
Abstract
The root and rhizome extract of Rhodiola rosea has been extensively used in traditional medicine to improve physical and mental performance and to protect against stress. We, and others, have reported that R. rosea can extend lifespan in flies, worms, and yeast. We also previously found that the extract can act independently of dietary restriction (DR), a treatment that can extend lifespan in a range of model organisms. In flies, DR is implemented through a reduction in dietary yeast content. Here, we report that the ability of R. rosea extract to extend lifespan in flies is dependent on the carbohydrate and caloric content when supplemented with a simplified diet composed of yeast and sucrose. R. rosea extract elevated the sugar content in flies and down-regulated hexokinase expression, suggesting that it perturbs carbohydrate metabolism in flies. In our previous studies, bananas, barley malt, and corn syrup provided dietary carbohydrates, and R. rosea extract could extend lifespan with a range of caloric levels. We conclude that the lifespan-extending effect of R. rosea extract in flies is dependent on dietary carbohydrate and caloric contents coupled with an interaction with complex dietary components present in bananas, barley, or corn.
Introduction
R
Currently, the most effective and robust way to delay aging and extend lifespan in model organisms is dietary restriction (DR), which is the reduction of total caloric intake without malnutrition. 5 We previously demonstrated that R. rosea extract acts through a mechanism independent from DR to extend lifespan. 6 A recent study, however, has suggested that R. rosea extract may extend lifespan in flies only under dietary conditions with low-protein-to-carbohydrate (P:C) ratios (i.e., high-dietary carbohydrate content). 7 Furthermore, inhibition of carbohydrate metabolism has been shown to extend lifespan in worms and mice presenting a plausible mechanism for R. rosea extract. 8 In addition to carbohydrates, protein content, as well as the P:C ratio, in the diet has been suggested to play a pivotal role in Drosophila lifespan. 7 This is somewhat in disagreement with our previous findings that R. rosea extract increased lifespan in Drosophila independently of the varying protein content in their diet. 6 Collectively, the aforementioned points underscore an important interplay between the type of carbohydrate and protein dietary contents in the lifespan of Drosophila.
In this study, we directly test the interaction of R. rosea extract with dietary carbohydrate contents, and determine its effect on fly body composition and on the expression levels of key regulatory enzymes in carbohydrate metabolism. The experiments were conducted with a simplified diet with energy supplied only by yeast and sucrose. Depending on the carbohydrate and caloric content of the diet, R. rosea extract elevated fly body sugar content and inhibited the expression of the glycolytic enzyme hexokinase. The ability of the extract to extend lifespan on this simplified diet was also dependent on both the carbohydrate and caloric content in the diet. Thus, in flies, the physiological effects of R. rosea extract are intertwined with the metabolism of carbohydrates.
Materials and Methods
Feeding and lifespan assays
The w1118 flies were obtained from the Bloomington Drosophila Stock Center at Indiana University. R. rosea (SHR-5) extract was obtained from the Swedish Herbal Institute (SHI), and UPLC analysis by SHI showed 1.75% salidroside and 4.36% total rosavin content. Flies were on a simple diet, with yeast as the protein source and sucrose as the carbohydrate source based on the methods described in Jafari et al. 2 The following feeding conditions with different P:C ratios were tested: (1) high-carbohydrate diet (1:1 P:C ratio) with 9% protein and 9% sucrose, (2) low-carbohydrate diet (100:1 P:C ratio) with 9% protein and 0.09% sucrose, and (3) low-calorie diet (1:1 P:C ratio) with 4.5% protein and 4.5% sucrose. The latter diet is included to compensate for the decreased caloric content in the low-carbohydrate diet relative to the high-carbohydrate diet. The treated groups were fed with 25 mg/mL R. rosea extract, whereas the control group was fed with vehicle solution. The 25 mg/mL of R. rosea extract was chosen for this study, because this dose has consistently extended the lifespan of male and female flies. 2,6 R. rosea extract was dissolved in a yeast solution and overlaid on top of the agar-based food preparation. Flies were maintained at 23°C in an incubator under a 12 h light 12 h dark cycle for all experiments. For the lifespan studies, flies were housed 12 per 8-dram vial (six males and six females to eliminate any sex-dependent changes), and this density was maintained as long as feasible. Flies were given fresh food every 2 days and deaths were recorded at these times. Flies were also housed 12 per 8-dram vial (six males and six females) for all other experiments, independent of the total number needed, and transferred to fresh food every other day.
Measurement of protein, water, and fat content
Flies were fed for 10 days on each of the three diets with and without 25 mg/mL R. rosea extract. The flies were then collected with CO2 and weighed. For the soluble protein assay, 50 flies per sample were homogenized in 500 μL of 100 mM potassium phosphate buffer, pH 7.4. The samples were centrifuged for 10 min at 10,000 g. Protein in the supernatant was measured by reaction with Coomassie brilliant blue and correlated to a standard curve generated with bovine serum albumin and normalized to fly weight. For water content, 10 flies per sample were weighed, dried for 48 h at 70°C, and then weighed again. The difference in weights divided by the initial weight was taken to be the water content. To determine fat content, the samples used for water measurement were then incubated at RT for 24 h in 200 volumes of diethyl ether per fly volume. The ether was removed and the samples were allowed to dry, and then weighed. Fat content was taken to be the difference in the weights before and after diethyl ether treatment divided by the initial weight (before drying at 70°C).
Gene expression assays
Approximately 400 flies were fed for 10 days with high-carbohydrate, low–carbohydrate, and low-calorie diet with and without 25 mg/mL R. rosea extract and frozen in groups of 10. RNA was extracted with TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. Samples were treated with DNase (New England Biolabs, Ipswich, MA) at 37°C for 10 min to remove contaminating DNA. DNase was heat inactivated by incubation at 75°C for 10 min in the presence of 5 mM EDTA. RNA was then purified by use of the RNeasy kit (Qiagen, Hilden, Germany). RNA quantity and quality (260/280 > 2.0) were measured by spectrophotometry. One milligram of RNA from each sample was converted to DNA by the iScript cDNA synthesis kit (Bio-Rad, Hercules, CA). Samples were diluted 100-fold. Quantitative PCR was performed on a MiniOpticon real-time PCR system with SYBR green dye (Bio-Rad). Relative amplification was calculated by the threshold cycle of each respective gene divided by the threshold cycle of the reference gene, RNA polymerase II. Primer sequences, listed in Supplementary Table S1 (Supplementary Data are available online at
Hexokinase activity
Four hundred eighty flies were fed for 10 days on a high-carbohydrate diet with and without 25 mg/mL R. rosea extract and frozen in groups of 50. Flies were homogenized in 2 mL ice-cold 50 mM potassium phosphate buffer, pH 7.2, and centrifuged at 4°C for 10 min at 10,000 g. Hexokinase activity was measured in the supernatant by using a commercially available kit (cat no. MAK091; Sigma-Aldrich) according to the manufacturer's instructions. The nicotinamide adenine dinucleotide produced was detected through a colorimetric reaction at 450 nm. The measured activity was normalized to total soluble protein. Hexokinase activity was reported as absorbance at 450 nm/mg soluble protein.
Statistical analyses
Statistical analyses were conducted using Prism software (GraphPad, La Jolla, CA). The tests used and sample sizes for each experiment are indicated in the figure captions. A P value of less that .05 was considered to be statistically significant.
Results
The ability of R. rosea extract to extend lifespan was tested under varying carbohydrate content to determine whether there was an interaction between dietary carbohydrates and the extract. We tested the following feeding conditions with different P:C ratios listed in Table 1. Decreasing the caloric content of the diet increased fly lifespan (Fig. 1A and Table 2). This occurred whether the carbohydrates in the diet were drastically reduced or both protein and carbohydrate contents were reduced equivalently (Fig. 1A and Table 2). R. rosea extract only improved survival on the high-carbohydrate diet (Fig. 1B–D and Table 2), although it did increase mean lifespan on the low-carbohydrate diet (Table 2).
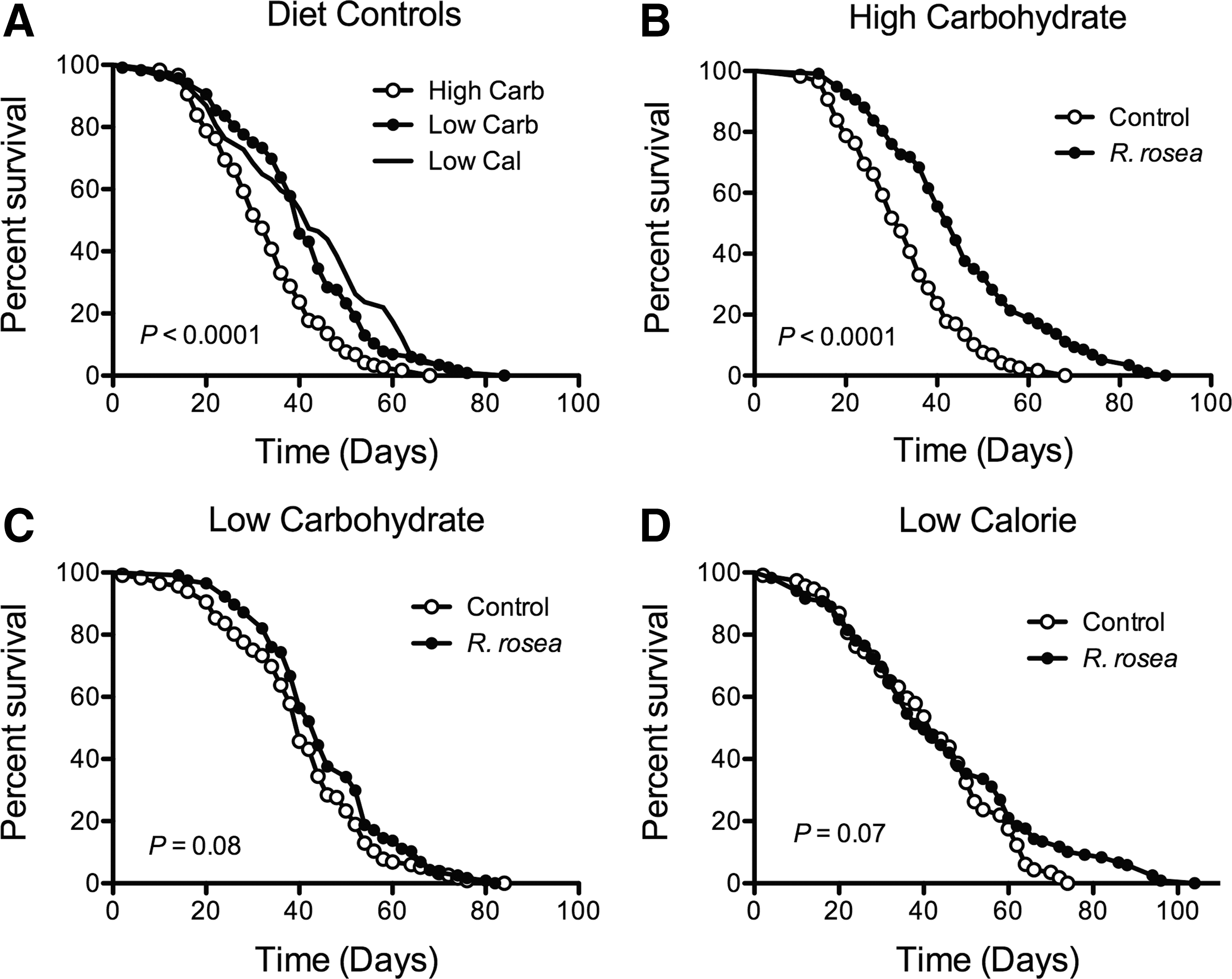
Survival of control and Rhodiola rosea extract-fed flies on various diets. Decreasing the carbohydrate or caloric content in the diet increased fly lifespan, n = 118 high carbohydrate, 116 low carbohydrate, 114 low calorie
Mean lifespans are mean ± SEM. Units are days.
P values were calculated by one-way ANOVA, Bonferroni post-test.
P values were calculated by t-test versus respective controls.
ANOVA, analysis of variance; SEM, standard error of the mean.
We measured water, sugar, soluble protein, and fat content in control and R. rosea extract-treated flies to determine how changing protein and carbohydrate contents of the diet with and without the treatment of R. rosea extract would affect these metabolic parameters. R. rosea extract decreased water content in flies on the low-carbohydrate diet, but had no effect on the other two diets (Fig. 2A). However, the extract elevated the sugar content of the flies on both the low-carbohydrate and low-calorie diets (Fig. 2B). R. rosea extract had no effect on protein or fat contents in the flies (Fig. 2C,D).
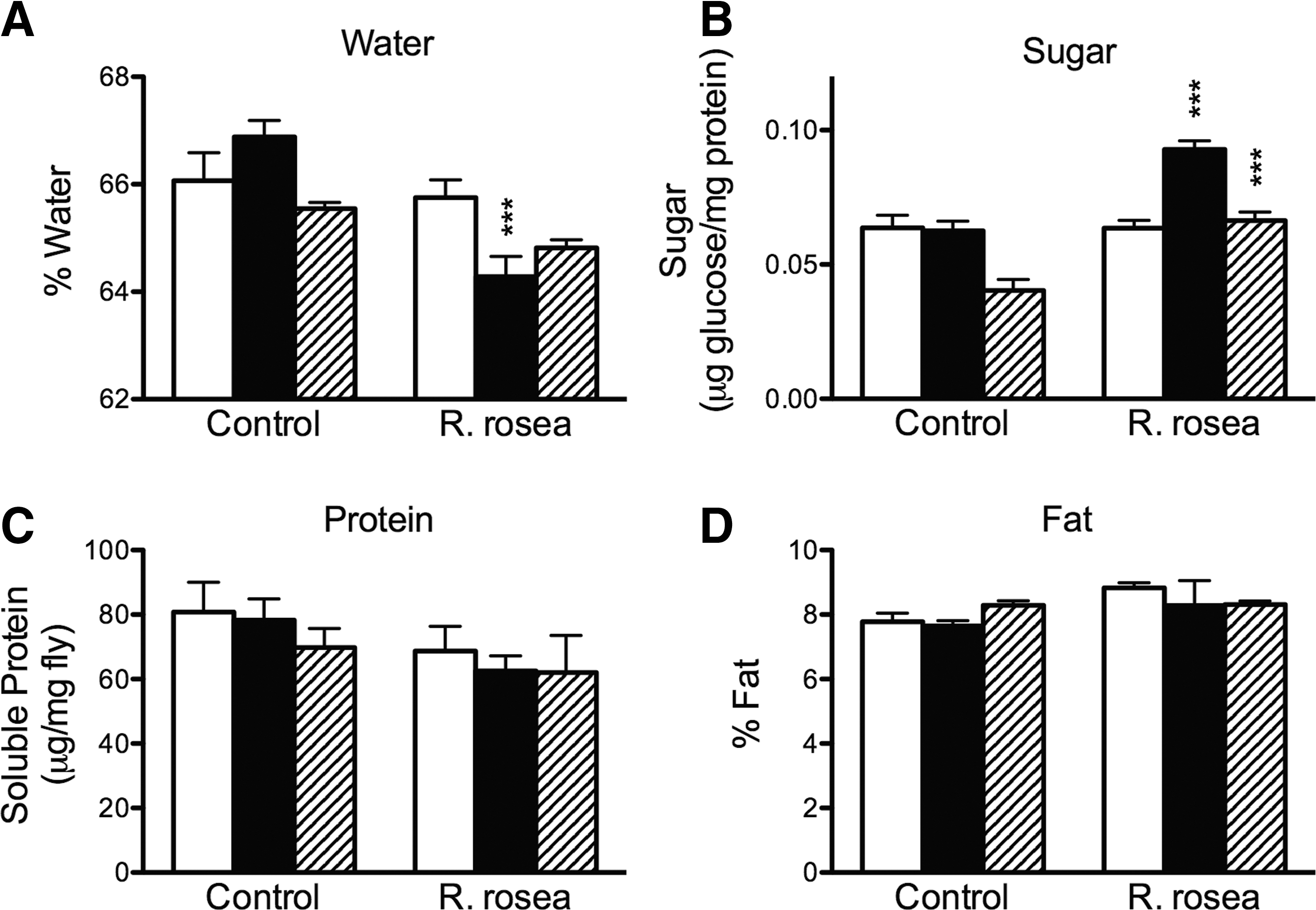
The effect of R. rosea extract and diet on body composition in flies. R. rosea extract decreased water content in flies fed the low-carbohydrate diet compared to control low-carbohydrate-fed flies
We examined the expression levels of rate-limiting enzymes in carbohydrate metabolism to determine the mechanism of action of R. rosea extract. The relative expression of hexokinase (glycolysis), glucose-6-phosphate dehydrogenase (pentose phosphate pathway), phosphofructokinase 1 (glycolysis), glycogen synthase (glycogen synthesis), glycogen phosphorylase (glycogen degradation), and fructose-1,6-bisphosphatase (gluconeogenesis) was compared between control and R. rosea-fed groups (Fig. 3). Among the six genes examined, R. rosea extract down-regulated hexokinase on the high- and low-carbohydrate diets, but not on the low-calorie diet (Fig. 3A). A similar effect may have occurred for glucose dehydrogenase, although this was not statistically significant (Fig. 3B). Despite the apparent down-regulation of hexokinase transcript, we found no decrease in enzyme activity as a result of treatment with R. rosea extract (Fig. 4).
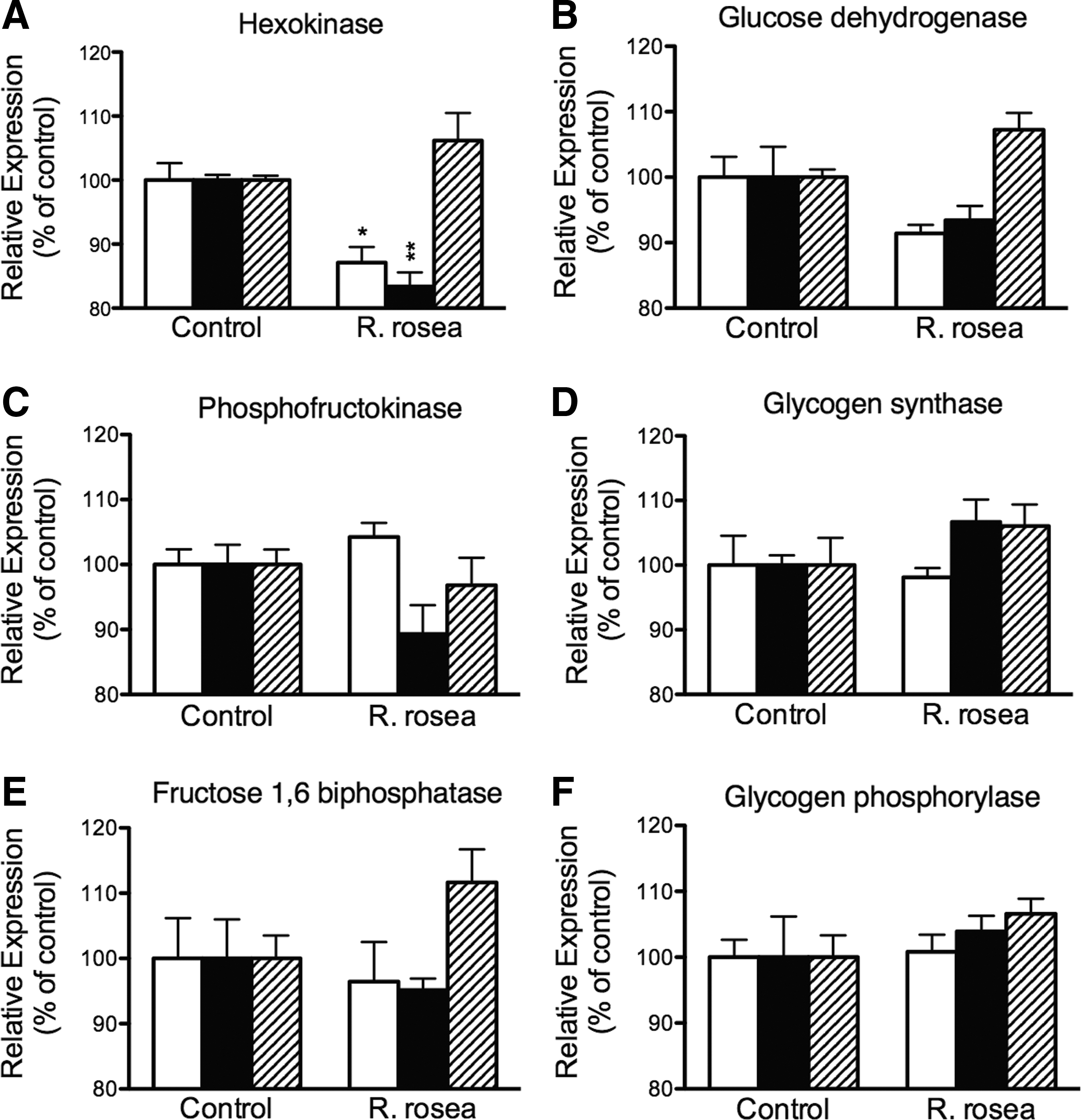
The effect of R. rosea extract on the expression levels of genes involved in carbohydrate metabolism. R. rosea extract down-regulated hexokinase when fed with the high- and low-carbohydrate diets, but not the low-calorie diet

Hexokinase activity in control flies and flies fed R. rosea extract. P = .98, t-test, n = 8 groups of 50 flies per bar.
Discussion
We, and others, have found that the root and rhizome extract of R. rosea can extend lifespan in model organisms, including yeast, worms, and flies. 2 –4,6 Although its molecular mechanism is not known, our previous findings suggest that it works independently of DR and the major aging pathways such as insulin and insulin-like signaling, the target of rapamycin, and the sirtuins. 6 Recently, it was reported that the ability of R. rosea extract to extend lifespan in flies was dependent on the P:C ratio of the food. The extract only worked at P:C ratios of less than 1; at a ratio of 20:1, R. rosea extract had a negative effect on lifespan. 7 This is seemingly at odds with our previous findings, where R. rosea extract extended fly lifespan at a P:C ratio of 1:1, just as well as it did at 1:90. 6 This observation may be explained by the fact that we used a complex carbohydrate diet, where the carbohydrates were derived from bananas, barley malt, and corn syrup. We also used a constant amount of sugar at 9%, and never investigated P:C ratios as high as Gospodaryov et al., 7 until now. Here, we examined R. rosea extract at P:C ratios of up to 100:1 and found that its ability to extend lifespan is indeed reduced or eliminated when the simple carbohydrate content in the fly diet is drastically decreased.
DR can be defined as a reduction in caloric intake without malnutrition. It is the most robust method for extending lifespan and improving health in model systems, both in breadth and magnitude of effects. 5 Similar to R. rosea extract, the mechanism of action of DR is not known. However, the reduced caloric intake, due, in part, to a decreased intake of carbohydrates, has been proposed to result in a lower concentration of toxic glycolytic intermediates. 9 Down-regulation of glycolytic enzymes is also a known effect of DR. 10 Therefore, drugs that block carbohydrate metabolism might be predicted to extend lifespan. 11 This has been found in worms and mice. In the worm C. elegans, feeding of compounds that inhibit glycolysis, glucosamine, or 2-deoxyglucose extends lifespan. 8,12 The former also extends lifespan in mice, whereas the latter was found to be toxic in rats. 8,13 Thus, interfering with carbohydrate metabolism is a plausible mechanism for enhancing lifespan and may replicate some features of DR.
Several findings have suggested that R. rosea extract may also inhibit carbohydrate metabolism. For example, we found that the extract down-regulated glyceraldehyde 3-phosphate dehydrogenase, enolase, and pyruvate kinase, and the three ageing-related Drosophila insulin-like peptides in flies. 6 We have also shown that the extract selectively eliminated cancer cells, which inherently display high-carbohydrate metabolism and up-regulated glycolysis, while leaving normal cells unharmed. 14 Finally, the putative active compounds of R. rosea extract are glycosides, 1 suggesting they are likely to be metabolized by, or inhibit, enzymes normally involved with carbohydrate metabolism. Combined with the findings of Gospodaryov et al., 7 it is tempting to imagine that R. rosea extract acts to down-regulate carbohydrate metabolism, which will result in its ability to extend lifespan in flies and other model species. This is not contrary to our previously reported findings, as the extract in that study was shown to work independently of dietary protein content. 6 To directly test whether R. rosea extract altered fly carbohydrate metabolism, we examined the ability of the extract to extend lifespan on three different diets of varying P:C ratios and caloric contents (Table 1). Interestingly, a reduction in caloric content at 1:1 P:C ratio was just as effective in extending fly lifespan as was the 100-fold reduction of carbohydrates from the diet (Table 2). However, R. rosea extract could not extend lifespan on the low-calorie diet and did not increase overall survival on the low-carbohydrate diet, although there was a significant increase in mean lifespan (Table 2). Thus, our findings here essentially agree with Gospodaryov et al. 7 that the effect on lifespan by R. rosea extract is dependent on the dietary nutrient content.
Not only was the lifespan effect of R. rosea extract dependent on the dietary carbohydrate content, but the extract also altered the carbohydrate content within the flies (Fig. 2B). The extract elevated fly sugar content when either the carbohydrate or caloric content of the diet was decreased. This could have resulted from an increased carbohydrate storage response induced by R. rosea extract due to the decreased calories. The decreased water content shown in Figure 2A may be due to displacement by the elevated sugar content. The root and rhizome extract of R. rosea also down-regulated hexokinase expression, on both the high-carbohydrate and low-carbohydrate diets, but not on the low-calorie diet (Fig. 3A). Despite this, there was no detectable effect on hexokinase enzyme activity. The reason for this observation may be due to limitations in the hexokinase assay. For example, there could have been a moderate decrease in activity, but we were unable to measure it. Alternatively, the decrease in RNA levels may have not resulted in decrease in enzyme activity due to altered translation control or enzymatic regulation. Nevertheless, these results combined with our previous findings do show that R. rosea extract can down-regulate glycolysis and perturb carbohydrate metabolism in the fly.
An important difference between this work and our previous study is that the dietary carbohydrate used in this study is sucrose. In our prior work, bananas, barley malt, and corn syrup provided a complex mixture of glucose, fructose, sucrose, and maltose. Previous work has shown that the type of carbohydrate can directly affect Drosophila lifespan, which may be a confounding factor. 15 This complex mixture of the dietary carbohydrate botanicals also provides a much richer supply of vitamins, minerals, phytochemicals, and other nutrients. Since the effect of R. rosea extract is minimized or eliminated on either the reduced calorie or reduced simple carbohydrate diets in this work, it is likely that the extract interacts with, or requires, some component of the added botanicals to mediate its effects in flies, or is dependent on a sugar other than sucrose. In future studies, we plan to add individual components of our complex diet to this simple diet to identify which dietary components of a more complex carbohydrate regimen play a role in the interaction of carbohydrates and lifespan. We will also test the action of R. rosea extract on different types of dietary sugars.
In summary, we previously found that the root and rhizome extract of R. rosea can extend fly lifespan independent of the dietary protein content and caloric content. 6 Whereas here we found that this effect is dependent on the carbohydrate or caloric content of the diet. A significant difference between these two studies is the type and source of dietary carbohydrates used. We conclude that the lifespan-extending effect of R. rosea extract in flies is dependent on the caloric and carbohydrate contents of the diets. However, there must also be an interaction between R. rosea extract and the source or type of carbohydrate present in the diet.
Footnotes
Acknowledgments
We thank Hoang M. Pham and Hanh Pham for technical assistance. This work was supported, in part, by the Anne S. Chatham Fellowship in Medicinal Botany awarded to T.E.L.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.