
Abstract
Vitamin D has been known for its anti-inflammatory properties. Extracts derived from Lentinula edodes (Shiitake) edible mushroom exert an anti-inflammatory effect. These extracts contain high levels of ergosterol, which converts into ergocalciferol (vitamin D2) following exposure to ultraviolet light, followed by absorption and hydroxylation into the active form 25-hydroxyvitamin D [25(OH)D]. To determine the anti-inflammatory effect of overexpression of vitamin D in edible mushrooms, L. edodes mushrooms were exposed to ultraviolet-B light, freeze-dried, followed by measurement of vitamin D2 contents, in their dry weight. C57B1/6 mice were orally treated with vitamin D2-enriched or nonenriched mushroom extract prior and during concanavalin A-immune-mediated liver injury. Exposure to ultraviolet light increased vitamin D2 content in Shiitake edible mushrooms. Following feeding of vitamin D-enriched mushroom extracts to mice with immune-mediated hepatitis, a significant decrease in liver damage was noted. This was shown by a decrease in alanine aminotransferase and aspartate aminotransferase serum levels, a decrease in proportion of mice with severe liver injury, and by improvement in liver histology. These effects were associated with a decrease in serum interferon gamma levels. A synergistic effect was noted between the anti-inflammatory effect of the mushroom extracts and that of vitamin D. Oral administration of vitamin D-enriched L. edodes edible mushroom exerts a synergistic anti-inflammatory effect in the immune-mediated hepatitis. The data support its potential use as safe immunomodulatory adjuvant for the treatment of HCV and nonalcoholic steatohepatitis.
Introduction
V
Vitamin D signaling modulates many inflammatory responses through regulation of genes that generate proinflammatory mediators, interference with transcription factors, such as NF-κB, which regulate the expression of inflammatory genes, and the activation of signaling cascades, such as MAP kinases, which mediate inflammatory responses. 4 In macrophages, 1,25D promotes antimicrobial responses through the induction of antibacterial proteins and stimulation of autophagy and autophagosome activity. In this way, variations in 25D levels have the potential to influence both innate and adaptive immune responses. 2 Cytokine signaling pathways influence the intracrine vitamin D system and either enhance or abrogate responses to 25D. 2 Vitamin D targets various tissues and cell types, a number of which belong to the immune system, such as monocytes/macrophages, DCs, as well as B and T cells, leading to individual responses of each cell type. One hallmark of these specific vitamin D effects is the cell-type-specific regulation of genes involved in the regulation of inflammatory processes and the interplay between vitamin D signaling and other signaling cascades involved in inflammation. 4 Vitamin D induces antimicrobial peptides, suppresses innate immune response, and induces Th2 cytokines and stimulation of T-regulatory T cells. 5
Extracts derived from Lentinula edodes (Shiitake) edible mushroom exert an anti-inflammatory effect. These extracts have relatively high levels of ergosterol, which converts into ergocalciferol (vitamin D2) following exposure to ultraviolet light.
The aim of the present study was to determine the effect of a vitamin D-enriched mushroom extract on the immune system by testing its effects on the immune-mediated liver damage, and exploring the possibility of a synergistic hepatoprotective effect of the above two elements—edible mushrooms and vitamin D.
Materials and Methods
Animals
Male C57BL/6 mice (11–12 weeks old) were obtained from Harlan Laboratories (Jerusalem, Israel) and maintained in the Animal Core of the Hadassah-Hebrew University Medical School. Mice were administered standard laboratory chow and water ad libitum and kept in a 12-h light/12-h dark cycle. Animal experiments were carried out according to the guidelines of the Hebrew University-Hadassah Institutional Committee for Care and Use of Laboratory Animals and with the committee's approval.
Preparation of vitamin D-enriched extract
L. edodes mushrooms were exposed, postharvest, to short pulses of ultraviolet-B light. The mushrooms were freeze-dried, milled to powder, and their vitamin D2 contents (on dry weight basis) were measured by HPLC (Bactochem laboratories, Ness Ziona, Israel).
Induction of concanavalin A hepatitis
Concanavalin A (ConA; MP Biomedicals, Santa Ana, CA, USA) was dissolved in 50 mM Tris pH 7, 150 mM NaCl, 4 mM CaCl2 and was injected into the tail vein at a dose of 500 μg/mouse (15 mg/kg) in the first experiment and 400 μg/mouse in the second experiment. Mice were sacrificed 16 h following ConA injection. Blood and liver tissue were collected for further analysis
Experimental groups
Two consecutive experiments were conducted. In the first study, four groups of mice were studied (n = 5–6 mice per group). Mice in the control group were treated 2 h before ConA injection with di distilled water. The first treated group received vitamin D oral supplement (1 IU of vitamin D) twice daily for 3 days before ConA-induced immune liver damage. The second treated group was treated orally with crude extraction derived from L. edodes edible mushrooms (7.5 mg of mushrooms) at the same timing and frequency, while the third treated group was treated with vitamin D-enriched L. edodes (7.5 mg of mushrooms containing 1.125 IU of vitamin D). In a subsequent study, four groups of mice (n = 5–7 mice per group) were treated by the same protocol detailed above, but the frequency of the oral treatment was increased from twice to thrice daily.
Assessment of the effect of treatment on liver damage
Liver enzymes
Serum was obtained from individual mice after sacrifice. The serum aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels were determined with an automatic analyzer.
Histological examination of the liver
Paraffin-embedded liver sections were prepared from each mouse. The livers were cut into 4–5-μm-thin slices and stained with hematoxylin–eosin. Slides were scored to assess the extent of liver damage using a previously described method. 6,7
Measurement of serum IFN- γ
Serum IFN-γ levels were measured using a commercially available Sandwich ELISA kit (Quantikine; R&D Systems, Minneapolis, MN, USA).
Statistical analysis
Statistical analysis was performed using the Student's t-test. A P value <.05 was considered significant.
Results
Effect of ultraviolet light irradiation on vitamin D content in the mushroom extracts
Table 1 shows that exposure to ultraviolet light increased the vitamin D2 content in the dry mushroom powder from 64.2 to 125.9 IU/g.
Effect of oral administration of vitamin D-enriched mushroom extract on immune-mediated hepatitis
Figure 1 shows that following feeding of the vitamin D-enriched mushroom extracts to immune-mediated hepatitis harboring mice, ALT serum levels decreased from 6283 U/L in the untreated control group to 2802, 3183, and 2872 U/L for vitamin D, nonenriched and enriched mushroom extract-treated groups, respectively (p ≤ .020, ≤.036, and ≤.022, respectively, between each of the treatment groups and untreated controls).
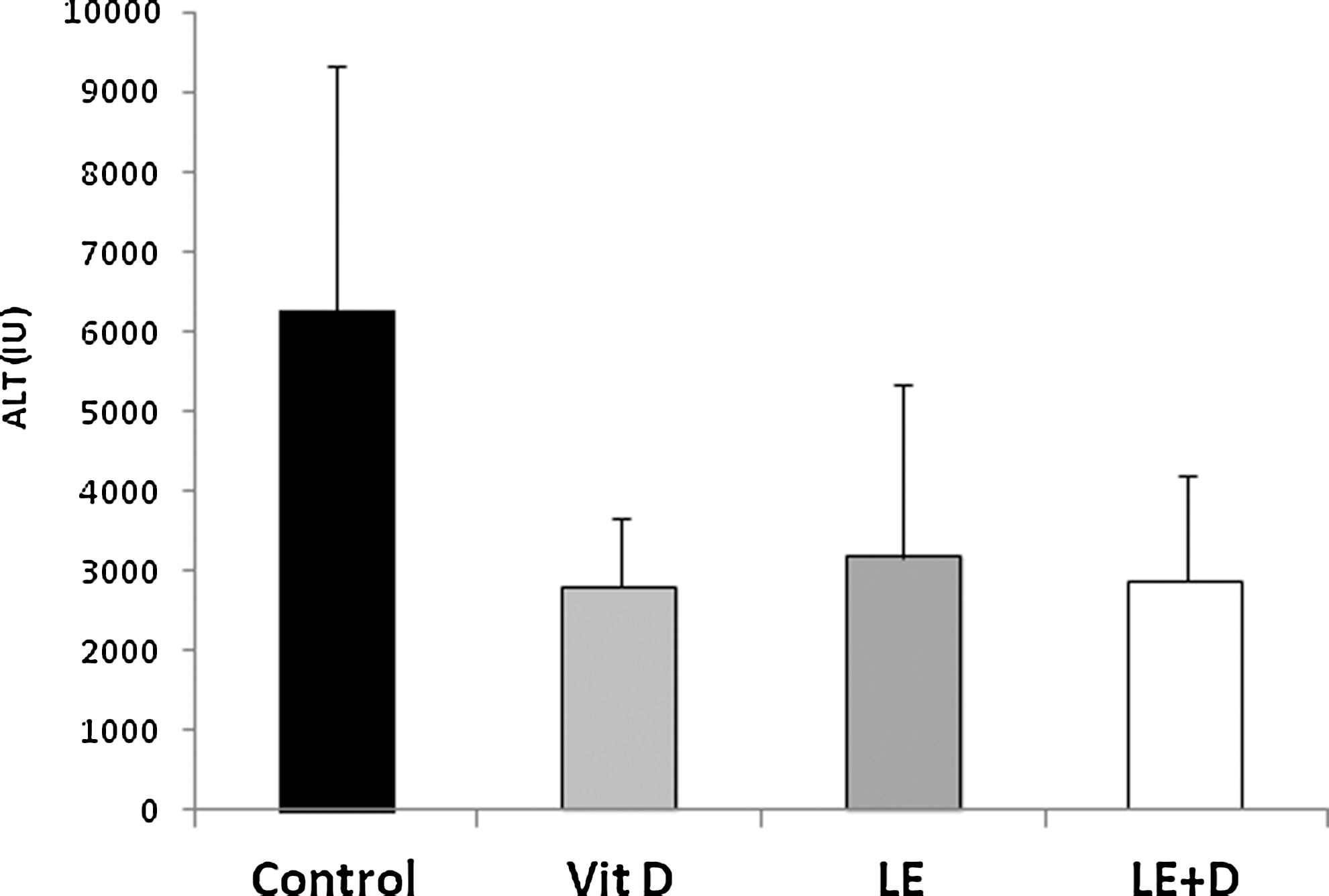
Effect of oral administration of vitamin D-enriched mushroom extract ALT serum levels in mice injected with ConA. ALT, alanine aminotransferase; ConA, concanavalin A; LE, Lentinus edodes; Vit D, vitamin D.
Figure 2 shows that the proportion of severe liver injury (defined as ALT ≥2611 U/L) was decreased from 83% in the control group to 60%, 50%, and 40% at the vitamin D, nonenriched and enriched mushroom extract-treated groups, respectively.
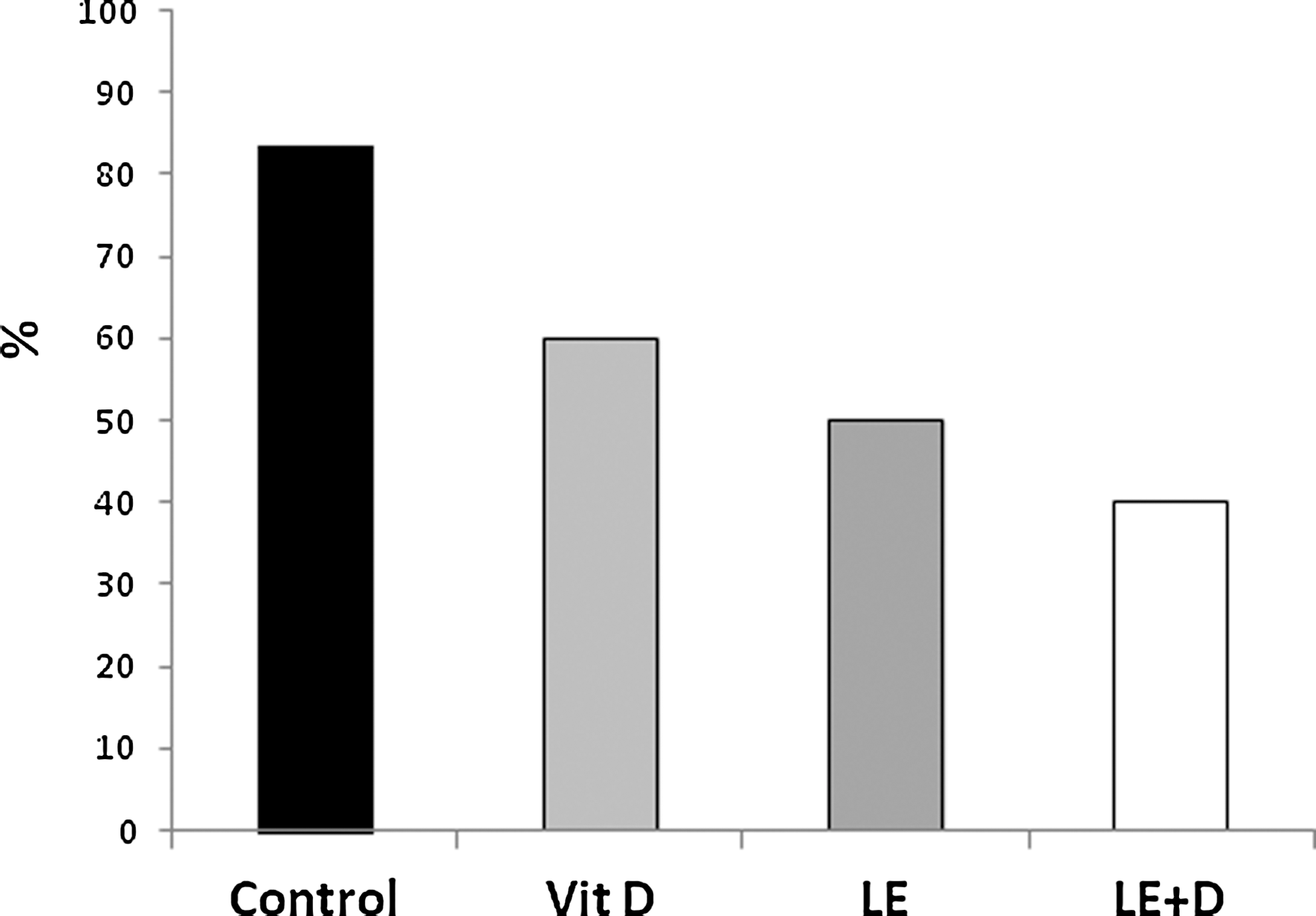
Proportion of severe liver injury (defined as ALT >2611 U/L) following oral administration of vitamin D-enriched mushroom extract ALT serum levels in mice injected with ConA.
Figure 3 displays representative pathological sections of liver biopsies showing a corresponding improvement of the immune-mediated liver injury of the first experiment, in accordance with the decrease in ALT.

Representative slides of histology of mice pathological examination of liver biopsies revealed a corresponding improvement of the immune-mediated liver injury of the first experiment.
A synergistic effect of vitamin D with the mushroom extract on alleviation of immune-mediated hepatitis
Figure 4 shows that following feeding of the vitamin D-enriched mushroom extracts to immune-mediated hepatitis harboring mice in the second study, ALT serum levels decreased from 5423 U/L for the untreated control group to 4040, 3455, and 1561 U/L for vitamin D, nonenriched and enriched mushroom extract-treated groups, respectively (p ≤ .103, ≤.049, and ≤.007, respectively, between each of the treatment groups and untreated controls).
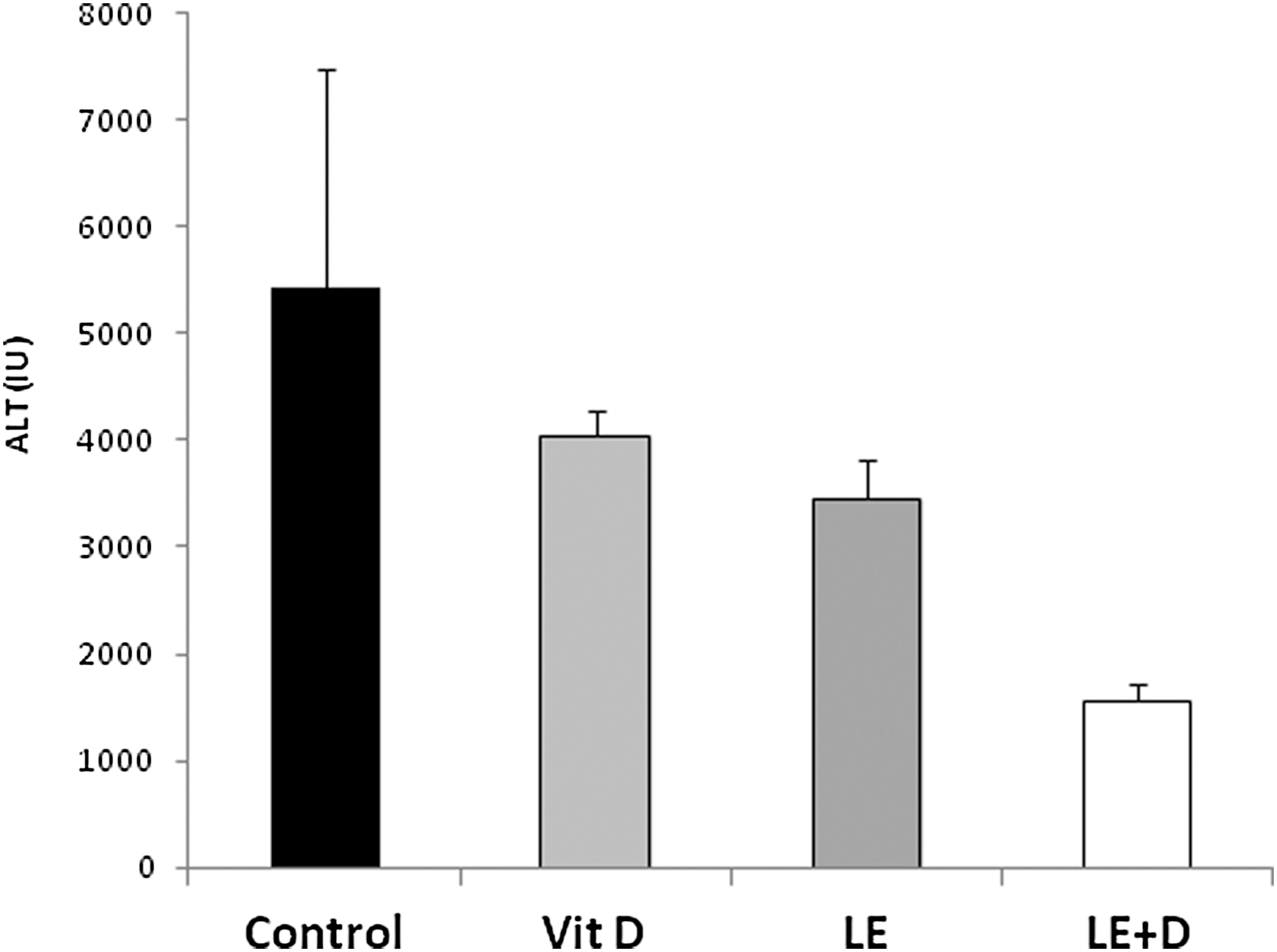
Effect of oral administration of vitamin D-enriched mushroom extract ALT serum levels in mice injected with ConA.
Figure 5 shows that the proportion of severe liver injury (defined as ALT >2000 U/L) was 100% at the first three groups (untreated, vitamin D, and nonenriched mushroom extract), but was dramatically decreased to 0% at the enriched mushroom extract-treated group, demonstrating the synergistic effect of the two different treatments, from the second experiment. A similar decrease was noted when comparing the proportion of severe liver injury by AST levers (defined as AST >2000 U/L), 100% at the first three groups and 14.3% at the fourth group (data not shown).
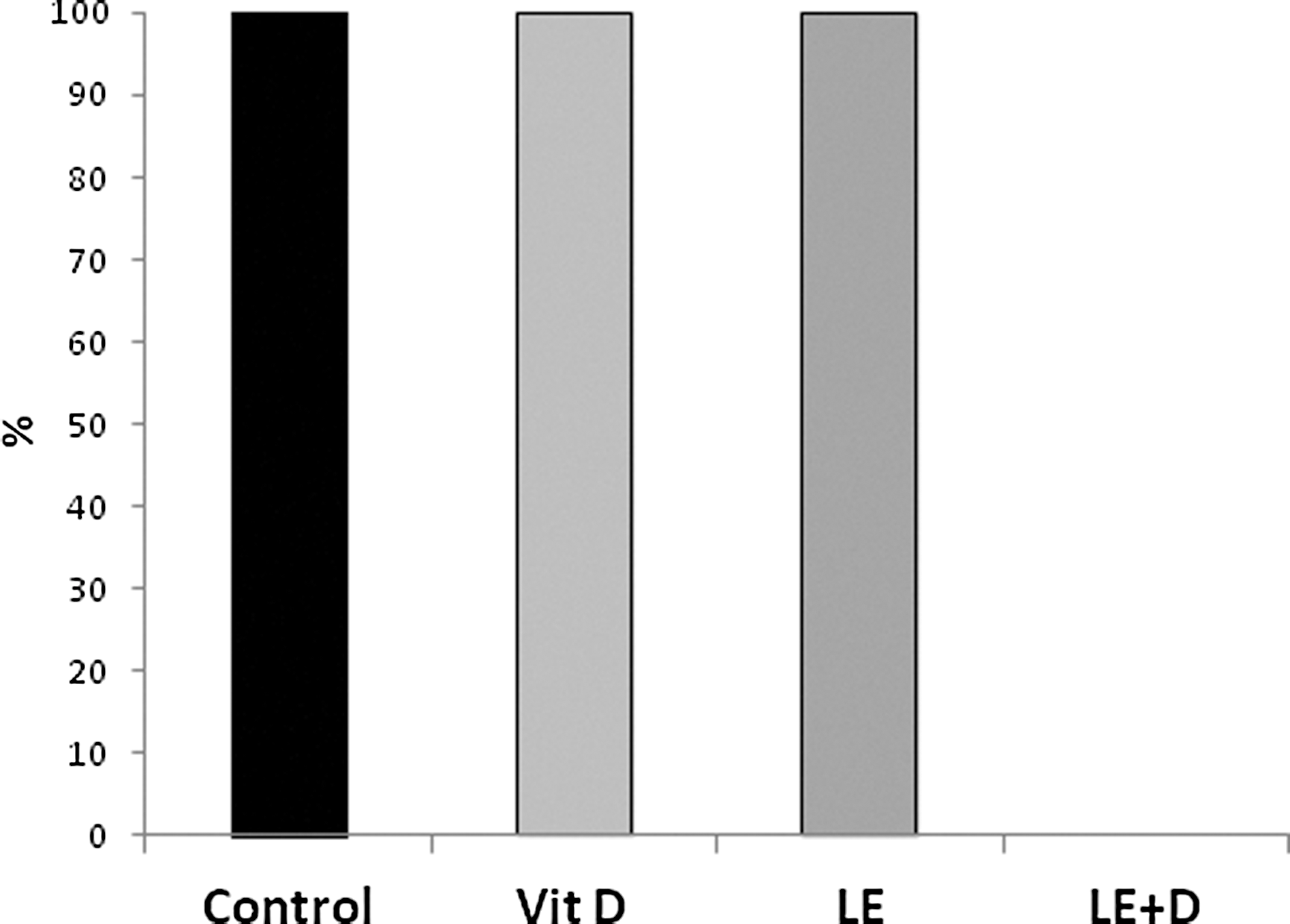
Proportion of severe liver injury (defined as ALT >2000 U/L) following oral administration of vitamin D-enriched mushroom extract ALT serum levels in mice injected with ConA.
Figure 6 shows that following feeding of the vitamin D-enriched mushroom extracts to immune-mediated hepatitis harboring mice in the first study, IFN-γ serum levels decreased from 1340 pg/mL for the control (untreated) group to 981 and 1081 pg/mL for vitamin D and enriched mushroom extract-treated groups, respectively, but increases to 1540 pg/mL for nonenriched mushroom extract-treated group.
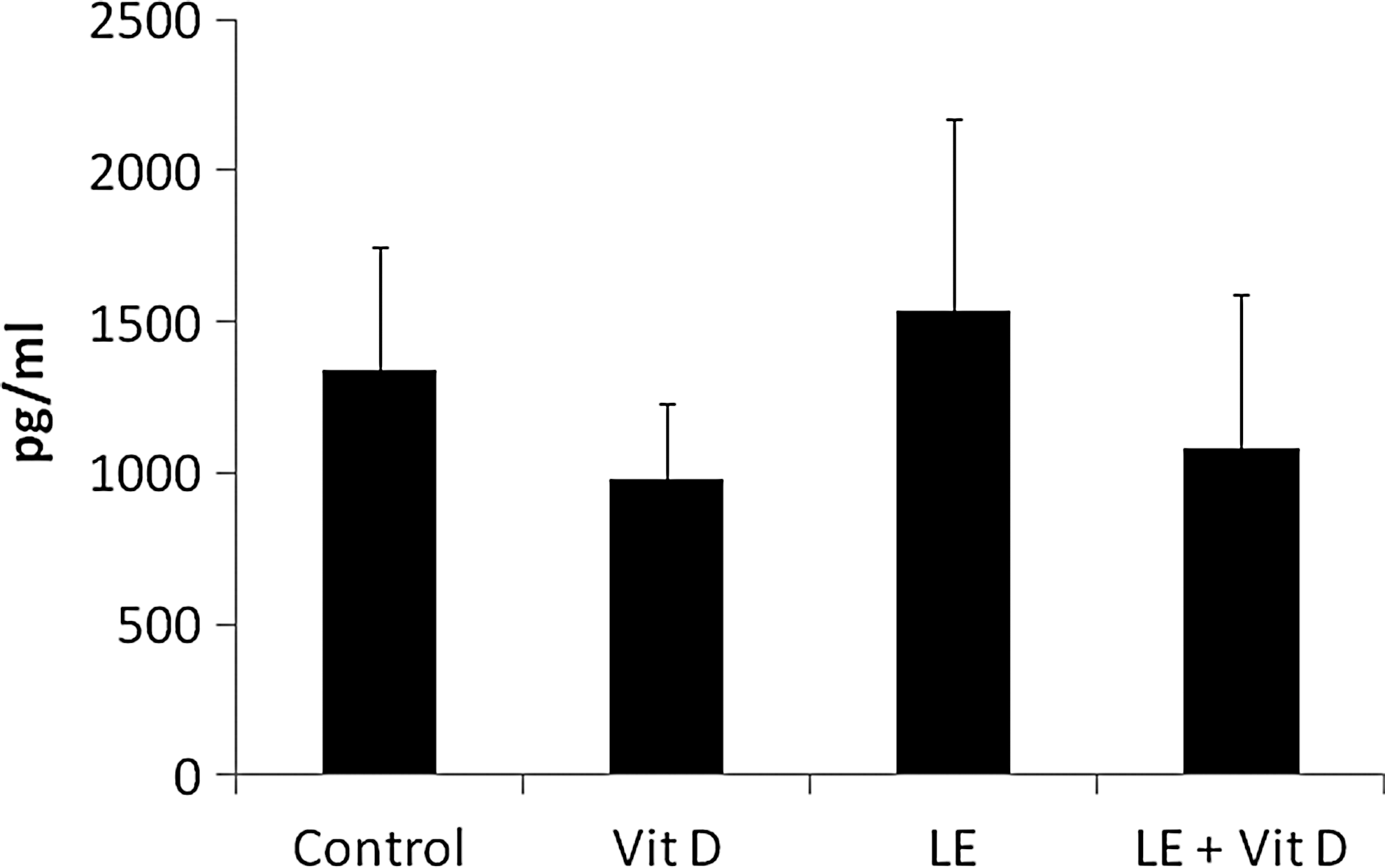
Effect of oral administration of vitamin D-enriched mushroom extract IFN-γ serum levels in mice injected with ConA as measured by ELISA.
Discussion
The data of the present study show that oral administration of L. edodes edible mushroom and/or vitamin D supplements exerts an anti-inflammatory effect in the immune-mediated hepatitis model.
Mushrooms have long been used by humans both as food and as medicine. 8 Extracts and isolated metabolites from mushrooms of different genera, such as Lentinula, Pleurotus, Auricularia, Flammulina, Tremella, Hericium, and Grifola, can modulate the immunomodulatory effect that is mediated by affecting the innate and adaptive immune pathways, 9 by activation of immune effector cells, 10 and by altering of the pro-anti-inflammatory cytokine paradigm. 9 Mushroom extracts were shown to modulate the differentiation capacity of CD4+ T cells to mature into Th1 and/or Th2 subsets, 11 and to have an effect in immune-mediated disorders. 12 Shiitake extract affected liver-mediated immune regulation by altering the NKT lymphocyte distribution and increasing intrahepatic CD8+ lymphocyte trapping, thereby leading to alleviation of immune-mediated colitis. 13 Extracts derived from L. edodes (Shiitake) edible mushroom exert an anti-inflammatory effect. Administration of Shiitake resulted in marked alleviation of immune-mediated colitis, manifested by significant improvement in the macroscopic and microscopic scores. These effects were accompanied by an increase in the number of intrahepatic natural killer T-regulatory cells. 13,14 Most of the bioactive molecules in mushrooms are polysaccharides. 15 The standard approach to studying the effect of mushrooms has been to isolate, characterize, and administer pure active constituents. However, components of a mushroom extract may have synergistic physiological effects. 16,17
Mushrooms have relatively high levels of ergosterol, which, under the influence of UV light, is converted in to ergocalciferol (known as vitamin D2). 18,19 It has been demonstrated that when white button mushrooms are exposed to ultraviolet B radiation, for a short period of time, the level of vitamin D increases to levels many times the minimum daily requirement. 20 –22 Extracts derived from L. edodes have relatively high levels of ergosterol, which converts into ergocalciferol (vitamin D2) following exposure to ultraviolet light. 19,23,24 Vitamin D2 is as effective and biologically active as vitamin D3. Vitamin D2 on consumption is converted to 25-hydroxycholecalciferol by the liver, and then to 1,25-dihydroxycholecalciferol (vitamin D3) by the kidneys. 25 The results of the present study show that exposure to ultraviolet light increased the vitamin D content in the mushroom extracts.
The data of the present study show that feeding of the vitamin D-enriched mushroom extracts alleviates immune-mediated hepatitis. This was manifested by a decrease in ALT and AST serum levels, a decrease in the proportion of mice with severe liver injury, and by alleviation of the histological liver damage. The data further support a beneficial immunomodulatory effect of vitamin D administered in combination with the mushroom extracts. The present study supports a synergistic effect between the anti-inflammatory action of the mushroom extracts and the immunomodulatory effect of vitamin D. As shown, the enriched extracts exerted a synergistic effect on alleviation of liver damage compared with the nonenriched mushroom extracts or vitamin D alone.
Vitamin D is now discovered as a highly versatile molecule with roles in immunity, cancer, infectious diseases, fibrosis, fatty liver diseases, and alcoholic liver diseases. 5 Biologically active vitamin D, 1,25-dihydroxylvitamin D3, is synthesized in two distinct systems. The first is the classic two-step hydroxylation in the liver and kidneys, which facilitates calcium absorption and homeostasis, and the second is 1,25-dihydroxylation by immune cells in response to infection and contributes to immune regulation. 5
Vitamin D is a natural immunomodulator and has well-established anti-inflammatory and immunomodulatory properties. Epidemiological data suggest a correlation between poor vitamin D status and high risk for chronic inflammatory illnesses of various etiologies, including autoimmune diseases. 26 Activation of VDRs regulates calcium metabolism, cellular growth, proliferation and apoptosis, and immunological functions. 26 Vitamin D plays a role in the development of several autoimmune diseases, based on its immunomodulatory properties. 27 VDR polymorphisms observed in some of autoimmune diseases further support a plausible pathogenic link. 26 Some of these effects are associated with the induction of immune tolerance by promoting tolerogenic DC function. A deficiency in vitamin D levels or VDRs is commonly indicated in autoimmune diseases such as multiple sclerosis, through suppression of proinflammatory IL-2, TNF-α, and IFN-γ by Th1 cells. 27 CD4+ Treg cells secrete high levels of IL-10 when stimulated in the presence of vitamin D3. Following stimulation by allergen, IL-10-secreting Treg cells inhibit cytokine secretion by allergen-specific Th2 cells in an IL-10-dependent manner. Addition of vitamin D3 with dexamethasone to cultures of steroid-resistant asthma patients' CD4+ T cells enhanced IL-10 synthesis. Vitamin D3 significantly overcame the inhibition of glucocorticoid receptor expression by dexamethasone. Administration of vitamin D3 to healthy individuals and to steroid-resistant asthmatic patients enhanced subsequent responsiveness to dexamethasone for induction of IL-10. 28 Vitamin D was also associated with induction of macrophage response to TB infection and with decreased proliferation and antibody production by B cells. 29,30
Chronic liver disease (CLD) and several related extrahepatic manifestations such as hepatic osteodystrophy, are associated with vitamin D deficiency. 31 Vitamin D undergoes hepatic 25-hydroxylation, rendering the liver critical to its metabolic activation. 31 Vitamin D deficiency is highly prevalent in CLD patients, and vitamin D levels are inversely related to the severity of CLD. Intervention studies showed improvements of CLD following supplementation.
The association between 25-hydroxyvitamin D [25(OH)D] and sustained virological response (SVR) in HCV-infected individuals was evaluated in the literature showing a high prevalence of vitamin D deficiency and high SVR rates in individuals with higher serum vitamin D levels or receiving vitamin D supplementation. 32 Vitamin D supplement improves the sensitivity to antiviral therapy for chronic HCV infection. 32 Low vitamin D status was associated with a higher likelihood of having acute liver failure and lower odds of achieving SVR following Peg-IFN/ribavirin therapy. 33 25-dihydroxyvitamin D, rather than 1,25-dihydroxyvitamin D, could directly suppress hepatitis C virus assembly. Clinical evidence has shown that vitamin D deficiency is associated with alcoholic and nonalcoholic fatty liver diseases (NAFLD). 5 Low levels of vitamin D were shown to be a predictor for the presence of advanced liver fibrosis. 34 In untreated-HCV patients, vitamin D supplementation improves the sensitivity of Peg-IFN/ribavirin therapy reducing the levels of IP-10, a biomarker for treatment response. 35 A possible explanation may be related to expression of hepatic VDR proteins, which are inversely and independently associated with the severity of liver fibrosis and inflammation in genotype 1 chronic HCV patients. 36
Similarly, vitamin D deficiency was suggested to play a role in the pathogenesis of NAFLD and nonalcoholic steatohepatitis (NASH). Studies have shown that vitamin D levels are inversely associated with NASH and fibrosis in children with NAFLD. 37 Vitamin D deficiency was found to promote NASH in animal models. 38 On multivariable linear regression with correction for age, gender, and BMI, 25-OH-D levels were found to be lower in children with NASH and lower in children with stage 1 or 2 fibrosis than in those with stage 0 fibrosis. Low plasma vitamin D concentration is an independent predictor of the severity of NAFLD. 39 Serum 25(OH)D3 levels were inversely associated with NAFLD, even in subjects with normal total body fat, suggesting a role of lower vitamin D levels in the development of NAFLD. 40 Lower serum 25(OH)D concentrations are significantly associated with NAFLD, independent of adiposity and insulin resistance. 41 Logistic regression showed that serum 25(OH)D3 levels were independently correlated with NAFLD and were inversely associated with fatty liver index in these patients. Plasma vitamin D concentration was significantly lower in NAFLD compared with healthy controls, and higher NAFLD activity scores were associated with lower plasma concentration of vitamin D. Patients with NASH had significantly lower vitamin D levels than those with steatosis alone, and lower concentrations of vitamin D were associated with greater severity of steatosis, hepatocyte ballooning, and fibrosis. 39
The present study does not explore the mechanism of the postulated synergism between the vitamin D and mushroom extract. In addition, there are several questions regarding UV treatment, among them the dose, wave-length, timing, retention of D2, and shelf life, which need to be considered.
In summary, the data of the present study show that oral administration of vitamin D-enriched L. edodes edible mushroom exerts a synergistic anti-inflammatory effect in the immune-mediated hepatitis. The data support its potential use as a safe immunomodulatory adjuvant for the treatment of HCV and NASH.
Footnotes
Acknowledgment
This work was supported, in part, by a grant from The Roman-Epstein Liver Research Foundation (to Y.I.).
Author Disclosure Statement
No competing financial interests exist.