaller values indicating greater ACE inhibiting power. In a previous study, the ACE inhibitory activity of the PPH and their ultrafiltered fractions was evaluated. The inhibitory effect of the PF <1 kDa was 10.2 mM/mg. 10 The IC50 values of the RP-HPLC PF are shown in Figure 2. Fractions F3 and F4 exhibited the highest effects. Fractions F1 and F5 also showed notable ACE inhibitory effects, that is, IC50 values lower than 6.0 μg/mL, and fraction F2 came last in terms of effect, but still exhibited a reasonable IC50 value.
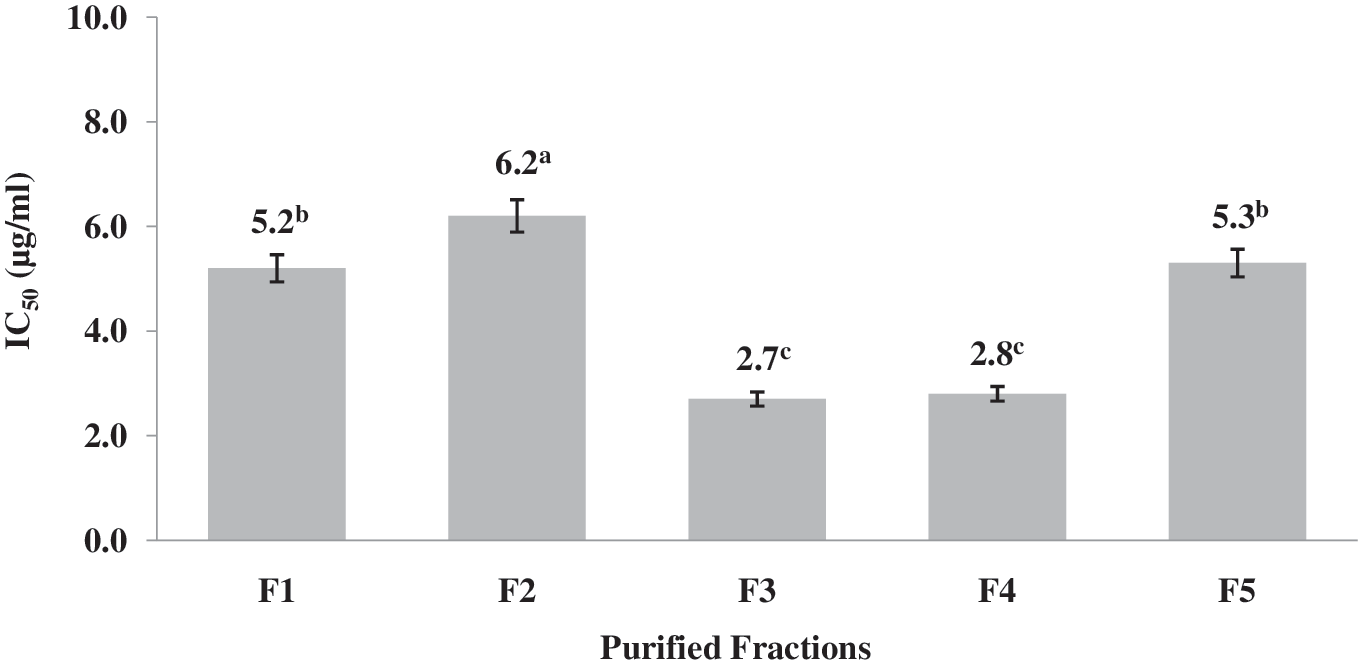
IC50 values of RP-HPLC peptide fractions. a–cDifferent letters indicate statistical difference (P < .05).
Tavares et al. 27 obtained six RP-HPLC fractions from whey protein hydrolysate that exhibited ACE inhibitory activity (IC50 values), ranging from 5.78 to 52.59 μg/mL. These authors identified a total of 14 peptides by HPLC-MS/MS in the six fractions. All peptides that were identified contained between 5 and 11 residues. In the present study, the IC50 values ranged from 2.7 to 6.2 μg/mL. This indicates that the fractions generated from the hydrolysate of M. pruriens have a higher ACE inhibitory activity and are likely to consist of low-molecular-weight peptides. Most of the reported peptides exhibiting ACE inhibitory activity are of a low molecular weight (approximately <12 amino acids). These types of peptides act as competitive inhibitors that can bind to the active site to block it or to the inhibitor-binding site that is remote from the active site so as to alter the enzyme conformation such that the substrate no longer binds to the active site. This causes a decrease in inhibition percentage as substrate concentration increases, very much like drugs such as captopril. This means that active peptides can be selected, to a degree, by ultrafiltration and RP-HPLC, resulting in the production of fractions with higher ACE inhibition than the original hydrolysate. 28 This is consistent with the behavior observed in the present study.
Trolox equivalent antioxidant capacity
The main characteristic of an antioxidant is its ability to trap free radicals. Highly reactive free radicals and oxygen species are present in biological systems from a wide variety of sources. These free radicals may oxidize nucleic acids, proteins, lipids, or DNA and can initiate degenerative disease. 29 In a previous study, the antioxidant activity of the PPH and their ultrafiltered fractions was evaluated. The inhibitory effect of the PF <1 kDa was 709.8 mM/mg. 10 The TEAC (free radical scavenging activity) values of the RP-HPLC fractions are shown in Figure 3. In terms of antioxidant activity, a significant difference was observed between the fractions (P > .05). TEAC values ranged from 132.2 (F1) to 507.43 mM/mg (F2).
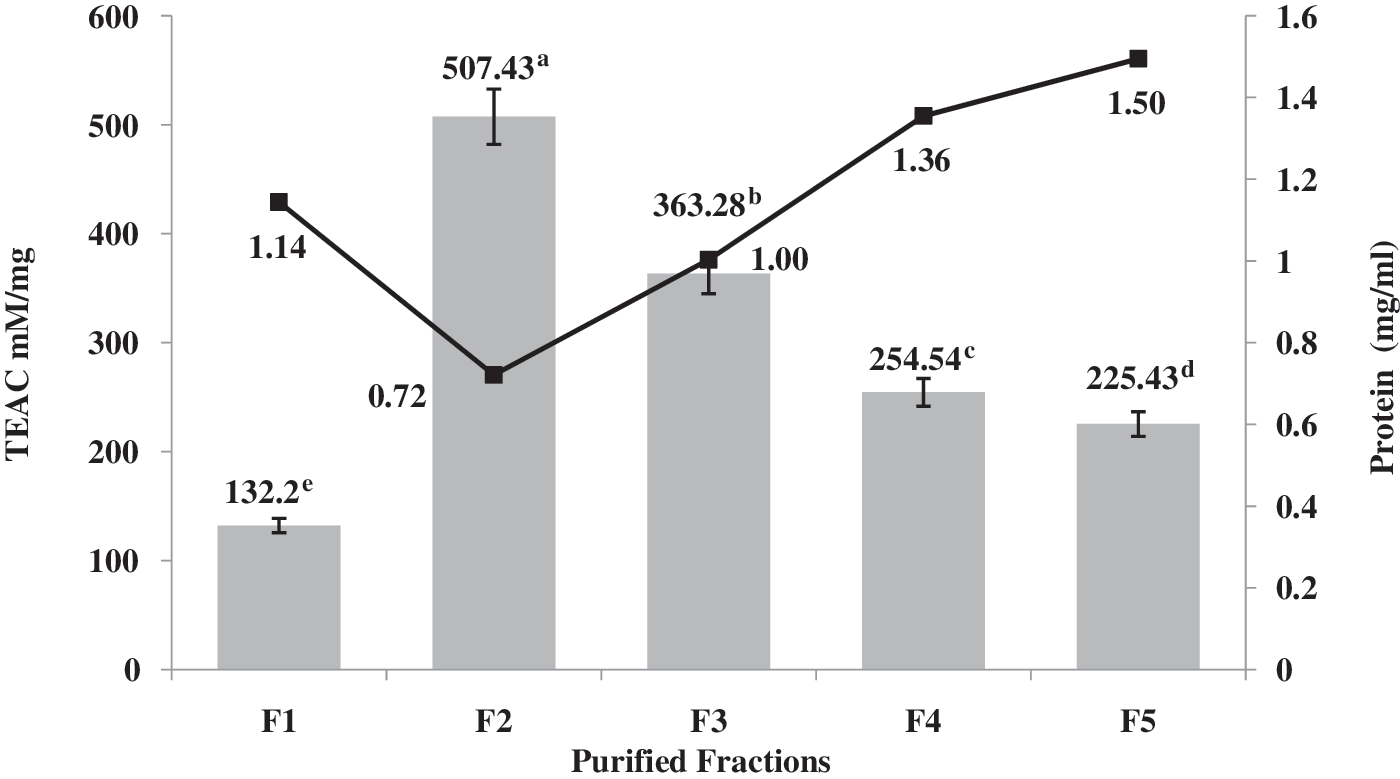
Trolox equivalent antioxidant capacity (TEAC) values of RP-HPLC peptide fractions. a–eDifferent letters indicate statistical difference (P < .05).
The results reveal that the scavenging activities of RP-HPLC fractions had no direct correlation with their hydrophobicity. According to Figure 1, fractions F3, F4, and F5 are more hydrophobic, but they had less scavenging activity than did F2. A similar conclusion was also made by Chen et al. 30 in the case of the His-containing peptides. The antioxidant activity of proteins in radical-mediated oxidation reactions may be due to their ability to act as a radical trapping device. The mode of action of the antioxidant peptides can be through metal binding activity, radical scavenging, active-oxygen quencher, and hydroxyl radical scavenger. The antioxidant capacity of amino acids such as tryptophan and tyrosine is due to its phenolic and indolic groups, which behave as hydrogen donors and thereby acted as antioxidants. However, all 20 amino acids found in proteins have the potential to interact with free radicals if the energy of the radical is high. Thus, the primary sequence of bioactive peptides plays an important role with regard to their antioxidant activity. Recently, it has been widely thought that hydrophobic amino acids act as effective antioxidants in a linoleic acid emulsion system by increasing the affinity to lipophilic fatty acids and thereby facilitating better interaction with free radicals. 31,32
Inhibition of human platelet aggregation
Atherothrombotic disease, such as coronary artery disease and stroke, is one of the major causes of death. Platelets play an important role in these arterial diseases. Suppression of platelet activity by materials may decrease incidence of the disease. In a previous study, the ability of the PPHs and their ultrafiltered fractions to inhibit platelet aggregation in PRP was evaluated. The inhibitory effect of the PF <1 kDa was 2.15%. After the RP-HPLC purification of PF <1 kDa, five fractions were obtained. The protein content of the fractions ranged from 0.721 to 1.495 mg/mL. Inhibition of human platelet aggregation ranged from 1.59% to 11.11%, with the highest inhibition in fraction F4 (Fig. 4). The percentages of inhibition are comparable to the moderate antithrombotic activities of synthetic antithrombotic compounds like aspirin and indomethacin.

Inhibition of human platelet aggregation of RP-HPLC fractions. a–eDifferent letters indicate statistical difference (P < .05).
Shimizu et al. 6 hydrolyzed pork meat using papain to obtain PF. The starting PF with mean molecular weight 2.5 kDa showed antithrombotic activity in vivo after oral administration to mice at 210 mg/kg body weight. The fraction with mean molecular weight 2.517 kDa, further purified by cation exchange chromatography, showed antithrombotic activity after oral administration at 70 mg/kg body weight. Antithrombotic activity of the purified PF was equivalent to that of aspirin at 50 mg/kg body weight. The purified fraction in the present study presented lower molecular weight than did those studied by Shimizu et al. 6 Studies have identified three distinct fibrinogen-derived peptides that are inhibitors of platelet aggregation. The amino acid sequences of these peptides form RGD, which recurs twice in the fibrinogen α chain; HHLGGAKQAGDV, the carboxyl terminus of the fibrinogen γ-chain; and GPRP. RGD-containing peptides and γ-chain-related peptides appear to inhibit platelet aggregation by binding to the platelet receptor complex GPIIb/IIIa and blocking adhesive glycoprotein attachment. GPRP binds directly to the fibrinogen D domain and inhibits the ability of fibrinogen to bind to GPI1b/IIIa. GPRP is also able to inhibit fibrin gel formation through binding to the D domain of fibrinogen. After RP-HPLC purification, the antithrombotic activity increased fivefold in F4, and the amount of protein required to exhibit the activity was 1.355 mg/mL. It is possible that this PF could be beneficial in preventing atherothrombosis.
In vitro cholesterol micellar solubility inhibition
Bile acid-binding agents contribute to the modulation of blood cholesterol levels in hypercholesterolemia. Bile acids are metabolites of cholesterol produced in the liver and reabsorbed in the intestine. Thus, inhibitors of bile acid absorption or bile acid sequestrants bind and prevent reabsorption of bile acids in the intestine. The resulting water-insoluble complex formed is subsequently removed through the feces, which leads to the upregulation of bile acid synthesis from cholesterol with concomitant decreases in hepatic and blood cholesterol levels. Inhibition of in vitro cholesterol micellar solubility of the RP-HPLC fractions is shown in Figure 5.

Cholesterol micellar solubility inhibition of RP-HPLC fractions. a–dDifferent letters indicate statistical difference (P < .05).
The in vitro cholesterol micellar solubility inhibition of the RP-HPLC fractions ranged from 0.24% (F2 and F3) to 0.47% (F5). The inhibitory activity micellar solubility of cholesterol is a strategy that has been used in the search for potent hypolipidemic and hypocholesterolemic food proteins and peptides. 18 Proteins and peptides could exhibit hypocholesterolemic and hypolipidemic in vivo effects by means of increased cholesterol catabolism, reduced cholesterol synthesis, or increased expression of the LDL reception genes. 22 Thus, the in vitro evaluation of the hypocholesterolemic activity could be correlated with the in vivo effect. Considering the above, the RP-HPLC PF obtained in this study could exhibit hypocholesterolemic effects in vivo.
Amino acid composition
The protein hydrolysates, ultrafiltered PF <1 kDa, and RP-HPLC fractions were subjected to amino acid composition analysis to determine the possible effects of the amino acids on the biological activities (Table 1). During hydrolysis, asparagine and glutamine were partially converted to aspartic acid and glutamic acid, respectively. The data for asparagine and/or aspartic acid were therefore reported as Asx, while those for glutamine and/or glutamic acid were reported as Glx. From the amino acid composition of hydrolysate, ultrafiltered PF, and RP-HPLC fractions, the hydrophobicity average was calculated using the hydrophobic values for each amino acid suggested by Tossavainen et al. 33
The data are expressed as means (n = 3).
Hydrophobicity values according to Tossavainen et al. 33
Aspartic acid + asparagine.
Glutamic acid + glutamine.
PF, peptide fractions.
In all cases, RP-HPLC PF showed higher average hydrophobicity than did the original hydrolysate and their ultrafiltered PF. These RP-HPLC PF had higher contents of certain hydrophobic amino acids in relation to the original protein hydrolysate. Thus, higher amounts of Val and Cys are observed in the RP-HPLC PF. The RP-HPLC fractions with the highest ACE inhibitory activities were F3 and F4. The higher antioxidant activity was observed in F2. In addition, F4 exhibited the greatest inhibition of human platelet aggregation and F5 had the highest inhibition of cholesterol micellar solubility. However, hydrophobicity is not the only factor in determining the biological activity of peptides. Other factors, such as the peptide size or amino acid sequence, may be critical for this inhibitory activity. In further studies, the sequence of the peptides present in fractions with higher biological activity will be determined. This will establish the relationship between the primary structure of the peptides and their mechanism of action.
In conclusion, we show in this work that M. pruriens proteins hydrolyzed with digestive proteases represent a source of peptides with biological activity. Ultrafiltration and RP-HPLC proved to be good strategies for obtaining PF with a greater biological activity than did the starting hydrolysate. PF could act as potential therapeutic agents due to their ability to inhibit the ACE, their capacity to act as radical scavengers, their ability to inhibit human platelet aggregation and the cholesterol micellar solubility. Reported results show that a combination of different characteristics, such as amino acid composition or hydrophobicity, may be responsible for the biological activity of the generated PF. Currently, further studies are being carried out to determine the sequence of the peptides as well as the in vivo biological activity of these M. pruriens-derived peptides.
Footnotes
Acknowledgment
This research forms part of Project 154307 “Investigación científica dirigida al desarrollo de derivados proteínicos de M. pruriens con potencial actividad biológica para la prevención y/o tratamiento de enfermedades crónicas asociadas al sobrepeso y la obesidad,” financed by the Consejo Nacional de Ciencia y Tecnología (CONACYT).
Author Disclosure Statement
No competing financial interests exist.