
Abstract
Silymarin (Si) is a herbal product with hepatoprotective potential, well-known for its antioxidant, anti-inflammatory, and immunomodulatory properties. We have recently demonstrated that the usual therapeutic doses of Si are capable of inhibiting the progression of incipient liver fibrosis. We aimed at further investigating the benefits of Si administration upon liver alterations after the hepatotoxin discontinuation, using CCl4 to induce liver injuries on rats. CCl4 administration induces first of all oxidative stress, but other mechanisms, such as inflammation and liver fibrosis are also triggered. Fifty Wistar rats were randomly divided into five groups (n = 10). The control group received sunflower oil twice a week for 8 weeks. Carboxymethyl cellulose group received sunflower oil twice a week, for 8 weeks and CMC daily, for the next 2 weeks. CCl4 group received CCl4 in sunflower oil, by gavage, twice a week, for 8 weeks. CCl4 + Si 50 group received CCl4 twice a week, for 8 weeks, and then 50 mg/body weight (b.w.) Silymarin for the next 2 weeks. CCl4 + Si 200 group was similar to the previous group, but with Si 200 mg/b.w. Ten weeks after the experiment had begun, we assessed inflammation (IL-6, MAPK, NF-κB, pNF-κB), fibrosis (hyaluronic acid), TGF-β1, MMP-9, markers of hepatic stellate cell activation (α-SMA expression), and proliferative capacity (proliferating cell nuclear antigen). Our data showed that Silymarin administered after the toxic liver injury is capable of reducing inflammation and liver fibrosis. The benefits were more important for the higher dose than for the usual therapeutic dose.
Introduction
N
More important, natural products from plants, with no or minimal side effects, which could influence the progression of liver fibrosis, are of crucial interest.
Silymarin (Si) is an extract obtained from the seeds of Silybum marianum and it is derived from the milk thistle plant. It is a unique flavonoid complex containing silybin, silydianin, and silychristin. Silymarin is a source of polyphenolic compounds with antioxidant, anti-inflammatory, and immunomodulatory properties 1 and it is used as an adjuvant in liver disorders for at least two reasons: it has cytoprotective properties and it has no side effects, as proven on volunteers. 2
We have recently demonstrated 3 that the usual therapeutic dose of Si can inhibit the progression of fibrosis in the early stages of liver alterations, in an experimental model of CCl4-induced liver fibrosis.
In the present study, the aim is to step forward and evaluate the capacity of the same natural product to accelerate the spontaneous reversion of liver fibrosis, after the hepatotoxin discontinuation, using also CCl4 to induce liver fibrosis on rats.
In drawing up the study, we have taken into consideration personal data and data from literature, which are already documented. First of all, it seems that the alterations induced by CCl4 administration become irreversible when administration is continued for more than 60 days. 4 Indeed, Muriel et al. have proven that the hepatoprotective properties of Silymarin are irrelevant to reverse established cirrhosis obtained after 3 months of CCl4 administration, whereas the discontinuation of CCl4 after 2 months of administration is followed by a spontaneous reversion of fibrosis. 5 So, in the present study, CCl4 has been administered for 2 months. In the meantime, in our previous studies, 6 using the same protocol as in this study, we induced moderate liver fibrosis through oral CCl4 administration for 2 months and we proved the validity of this model through histological examination.
Second, most of the compounds with hepatoprotective potential have been studied by simultaneous administration of the drug and the CCl4. However, many of these compounds may interfere with the cytochrome P450, and their proven efficiency could be the result of the inhibition of CCl4 action and, because of that, the absence of the fibrogenetic events. 7 So, in our study, Silymarin has been administered after the discontinuation of CCl4 administration.
Third, CCl4 administration is an oxidative stress model to induce fibrosis. When testing compounds with antioxidative properties for their antifibrotic potential, it might seem that the antioxidant capacity is no longer important when the product is administered once CCl4 is discontinued. 5 However, when studying the dynamics of oxidative stress parameters, mainly protein oxidation, after the discontinuation of CCl4 administration for 10 weeks, their augmentation was still emphasized for 5–10 weeks, and more important, active fibrogenesis was persistent. 4 So, antioxidant compounds could be helpful to reduce the progression of fibrosis after the hepatotoxin discontinuation, even when the demonstration is realized on a model of oxidative stress-induced liver fibrosis.
How could Silymarin improve the reversion of moderate fibrosis after toxin discontinuation, what fibrogenic mechanisms could be triggered, and in what dose this natural compound could have a better effect? These are the questions we have aimed at answering in this study.
Our interest has been focused upon the influence of Sylimarin on liver inflammation and fibrosis, after the hepatotoxin was withdrawn, and only indirectly upon oxidative stress evolution, through nuclear factor-kappa B (NF-κB) assessment. We have also taken into account the relation between the fibrosis reversion and the activity of metalloproteinases, since one of the factors considered to be responsible for the loss of fibrosis reversibility is the gradual loss of expression of metalloproteinases. 8 We have also considered the possible influence of Silymarin upon the proliferative capacity of hepatocytes, knowing the carcinogenic potential of chronic hepatopathies evolution.
The results of this study show that Silymarin is effective in reducing liver injuries produced by CCl4 administration, including liver fibrosis, in an experimental model that resembles the human pathology.
Materials and Methods
Experimental protocol
In the experiment, 50 male Wistar rats, with the weight of 200 ± 10 g were used. The animals were housed in the animal facility of the Physiology Department within “Iuliu Hatieganu” University of Medicine Cluj-Napoca, at a temperature of 24°C and with a 12-h light–12-h dark cycle. They had free access to water and standard pellet diet. All experiments were performed according to the approved animal care protocols of the Ethics Committee on Animal Welfare and in accordance with the directives of the Romanian Ministry of Health.
The rats were randomly divided into five groups of 10 animals each. The control group received 1 mL/kg body weight (b.w.) sunflower oil twice a week, by orogastric administration, for 8 weeks and then no substance was added for the next 2 weeks. The vehicle group (carboxymethyl cellulose [CMC]) received sunflower oil, as the control group, for 8 weeks, and then, for another 2 weeks, this group received 0.7 mL CMC 0.5% solution daily, by gavage. The CCl4 group (CCl4) received CCl4 solution (25% in sunflower oil) 1.2 mL/kg b.w by gavage, twice a week for 8 weeks, with the toxin discontinuation for another 2 weeks. The last two groups were treated with Silymarin. In the first 8 weeks of the experiment they received the same treatment as CCl4 group, but then the hepatotoxic administration was stopped and Silymarin extract suspended in 0.7 mL CMC 0.5% solution as vehicle was added for the next 2 weeks. Si was administered by gavage, daily, in a dose of 50 mg Silymarin/kg b.w. (group CCl4 + Si50), corresponding to the standard therapeutic dose 9 or at a dose of 200 mg Silymarin/kg b.w. (group CCl4 + Si200). Ten weeks after the beginning of the experiment, all the animals were sacrificed and blood and liver were collected.
To prepare the biological samples, blood was centrifuged at 1000 g for 5 min to separate the plasma. The left lateral lobe and right medial lobe of the liver were immersed in 10% formalin solution and prepared for histological analysis. The rest of the liver was washed with cold saline and homogenized with a POLYTRON homogenizer (Brinkmann KINEMATICA, Switzerland) in 50 mM TRIS—10 mM EDTA buffer (pH 7.4). The homogenates were centrifuged (4000 rotation/min for 10 min) and the supernatants were collected. From each animal, supernatants were stored in Eppendorf tubes at −80°C until assayed.
Quantification of interleukin-6 and mitogen-activated protein kinase
Interleukin-6 (IL-6), as a proinflammatory cytokine and key mediator in inflammation, was assessed in the liver of the experimental rats using the commercial ELISA Kits (R&D Systems, Minneapolis, MN), according to the manufacturer's instructions. The detection limit for IL-6 was 5 pg/mL. The results were expressed in pg/mg protein.
Mitogen-activated protein kinase (MAPK) (ERK1/2) was assayed using the ELISA Kit (Cell Signaling Technology, Inc., Danvers, MA), according to the manufacturer's instruction. MAPK is an intracellular pathway activated by oxidative stress.
Hyaluronic acid assay was performed according to the manufacturer's instructions, using a Hyaluronan DuoSet DY 3614 ELISA Kit (R&D Systems). It is synthesized by the hepatic stellate cell (HSC) in liver, and in excess, it is a marker of extracellular matrix (ECM) deposition.
Immunohistochemical analysis
The immunohistochemical study was performed using a Leica Bond-Max ™ Immunohistochemistry system. Quantification of proliferating cell nuclear antigen (PCNA)-positive hepatocytes was realized by counting 10 fields of 40× nonoverlapping fields for each slide and the data expressed as percentage of positive cells. Also, the immunohistochemical identification of alpha smooth muscle actin (α-SMA)-positive cells was performed. The anti-SMA antibodies were used for myofibroblast identification.
Western blot assay
Liver tissue samples were lysed using a buffer containing IGEPAL-Nonidet 1%, protease inhibitor complex 1% in phosphate-buffered saline for 1 h, on ice. Cell extracts were spun at 14,000 g for 30 min, at 4°C. The protein was quantified with the use of Bio-Rad protein assay reagents. The lysates were subjected to precast 12% SDS-PAGE gels under reducing conditions. Prestained molecular markers were used to estimate the molecular weight of the samples. Proteins were transferred to polyvinylidenedifluoride membranes under standard conditions. After nonspecific sites were blocked, blots were incubated with antibodies against phosphorylated nuclear factor-kappa B (pNF-κB), NF-κB, matrix metalloproteinase 9 (MMP-9), GAPDH (Santa Cruz Biotechnology, Santa Cruz, CA), transforming growth factor-beta 1 (TGF-β1), and α-SMA (Abcam plc., Cambridge, United Kingdom) diluted 1:200, overnight, at 4°C. After they had been washed, blots were incubated with secondary anti-mouse antibody and finally, chemiluminescence staining was performed by using SuperSignal West Femto Chemiluminescent substrate (Thermo Fisher Scientific, Rockford, IL) according to the manufacturer's instructions. Western blots were quantified (optical density) by Quantity One analysis software (Bio-Rad) and GAPDH was used as protein loading control.
Results
Effects of Silymarin on liver inflammation
IL-6 (Fig. 1A) was significantly increased in liver homogenates (17.68 ± 7.43 pg/mgP) in CCl4 group as compared with the control group (4.28 ± 1.69 pg/mgP) and CMC group (9.14 ± 2.7 pg/mgP). The administration of 200 mg/b.w. Silymarin significantly decreased the IL-6 level (6.2 ± 2.24 pg/mgP) as compared with the CCl4 group. There was no other statistical significance between the experimental groups.
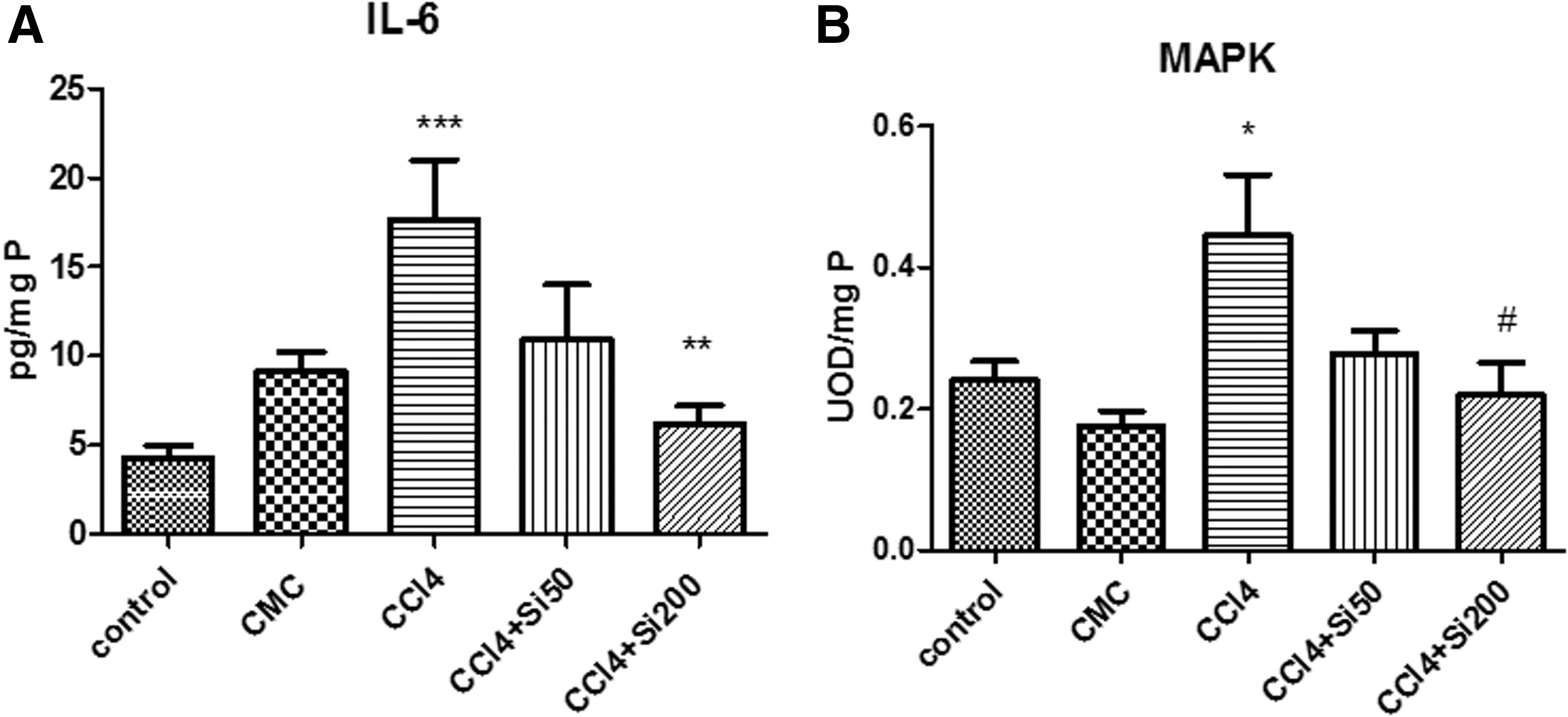
Effects of Silymarin administration upon the IL-6 level and MAPK activation in the liver homogenates. CCl4 was administered in sunflower oil, by gavage, twice a week for 8 weeks, in CCl4 group with the discontinuation of the hepatotoxin for the next 2 weeks. The groups with Silymarin administration received CCl4 by gavage, twice a week, for 8 weeks and for the next 2 weeks Silymarin alone was administered, in a dose of 50 mg/b.w. (CCl4 + Si 50 group) or 200 mg/b.w. (CCl4 + Si 200 group), respectively. The control group received sunflower oil for the first 8 weeks, and then no substance was added for the next 2 weeks. The vehicle group (CMC) received sunflower oil for the first 8 weeks, and CMC for the next 2 weeks.
Analysis of liver homogenates for changes in MAPK activation (Fig. 1B) revealed that the levels significantly increased in CCl4 group (0.44 ± 0.18 UOD/mgP) as compared with the control group (0.24 ± 0.06 UOD/mgP) and CMC group (0.17 ± 0.05 UOD/mgP). The administration of 200 mg/b.w. Silymarin significantly decreased (0.21 ± 0.10 UOD/mgP) the level of MAPK, with no other significant differences between the groups used in the experiment.
The synthesis of NF-κB (Fig. 2A–C), an important factor involved in transcription, was induced by the administration of CCl4 for 8 weeks and toxic discontinuation for another 2 weeks (we registered a 2.18-fold increase of pNF-κB, the phosphorylated, active form of this transcription factor, in the CCl4 group compared with the control group). When the rats were treated with Silymarin, we obtained a significant decrease in the pNF-κB levels (with a P < .01 for the CCl4 + Si50 group compared with the CCl4 group, and a P < .001 for the CCl4 + Si200 group compared with the CCl4 group), which was more important for the group that received a dose of 200 mg Silymarin (this dose was capable of lowering the level of pNF-κB down to 0.73× compared with the control group). Although the administration of CCl4 led to a significant increase in the synthesis of pNF-κB, we did not find a significant increase in the inactive form of this transcription factor. The 200 mg dose of Silymarin had the same effect on NF-κB (Fig. 2B) and it decreased its level down to 0.74× compared with the control group.

Effects of Silymarin on liver fibrosis
Liver fibrosis was assessed using the quantification of hyaluronic acid through ELISA technique and α-SMA, TGF-β1, and MMP-9 quantification through western blot.
The fibrosis score from the histological examination (graphs not shown) was 0 for control and CMC groups, 2.6 ± 0.54 for CCl4 group, 1 for CCl4 + Si50 group, and 0.2 ± 0.04 for CCl4 + Si 200 group. The administration of Silymarin, in both doses, significantly decreased the fibrosis score as compared with CCl4 effect alone, the effect being significantly more important for the 200 mg/b.w. Silymarin administration.
α-SMA quantification
The administration of CCl4 led to a significant increase in the expression of α-SMA, 3.83× the value for the control group (P < .001). The treatment with the two doses of Silymarin (Fig. 3F, G) was found to significantly decrease this marker of fibroblast activation, with only 2.9× the value of the control group for the low dose of Silymarin (P < .001 compared with the CCl4 group) and a 3.2× increase for the 200 mg Silymarin dose (P < .1 compared with the CCl4 group).

Effects of Silymarin administration upon liver fibrosis resolution. Rats treated with CCl4 for 8 weeks and with no other substance for another 2 weeks (CCl4 group), with CCl4 for 8 weeks and 50 mg/b.w. Silymarin for the next 2 weeks (CCl4 + Si 50 group), with CCl4 for 8 weeks and 200 mg/b.w. Silymarin for the next 2 weeks (CCl4 + Si 200 group) or only with vehicles (control and CMC groups).
TGF-β1 evaluation
The TGF-β1 (Fig. 3F, H) was significantly increased by the administration of CCl4 (P < .001 vs. the control group), but the treatment with Silymarin in both doses managed to significantly decrease the expression of this factor compared with the group that only received the profibrotic agent (CCl4 with a P < .001 for both doses).
MMP-9 quantification
CCl4 administration (Fig. 3F, 3I) was able to lower the level of MMP-9 (the level of MMP-9 was 0.84 times lower in this group compared with the control group, P < .001). Only the 200 mg dose of Silymarin increased the level of MMP-9 to 1.24-fold the value of the control group (P < .001 compared with the control group).
Hyaluronic acid quantification
The hyaluronic acid in liver homogenates (Fig. 4) was significantly increased by the CCl4 administration as compared with the control (1.75 × ) and CMC (1.83 × ) groups. Silymarin 50 mg/b.w. significantly decreased (1.4 × ) the hyaluronic acid content of the liver as compared with the CCl4 group, the levels remaining significantly higher when compared with the control and CMC groups. In the meantime, the 200 mg/b.w. Silymarin administration significantly decreased the hyaluronic acid in the liver to values comparable to those of the control and vehicle groups. There was no statistical difference between the CCl4 + Si 50 group and the CCl4 + Si 200 group.
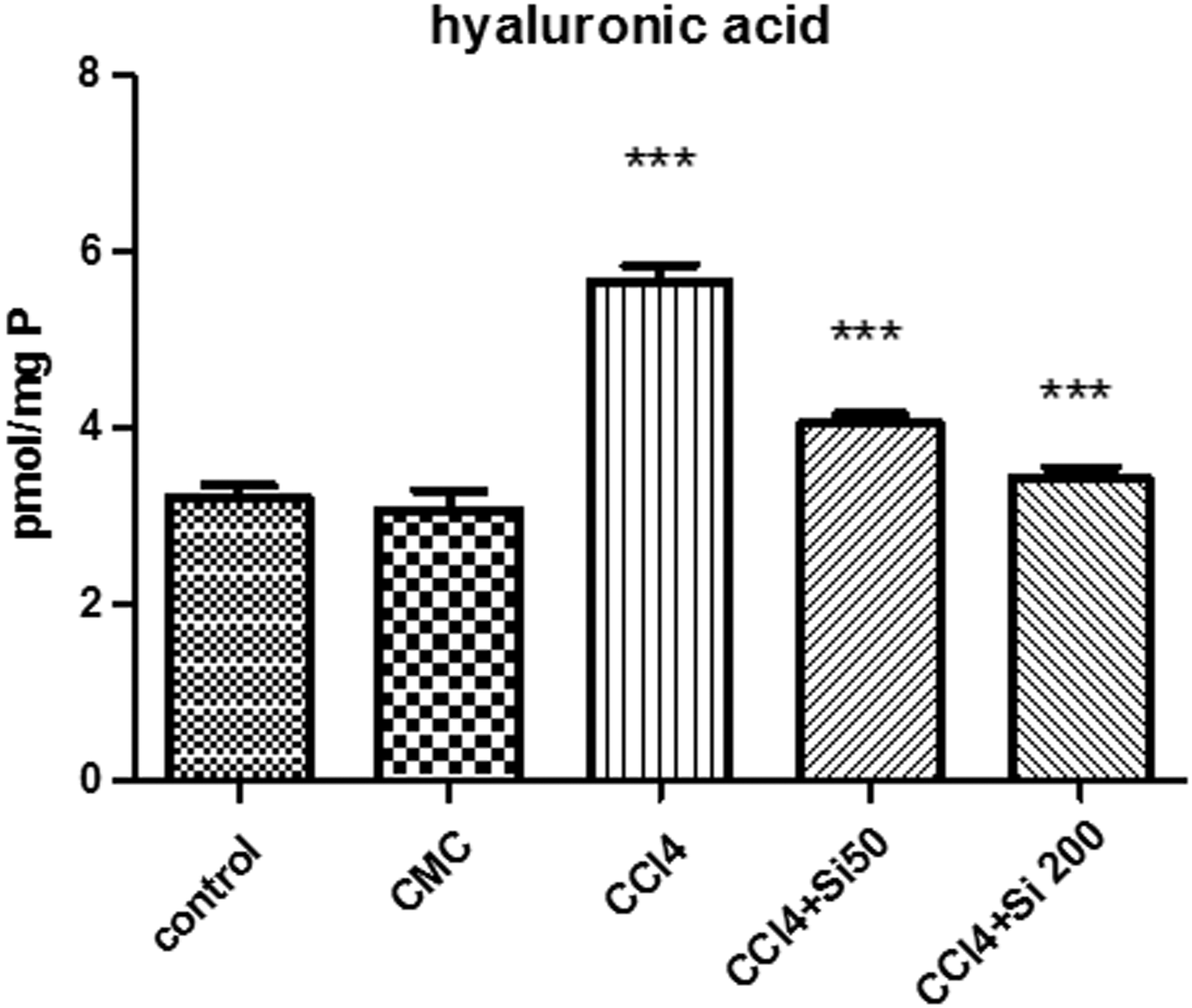
Effects of Silymarin administration upon the hyaluronic acid content in liver homogenates. The CCl4 administration for 8 weeks significantly increased the level of hyaluronic acid in the liver (***P < .001) as compared with control and CMC groups. Both doses of Silymarin (50 or 200 mg/b.w.), administered after 8 weeks of CCl4 administration, significantly decreased the hyaluronic acid level as compared with the CCl4 group (***P < .001), without a significant difference between the two results.
Effects of Silymarin on cellular proliferation
PCNA was used as a marker of hepatocyte proliferation (Fig. 5). CCl4 administration significantly increased the number of PCNA-positive cells (46.83% ± 6.66%) as compared with the control group (35.16% ± 4.88%) and CMC group (33.68% ± 2.97%). Silymarin administration, in both doses, significantly reduced the number of PCNA-positive hepatocytes in the experimental groups as compared with the CCl4 group. The values for CCl4 + Si 50 mg/b.w. group (31.82% ± 6.26%) and for CCl4 + Si 200 mg/b.w. group (28.37% ± 8.49%) were similar to those of the control and CMC groups without any significant difference between these groups.
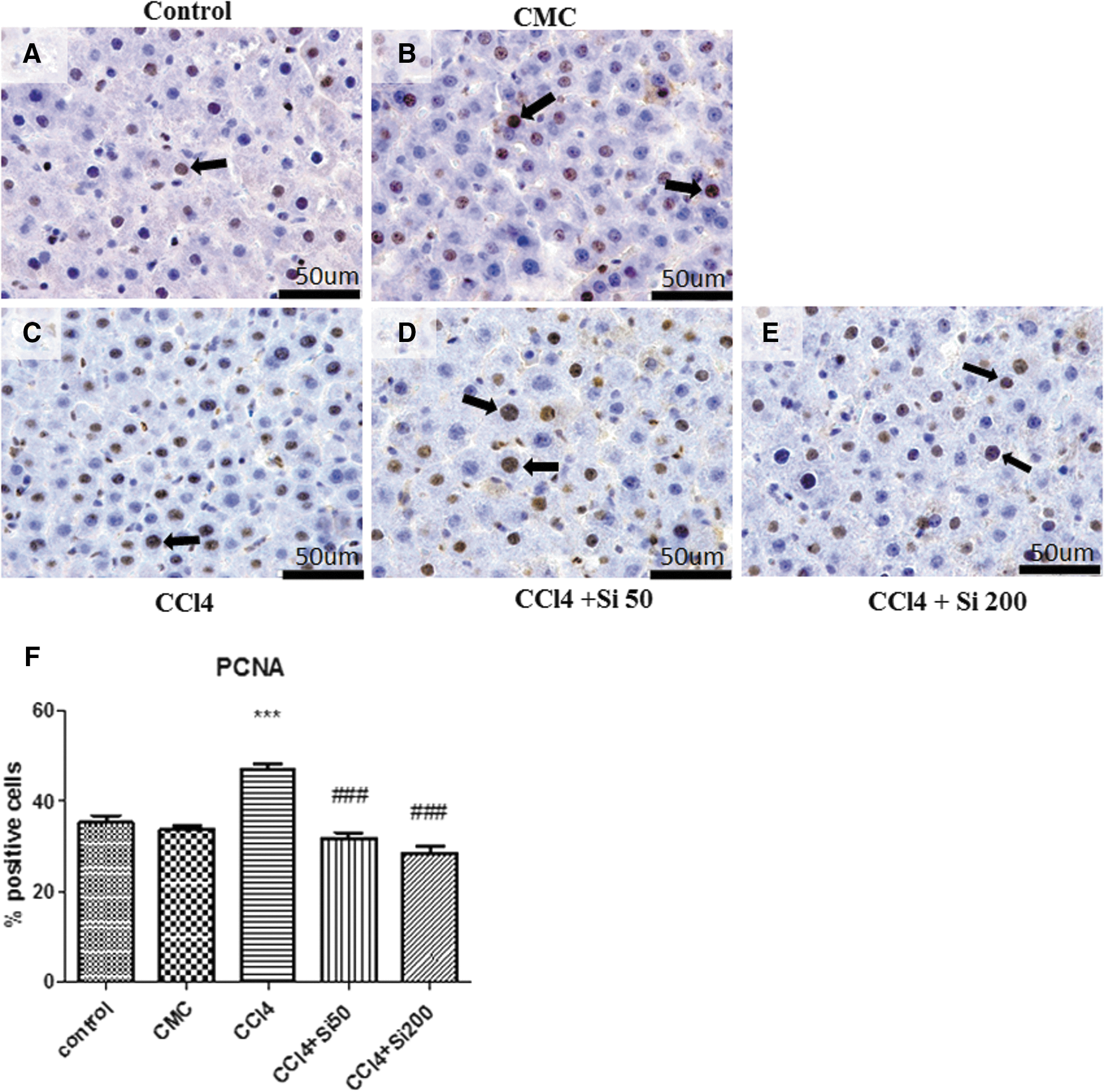
Effects of Silymarin administration upon the PCNA expression in the hepatic parenchyma of Wistar rats, which were treated for 8 weeks with CCl4 followed by the discontinuation of the hepatotoxic administration.
Discussion
Liver injury induced by CCl4 administration is a widely used model, not only to induce hepatic fibrosis, but also for screening the hepatoprotective potential of different drugs. 8
Our study followed the effects of administering Silymarin, a hepatic protector substance, known for its antioxidant and anti-inflammatory properties, on the hepatic alterations, especially liver fibrosis, obtained through the administration of CCl4 for 8 weeks. We followed both the changes produced by simply stopping the toxic administration, corresponding to a spontaneous resolution, and those caused by the administration of two doses of Silymarin (50 and 200 mg/b.w.) after the toxic administration was discontinued. In our previous publication, 6 using the same experimental model, the fibrosis score was assessed at 2 weeks (0), 3 weeks (1), 4 weeks (3), and 8 weeks (3) of CCl4 administration. In the present study, the fibrosis score after the CCl4 discontinuation was 2.6 ± 0.54, whereas under the action of 50 and 200 mg/b.w. Silymarin, it was 1 and 0.2 ± 0.04, respectively. The difference between the fibrosis score at 8 weeks of CCl4 administration and the values obtained after the toxic administration was discontinued, with or without Silymarin administration, is a confirmation not only of the mild spontaneous resolution, but also of the Silymarin contribution to the acceleration of this process.
But what are the mechanisms triggered by the CCl4 administration in the first place (Supplementary Fig. S1; Supplementary Data are available online at
The next step is the activation of NF-κB. Oxidative stress is one of the factors which lead to NF-κB activation. 11 In return, the active form of NF-κB enhances the transcription of proinflammatory cytokines, such as TNF-α and IL-6, among other factors. The cytokines, in return, will continue to activate NF-κB. This will contribute to inflammatory amplification and liver injury. Thus, oxidative stress generation, through NF-κB and inflammation, will determine liver injury. 12
In our experiment, Silymarin determined an important decrease of hepatic IL-6 level and MAPK activation, the higher dose of 200 mg/b.w. Silymarin being more effective as an anti-inflammatory agent.
The same effects were obtained upon the reduction of NF-κB expression in the liver, 200 mg/b.w. Silymarin having a more powerful impact, with better results even upon the active form of NF-κB. Silymarin reduces the inflammation produced by CCl4 administration, and the reduction of proinflammatory cytokines reduces, in return, the activation of NF-κB. In our study, Silymarin administered during the two last weeks of the experiment determined a reduction of the active form of NF-κB (pNF-κB), even though we did not obtain a similar effect on the inactive form (NF-κB). It is possible that Silymarin increases the NF-κB inhibitors, which will then maintain the nuclear factor NF-κB in its inactive form. It is also possible that Silymarin reduces the active form of NF-κB and this reduction slows down the dissociation of NF-κB from its inhibitors, which in turn will reduce the process of its translocation into the nucleus and will be responsible for the reduced expression of proinflammatory cytokines.
The injuries induced to hepatocytes will then determine their destruction. Dead hepatocytes will be engulfed by Kupffer cells, which will contribute to the activation of HSC. In fact, the trigger event for the initiation and progression of liver fibrosis is the activation of HSC. Reactive oxygen species and some cytokines, especially TGF-β1, are among the most potent activators of HSC. 13
Activation of HSC promotes alterations of the ECM, including synthesis, degradation, and even remodeling at this level. 14 CCl4 administration induces the reorganization of the cytoskeleton and the differentiation of HSC into myofibroblast-like cells 15 and also induces the expression of TGF-β1 and α-SMA. Silymarin has the capacity to inhibit the TGF β1-mediated pathways, which plays a key role in the progression of liver fibrosis. 16
The antifibrotic effects of Silymarin in vivo could be explained by the changes occurred in the expression of the genes involved in the cytoskeleton organization and the mitochondrial electron transfer chain. 16
It is also proven that Silymarin has a direct antifibrotic activity by retarding or even inhibiting the activation of HSC, 17 with the downregulation of the expression of cytoskeleton component genes, and thus ameliorating the fibrosis of the liver. 16
In our experiment, we obtained a significant reduction of TGF-β1 and also of α-SMA expression, probably due to the inhibition exerted by Silymarin directly upon the HSC. We would expect a dose-dependent effect, but our results showed that the usual, therapeutic dose of Silymarin has a better antifibrotic effect than a higher dose. The results are in concordance with our latest research, 3 where we found a similar effect for this dose during the early phases of fibrosis. We do not yet have a solid explanation for this result. We have also obtained a reduction of hyaluronic acid in liver content under the action of Silymarin administration, with no differences between the two doses.
MMPs play a role in the inhibition of hepatic fibrosis. In experimental models, a spontaneous resolution of fibrosis was emphasized, once the toxic agent had been removed. 5 However, the reversibility of fibrosis seems to be correlated with the stage of fibrosis, since the alterations occurred during established cirrhosis were considered to be irreversible. 5 Two mechanisms are responsible for the irreversibility of cirrhosis: the hepatic deposition of connective tissue, due to not only increased production but also to deficient degradation of ECM and the impaired capacity of the liver to regenerate. 18 The loss of capacity to degrade ECM will contribute to persistent liver fibrosis.
One of the factors considered to be responsible for the loss of fibrosis reversibility is the gradual loss of expression of metalloproteinases. 8 In our experiment, the higher dose of Si 200 mg/b.w. increases the expression of metalloproteinases. Maybe, this effect could be an important one for the restoration of the reversible character of fibrosis under Silymarin administration.
Hernández-Muñoz et al. 4 also found an increased PCNA after the discontinuation of the toxin, but without effective proliferative capacity of the liver. In our experiment, Silymarin was capable of restoring the normal proliferative capacity of hepatocytes, with no significant difference between the doses. It could be a crucial step for the beneficial effect of Silymarin upon the carcinogenic potential of chronic hepatopathies evolution. In experimental models, Si reduces cellular proliferation and inhibits the progression of metastasis. 19
So the idea of administering substances with antioxidant capacity, for example Silymarin, after the discontinuation of the toxin that triggered the oxidative stress generation in the first place, could have a beneficial impact on the restoration of liver functions after this toxic injury.
In our study, Silymarin was administered after the discontinuation of the administration of the toxin and was capable of reducing inflammation in the liver. The fibrosis alterations were also improved by Silymarin administration, with decreased levels of hyaluronic acid content, α-SMA, TGF-β1, and the increase of MMP-9 activity. In the meantime, in our experiment, Silymarin was able to restore toward normal values the capacity of liver cell proliferation. The anti-inflammatory and antifibrotic capacities were more important for the higher dose of 200 mg Si/b.w. than for 50 mg Si/b.w. Tsai et al. 20 have also demonstrated that the administration of 200 mg/b.w. Silymarin in 8 weeks CCl4-induced liver fibrosis, after the discontinuation of toxin, decreases aspartate amino transferase (ASAT), alanine amino transferase (ALAT), and also the α-SMA in the liver, with the potential to increase the resolution of liver fibrosis in rats. Also, a recent study 21 focused especially on oxidative stress alterations and anti-inflammatory actions of Silymarin and Taurine administration, after the discontinuation of CCl4 4 week's treatment, confirmed the capacity of Silymarin to reduce oxidative stress parameters, proinflammatory cytokines, and TGF-β1.
Taking into account all these results, in Conclusion Section, we consider that Silymarin is a potent candidate to improve the course of liver fibrosis and hepatic alterations after a hepatotoxic injury and can interfere with the mechanisms involved in liver fibrosis progression, higher doses than the usual therapeutic dosage proving a better benefit.
Footnotes
Acknowledgments
The authors are grateful to Dr. Simion Baltes for the Silymarin extract. They thank Mr. Remus Moldovan for animal handling and Mrs. Nicoleta Decea for the assessment of biochemical parameters.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.