
Abstract
Capsicum baccatum is the most consumed red pepper species in Brazil. Our previous studies demonstrated the anti-inflammatory properties of its crude extract, whose activity is yet to be fully characterized. Herein, we examined the anti-inflammatory in vivo effects of enriched extracts obtained through bioguided fractionation as dichloromethane (DCM), butanol (BUT), and residual aqueous (RAq) extracts and its influence on inflammatory mediators produced by macrophages in vitro. We demonstrated that all C. baccatum extracts presented anti-inflammatory activity in vivo. In addition, we showed that BUT and RAq were more effective in inhibiting the neutrophil migration induced by carrageenan (Cg) to peritoneal cavity and both extracts inhibited paw edema induced by Cg, prostaglandin E2, and histamine in mice. Furthermore, the pretreatment with C. baccatum extracts significantly reduced the levels of myeloperoxidase (MPO) in the paw tissues of mice compared with the carrageenan group. Once again, RAq and BUT caused the greatest reduction in MPO levels. Moreover, it was demonstrated for the first time that C. baccatum inhibited the nitric oxide and tumor necrosis factor-alpha production by lipopolysaccharide/interferon gamma (IFN-γ)-stimulated macrophages. These anti-inflammatory effects seem to be at least, in part, independent of capsaicin. Hence, red pepper has bioactive compounds and might be used to develop food-derived extracts to treat related inflammatory diseases.
Introduction
T
Peppers from Capsicum species are used as a food ingredient worldwide and have been found to present hypocholesterolemic, anti-inflammatory, antioxidant, and antinociceptive effects. 10 –12 C. baccatum var. pendulum, a red pepper widely consumed in Brazil, also presents anti-inflammatory properties that need to be better investigated. In a previous study, our group demonstrated that extracts obtained from C. baccatum fruits present antioxidant and anti-inflammatory properties. It was demonstrated that the treatment of mice with ethanolic and butanol extracts of C. baccatum (200 mg/kg, p.o.) reduced the number of leukocytes and protein exudation to the lung of animals inoculated with carrageenan (Cg). 13 In the present study, to enlarge the understanding to the anti-inflammatory activity of C. baccatum, the effects of the crude ethanol extract together with three enriched (dichloromethane, butanol, and residual aqueous) extracts from fruits of C. baccatum were investigated through carrageenan-induced peritonitis model and paw edema formation induced by carrageenan, histamine, and prostaglandin E2 (PGE2) in mice. In addition, the influence of C. baccatum on inflammatory mediators produced by macrophages was also evaluated.
Materials and Methods
Plant material
Fruits of Capsicum baccatum var. pendulum (Willd.) Eshbaugh (Solanaceae) were obtained from a cultivated area in Turuçu, Rio Grande do Sul, Brazil. A voucher specimen (number P278) was identified and deposited at the Herbarium of the Brazilian Government Research Institute EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária).
Preparation of extracts and chemical composition
The plant material used in the present study was the same prepared and characterized by Zimmer et al. (2012) 13 and they were maintained at −80°C until use to obtain the results presented herein. The sample chemical analysis had been repeated over this period and it did not detect significant differences. The fruit without seeds was dried in a circulating air oven (40°C) for seven days and then triturated to powder. To obtain the ethanolic extract, 1 kg of fruits was extracted with 70% ethanol (plant:solvent, 1:10, w/v) under reflux for 4 h. The fruit of C. baccatum (5 kg) was also submitted to successive extractions in a Soxhlet apparatus using dichloromethane, n-butanol, and water until complete exhaustion. The solvents were then evaporated under reduced pressure to dryness and, the aqueous extract was lyophilized, to obtain the respective residues from fruits named crude ethanol (CE) extract, dichloromethane (DCM) extract, butanol (BUT) extract, and residual aqueous (RAq) extract. All extracts were maintained at −80°C until use. The content of total phenolic compounds in the extracts was estimated using the Folin–Ciocalteu method described by Singleton et al. (1999), 14 while the total content of flavonoids was analyzed using the aluminum chloride colorimetric method described by Chang et al. (2002). 15 The contents of the total phenolic (expressed as mg GAE/g extract) and total flavonoid (expressed as mg QE/g extract) were 180.08 ± 3.76 and 34.36 ± 4.04 for CE, 149.28 ± 2.46 and 102.48 ± 6.38 for DCM, 187.51 ± 2.34 and 54.68 ± 2.92 for BUT, and 186.00 ± 6.82 and 20.80 ± 1.56 for RAq, respectively. In addition, the presence of capsaicin and the flavonoids, quercetin and rutin, in the extracts was analyzed by HPLC. Quercetin and rutin were not identified in C. baccatum extracts, and only CE and DCM extracts contained capsaicin.
Animals
Male BALB/c mice (6–8 weeks old) were obtained from the State Foundation for Health Science Research, Porto Alegre, Brazil and housed under standard conditions (22°C, 12-h light/ 12-h dark cycle) with food and water ad libitum in groups of 6 per cage (41 × 34 × 16 cm) in the animal facility of the Universidade Federal de Ciências da Saúde de Porto Alegre (UFCSPA) for at least one week before experiments. All of the experimental procedures were performed in accordance with the guidelines of the National Institute of Health and the Brazilian Society for Science on Animals of Laboratory with the approval of local ethics committee (CEUA UFCSPA number 105/13).
Carrageenan-induced peritonitis in mice
The groups of BALB/c mice (n = 6) were pretreated s.c. with phosphate-buffered saline (PBS) (control group 0.1 mL/10 g) or with C. baccatum extracts (CE, BUT, DCM, and RAq at doses of 50, 100, and 300 mg/kg/0.2 mL) dissolved in PBS plus Tween 80 (0.5%) 30 min before carrageenan injection (i.p., 500 μL/cavity/0.2 mL). Dexamethasone (v.o., 2 mg/kg) was used as the positive control drug and was administered 1 h before carrageenan injection. After 4 h, the mice were euthanized under ketamine/xylazine and the peritoneal cavity cells were harvested by washing the cavity with 3 mL of PBS containing EDTA 1 mM. The recovered volumes were similar in all animals of each experimental group. The total number of leukocytes was determined using a neubauer chamber, and differential cell counts were carried out on cytocentrifuge slides stained with panotic. The results are presented as number of neutrophils per cavity.
Paw edema induced by inflammatory stimuli
Groups of BALB/c mice (n = 6) were pretreated s.c. with PBS (0.1 mL/10 g, control group) or with extracts (100 mg/kg/0.2 mL) dissolved in PBS plus Tween 80 (0.5%) 30 min before edema induction. Dexamethasone (v.o., 2 mg/kg, positive control group) was administered 1 h before edema induction. Then, the animals received a 50 μL intraplantar injection of carrageenan solution (500 μg/paw), histamine (100 μg/paw), or prostaglandin E2 (5 μg/paw) into the right hind paw. The left paw received 50 μL of PBS. Paw edema was determined using a plethysmometer at 1–4 h after carrageenan injection and at 15, 30, 60, and 90 min after prostaglandin E2 or histamine injection. The difference between the right and left paw volumes was determined. Shortly after, the animals that received carrageenan into the paw were euthanized as described before, and the subcutaneous tissue of the right hind paw was removed and stored at −80°C for posterior myeloperoxidase (MPO) analysis.
MPO activity in carrageenan-induced paw edema
The MPO assay was carried out according to the literature. 16,17 Tissue samples of the right carrageenan-induced hind paw edema were homogenized at 5% (w/v) in EDTA/NaCl buffer (pH 4.7) and centrifuged at 644 g for 15 min at 4°C. The pellet was resuspended in 0.5% hexadecyltrimethyl ammonium bromide buffer (pH 5.4), and the samples were frozen and thawed thrice in liquid nitrogen. Upon thawing, the samples were recentrifuged (16,099 g, 15 min, 4°C), and 5 μL of the supernatant was used for the MPO assay. The enzymatic reaction was assessed with 1.6 mM tetramethylbenzidine, 80 mM of sodium phosphate buffer, and 0.3 mM hydrogen peroxide. The reaction was stopped by the addition of 4 M H2SO4 and absorbance was measured at 450 nm. To estimate the number of accumulated neutrophils into the paw of animals, a standard curve of neutrophils (0.3 × 104–10 × 104 cell/well) was prepared through serial dilutions of a cell suspension obtained from the peritoneal cavity of mice injected with carrageenan. Then, the number of neutrophils accumulated into paw of each mouse was estimated by interpolation from values of MPO activity obtained from a standard curve of neutrophils. 18
Macrophage viability assay
The macrophage viability was measured using the MTT assay. 19 Peritoneal macrophages were harvested from BALB/c mice that had been injected 3 days previously with a sterile thioglycolate solution (i.p., 2 mL, 3% w/v in PBS). The cells were plated at 2 × 105 cells/0.2 mL in DMEM plus 10% FBS, 100 U/mL penicillin, and streptomycin (100 μg/mL) in 96-well microtiter plates and were incubated overnight at 37°C in an atmosphere of 5% CO2. Then, the plates were washed before being cultured for 48 h at 37°C in DMEM in the presence or absence of extracts (3–300 μg/mL) previously dissolved in DMEM and filtered. Subsequently, MTT (5 mg/mL in PBS) was added (20 μL/well) and the incubations continued for 4 h. The purple formazan product was dissolved by the addition of acidic isopropanol, and the absorbance at 570 nm was measured using a SpectraMax M2 (Molecular Devices).
Measurement of nitric oxide production and the TNF-α content by macrophages
Peritoneal macrophages and the macrophage line RAW 264.7 (5 × 105 cells/well in 24-well plates) were preincubated for 24 h with 30–300 μg/mL of C. baccatum extracts previously dissolved in DMEM and filtered. The medium was discarded and the cells were stimulated with lipopolysaccharide [LPS, 10 ng/mL and interferon gamma (IFN-γ) (1 ng/mL)] in DMEM for an additional 24 h. At the end of this period, nitric oxide (NO) production was determined in the cell culture supernatants according to a colorimetric assay based on the Griess reaction. 19 Moreover, the TNF-α content was determined in the supernatants by ELISA Kit (eBioscience) in accordance with manufacturer's instructions.
Statistical analysis
Statistical analysis was performed by one-way ANOVA or repeated measure ANOVA followed by Bonferroni's test using GraphPad Software and SPSS Statistics. Values of P < .05 were considered significant.
Results
All the extracts of C. baccatum were able to inhibit the neutrophil migration elicited by carrageenan (Fig. 1). When the animals were treated with the extracts at doses of 100 and 300 mg/kg, the inhibition rates were of 27.9% and 42.5% (CE); 35.2% and 39.5% (DCM); 57.5% and 52.9% (BUT), and 60.1% and 75.4% (RAq), respectively. As expected, dexamethasone inhibited the neutrophil migration induced by Cg (Fig. 1A–D).
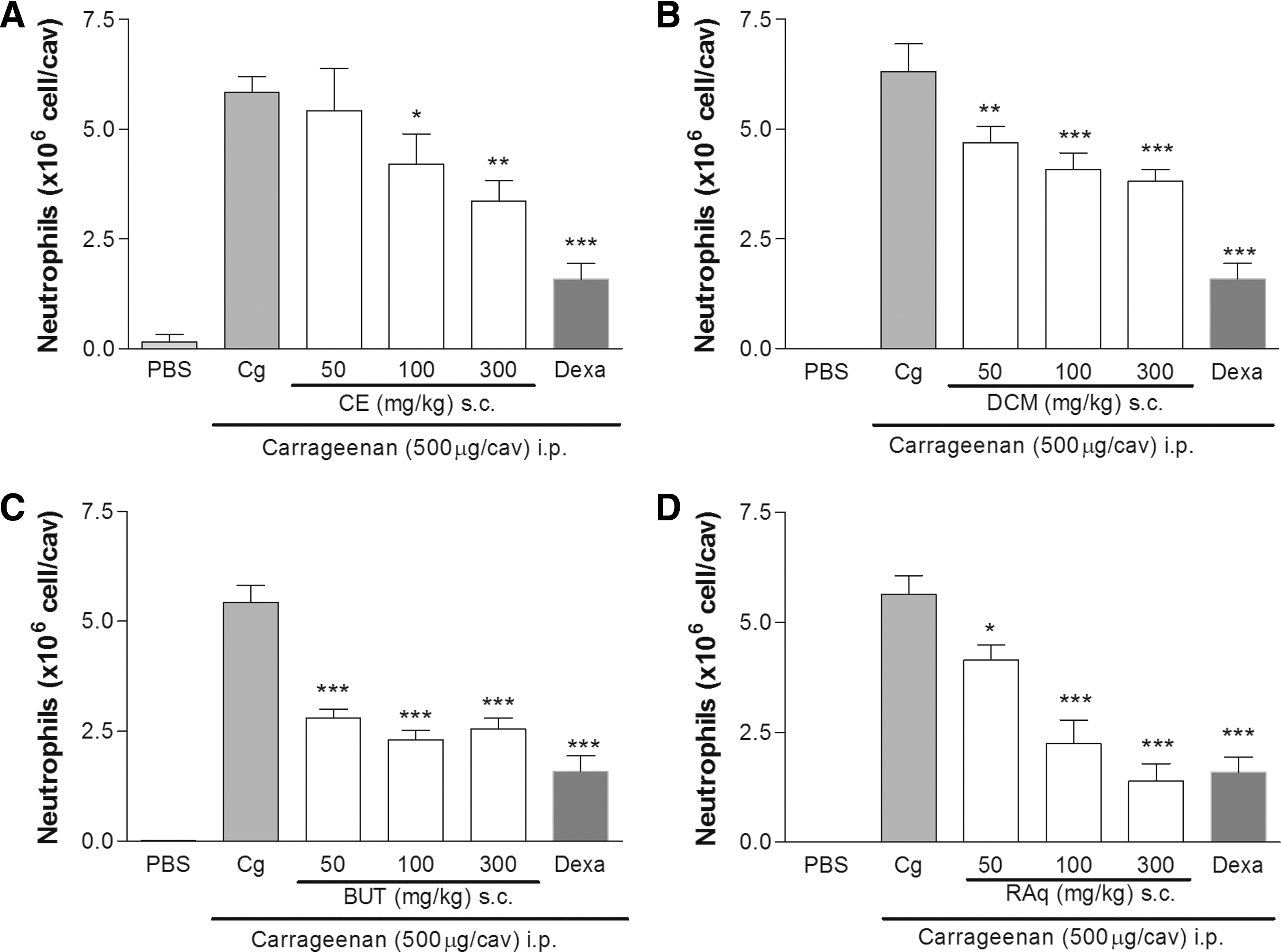
Effects of Capsicum baccatum extracts
C. baccatum extracts at 100 mg/kg induced a significant antiedematogenic response in paw edema elicited by carrageenan, histamine, and PGE2 in mice. The peak of edematogenic response was 3 h for Cg (Fig. 2A), 15 min for PGE2 (Fig. 2B), and 30 min for histamine (Fig. 2C). For effects of extracts on paw edema induced by Cg, repeated ANOVA measures revealed significant main effects for time × group (P = .007). The Bonferroni's post hoc revealed that at 1 h the inhibitory activity of RAq was significantly higher than those presented by DCM, BUT, and DEXA treatments. At 3 h, RAq also presented better activity than CE and DCM extracts. However, at 4 h, all extracts caused significant reduction in edema formation in relation to Cg group (Fig. 2A). Moreover, at the peak of response to Cg, one-way ANOVA revealed that dexamethasone and all extracts caused significant reduction in edema formation (CE 30% P < .05; DCM 34% P < .01; BUT 50% P < .001; RAq 63% P < .001; and dexamethasone 77% inhibition P < .001). In contrast, the area under curve analysis indicated that compared to CE (35.2%), DCM (35.7%), and BUT (39.2%), RAq presented higher inhibitory activity (RAq = 59.2% inhibition) (Fig. 2A filled area).

Effects of C. baccatum extracts on paw edema induced by carrageenan
Regarding the effect of C. baccatum extracts on paw edema induced by PGE2, no significant main effect for time × group was observed (repeated ANOVA measures P = .114) (Fig. 2B). Moreover, at the peak of edematogenic response, the one-way ANOVA revealed that DCM, BUT, and RAq extracts were able to acutely inhibit PGE2-induced paw edema, resulting in 23% (P < .01), 38% (P < .01), and 68% (P < .001) inhibition, respectively. However, the area under curve indicated that only BUT and RAq presented significant inhibitory activity (BUT 53% inhibition and RAq 73.5% inhibition) (Fig. 2B filled area).
For effects of extracts on paw edema induced by histamine, repeated ANOVA measures revealed significant main effects for time × group (P = .001). The Bonferroni's post hoc revealed that the treatments with BUT and RAq caused significant inhibition on histamine-induced paw edema at 30 and 60 min, respectively (Fig. 2C). In addition, at the peak of edematogenic response, the one-way ANOVA showed that BUT and RAq extracts caused significant inhibition on paw edema induced by histamine (BUT 46% inhibition, P < .01 and RAq 56% inhibition, P < .01). The area under curve indicated that BUT and RAq presented similar inhibitory activities causing 36.8% and 34% of inhibition, respectively (Fig. 2C filled area).
All of the extracts of C. baccatum were effective at 100 mg/kg for inhibiting the increase in Cg-induced MPO activity in the paws of mice (Fig. 3). The magnitude of inhibition was 39%, 42%, 83%, and 66% to CE, DCM, BUT, and RAq extracts, respectively, compared to the carrageenan group.
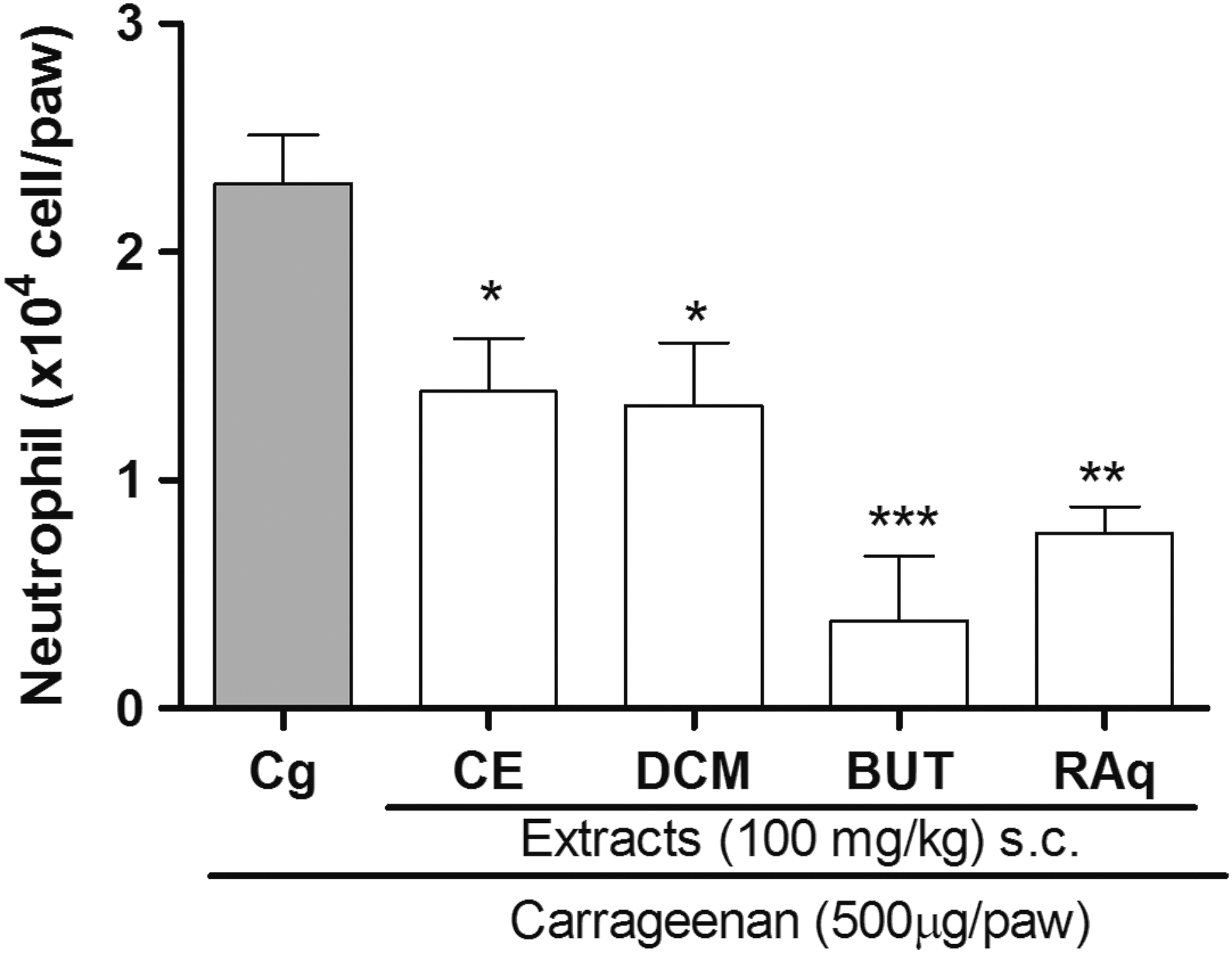
Effects of C. baccatum extracts on neutrophil infiltration to the paw tissues induced by carrageenan. Data are expressed as mean + SEM of four replicates and are representative of three experiments. ***P < .001, **P < .01, *P < .05 (one-way ANOVA followed by Bonferroni's test). Cg, carrageenan; CE, crude ethanolic extract; DCM, dichloromethane extract; BUT, butanol extract; RAq, residual aqueous extract.
None of the tested concentrations (30–300 μg/mL over 48 h) presented macrophage cytotoxicity in vitro (data not shown). The NO production induced by LPS/IFN-γ was significantly inhibited by all tested concentrations of four tested C. baccatum extracts compared to untreated cells (Fig. 4). The stimulation of peritoneal macrophages in vitro with LPS (10 ng/mL) plus IFN- γ (1 ng/mL) caused a significant increase in TNF-α production. The pretreatment of macrophages with CE, DCM, and BUT at 300 μg/mL for 24 h significantly decreased the TNF-α production induced by immune stimuli by ∼40% compared to untreated cells (Fig. 5).
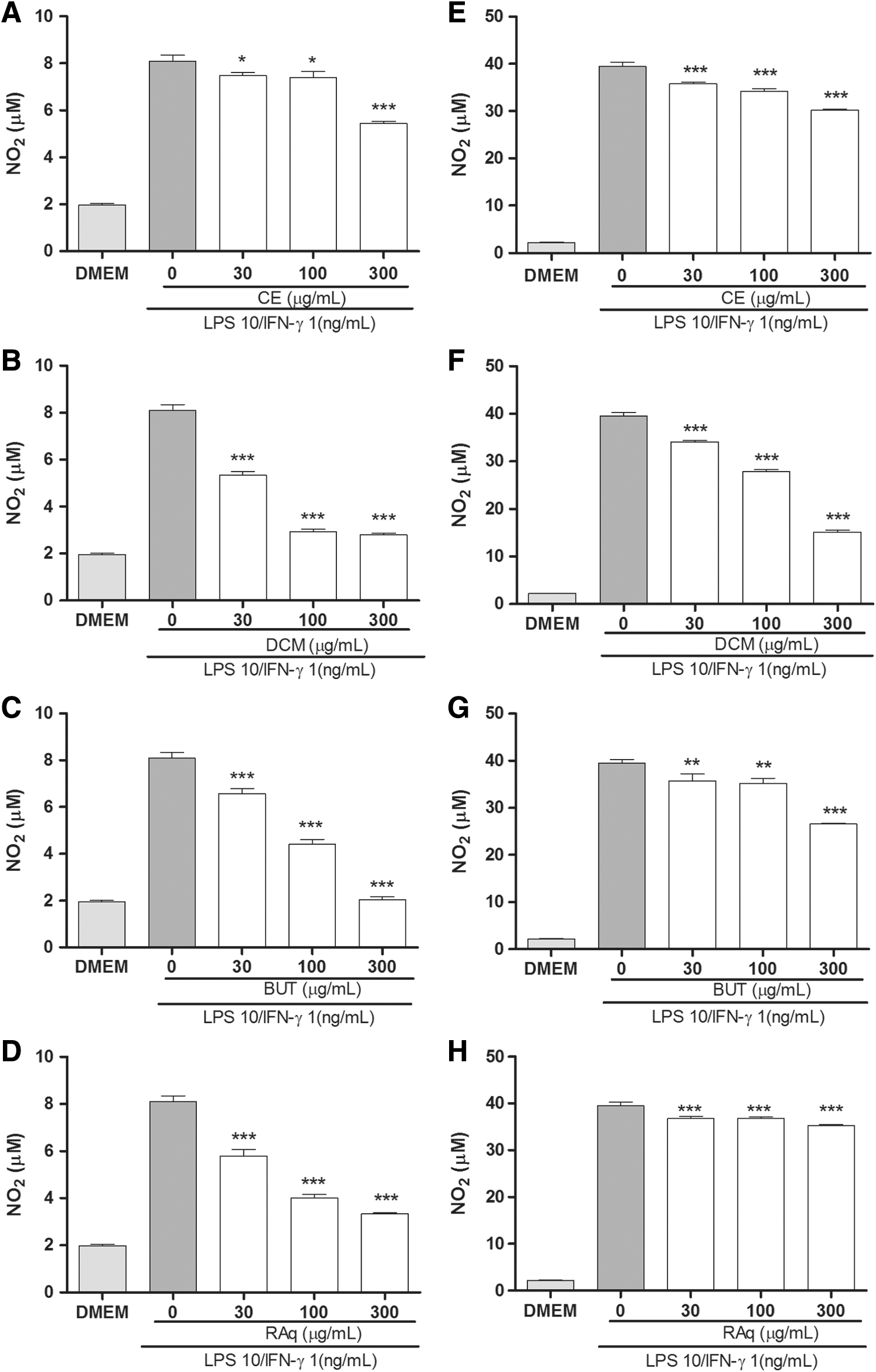
Effects of C. baccatum extracts on the production of nitric oxide by peritoneal

Effects of C. baccatum extracts on TNF-α production by peritoneal macrophages stimulated by LPS 10/IFN-γ 1 (ng/mL). Data are expressed as mean ± SEM of four replicates. *P < .05. (one-way ANOVA followed by Bonferroni's test). DMEM, culture medium; C, control group; CE, crude ethanolic extract; DCM, dichloromethane extract; BUT, butanol extract; RAq, residual aqueous extract.
Discussion
Acute inflammation is characterized by increased vascular permeability, resulting in the extravasation of fluids and proteins and the infiltration of leukocytes to the site of challenge or injury. Generally, neutrophils are one of the first inflammatory cells to migrate and accumulate at the site of injury, followed by mononuclear cells, including macrophages, and later by T lymphocytes. 3 These events are essential to the induction of the protective immune response during vaccinations and infectious challenges. However, in some cases, it can be dangerous to the host, for example, in exacerbated or prolonged responses to pathogens, in response to environmental and toxic compounds, as well as during autoimmune or chronic inflammatory disturbances. 20
It has been shown that many plant-derived compounds exhibit significant anti-inflammatory activities, including effects on the expression of pro-inflammatory molecules that are key to inflammation, such as inducible NO synthase, cyclooxygenase-2, TNF- α, among others. 9,21–22 In this context, we demonstrated herein that all C. baccatum extracts, CE, BUT, DCM, and RAq, inhibited the neutrophil migration induced by Cg to peritoneal cavity and also the paw edema induced by Cg, PGE2, and histamine in mice. Moreover, C. baccatum inhibited the production of two important inflammatory mediators, NO and TNF-α in vitro.
In the literature, there are few studies showing the anti-inflammatory properties of C. baccatum. Spiller et al. 23 demonstrated the anti-inflammatory effect of C. baccatum using a juice at very high dose (2.0 g/kg) and suggested that this effect may be related to capsaicin. Herein, we showed that the crude extract (CE) of C. baccatum at 100 mg/kg and the enriched extracts at 50 mg/kg (DCM, BUT, and RAq), administered subcutaneously in BALB/c mice, significantly decreased the number of neutrophils recruited to the peritoneal cavity in response to carrageenan, confirming our previous data showing that CE and BUT extracts administrated by gavage at 200 mg/kg inhibited total leukocyte infiltration into the lungs of mice inoculated with carrageenan. 13
Vasodilation and increased vascular permeability are events of the inflammatory process that precedes the cellular infiltration. To assess the effects of C. baccatum extracts on the edema, we evaluated the effects of extracts in the paw edema model induced by carrageenan, which induces a production of several inflammatory mediators, including histamine, PGE2, NO, and cytokines. 24 –26 Considering that all C. baccatum extracts were able to reduce carrageenan-induced neutrophil migration to peritoneal cavity at 100 mg/kg, this dose was used to carry out other investigations. As observed in neutrophil migration, the pretreatment of mice with all tested C. baccatum extracts caused significant reduction in paw edema formation in response to carrageenan, especially RAq. Furthermore, we also demonstrated for the first time that BUT and RAq extracts caused significant inhibition in paw edema induced by PGE2 and histamine at 100 mg/kg. Comparing the effects of extracts on paw edema, as well as in peritonitis model, the results presented in this study suggest an increased potency of the extracts with CE <DCM <BUT <Raq. In fact, BUT and RAq were more effective in inhibiting the paw edema induced by Cg, PGE2, and histamine compared to other fractions (EC and DCM), suggesting a better profile for inhibition of nonspecific inflammatory events.
As commented before, edema formation is accompanied by neutrophil infiltration to the inflammatory site. Thus, neutrophil infiltration into the paw tissue of mice injected with carrageenan was indirectly evaluated through the MPO activity assay. MPO secreted by neutrophils can induce damage to adjacent tissue contributing to the pathogenesis of inflammation. It is widely used as a marker of neutrophil infiltration. 16,27,28 Our results showed that pretreatment with C. baccatum extracts significantly reduced the levels of MPO in the paw tissues of mice compared with the carrageenan group, with RAq and BUT causing the greatest reduction in MPO levels. In summary, these data suggest that the antiedematogenic effect promoted by C. baccatum can be, at least in part, due to the inhibition of histamine, PGE2, and neutrophil migration. Some authors have suggested that capsaicin might be the major compound responsible for the anti-inflammatory effects of Capsicum species. 23,29 CE, DCM, BUT, and RAq extracts used in this study exhibited significant amount of phenolic and flavonoids, compounds that are not quercetin and rutin. In contrast, the presence of capsaicin was found only in CE and DCM extracts, 13 which presented the lowest anti-inflammatory effects. It is important to note that the amount of flavonoids and phenolic compounds present in the extracts was higher than those found in other Capsicum species. 13
Considering that BUT and RAq, which did not present detectable capsaicin, 13 were more effective in the inhibition of neutrophil infiltration and paw edema induced by different stimuli in vivo, we suggested that the anti-inflammatory effects of BUT and RAq are not due to capsaicin or quercetin and rutin. It is possible that other phenolic compounds can be involved in this effect. 22,30
During acute inflammation, resident cells, such as macrophages, sense the antigens and release inflammatory mediators that initiate the inflammatory process. Thus, the effects of C. baccatum extracts on macrophages were investigated. We demonstrated, for the first time, the effects of C. baccatum extracts on the production of TNF-α and NO by macrophages (peritoneal and RAW 264.7) stimulated by LPS/IFN-γ, a well-known in vitro model for studying new candidates for anti-inflammatory drugs. LPS, a component of the outer membrane of Gram-negative bacteria, is one of the most potent microbial initiators of inflammation and is responsible for activating monocytes and macrophages to produce cytokines, such as TNF- α, IL-1, and IL-6. 31
LPS associated with cytokines, such as IFN- γ, induces iNOS (inducible NO synthase) expression resulting in the production of abundant amounts of NO, which present many biological properties. 32 At physiological concentrations, it is responsible for maintaining a low vascular tone and inhibits leukocyte adhesion and transmigration, pro-inflammatory platelet aggregation, and oxidative injury in addition to antiproliferative, antioxidant, microbicidal, antiviral, antiparasitic, and antitumoral effects. 33,34 However, during the inflammatory process, large amounts of NO, produced mainly by neutrophils and macrophages, can be deleterious for the host. Recent evidence indicates that most of the cytotoxicity attributed to NO is actually due to peroxynitrite, which is produced from the reaction between NO and the free radical superoxide anion. 35 In vivo, peroxynitrite causes DNA damage and LDL (low density lipoprotein) oxidation, often associated with pathophysiological conditions, such as asthma, arthritis, multiple sclerosis, colitis, psoriasis, neurodegenerative diseases, tumor development, transplant rejection, and septic shock. 36,37 Thus, research on the design of new drugs for the treatment of NO-mediated diseases has focused on controlling the overproduction of NO through iNOS inhibition. 32 Mueller et al. 38 demonstrated that a crude extract of Capsicum annuum at 0.5 mg/mL reduced TNF-α and IL-6 secretion and iNOS expression in RAW 264.7 stimulated by LPS. Capsaicin, a compound of pepper, was also tested and positively influenced the profile of the secreted cytokines and reduced iNOS expression. We demonstrated that all of the extracts of C. baccatum at 30 μg/mL inhibited NO production induced by LPS plus IFN- γ, including BUT and RAq, which showed no detectable amount of capsaicin. 13
Our results revealed that the inhibitory effect of extracts on NO production was more evident in peritoneal macrophages compared to RAW 264.7 cells stimulated with LPS/IFN. RAW 264.7 macrophages are transformed macrophages that could behave differently compared to mice macrophages. In contrast, compared to peritoneal macrophages, RAW cells produced elevated levels (five-fold increase) of NO when stimulated by LPS/IFN-γ, which may hinder the effect of extracts in reducing these mediators. These results are in accordance with Huang et al. (2011). 39
Several pro-inflammatory cytokines are known to be involved in the pathogenesis of inflammation in addition to NO. Data from literature demonstrated the relationship between iNOS induction and its dependence on pro-inflammatory cytokines, such as TNF-α signaling. 40 TNF-α is one of the main cytokines involved in neutrophil recruitment during acute inflammation and is considered to be a key target in several inflammation-related disorders. 41,42 Herein, it was demonstrated that CE, DCM, and BUT extracts were able to inhibit the TNF-α production induced by LPS/IFN-γ in peritoneal macrophages.
As phenolic compounds have anti-inflammatory activity in vivo and in vitro, 22 they could be involved in the anti-inflammatory effects of C. baccatum extracts. However, we cannot rule out the possibility of degradation of some compounds during in vitro culture, with subsequent generation of ROS, which is known to induce transcriptional activity associated with the anti-inflammatory response. 43
The present study demonstrated that C. baccatum presented anti-inflammatory activities, being able to inhibit the production of NO and TNF-α in vitro, and decrease the neutrophil recruitment elicited by carrageenan and paw edema induced by carrageenan, histamine, PGE2 in vivo. These effects do not seem to have correlation to capsaicin. Further investigations might determine which compounds are involved in the activity to develop red pepper–derived extracts to treat inflammation.
Footnotes
Acknowledgments
The authors are grateful to the Brazilian agencies Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Cientifico e Tecnológico (CNPq), and Fundação de Amparo a Pesquisa do Estado do Rio Grande do Sul (FAPERGS) for financial support. GG and PRTR also thank CNPq for fellowships.
Author Disclosure Statement
No competing financial interests exist.