
Abstract
Persistent inflammatory conditions can have severe pathological consequences. Although the use of nonsteroidal anti-inflammatory drugs (NSAIDs) is effective, it has side effects, particularly at the gastrointestinal level. There is then a high interest to identify natural anti-inflammatory compounds with no side effects. The anti-inflammatory and anti-nociceptive activities of hen egg lysozyme (LZ), both in its native form and modified by heat treatment, chemically or by enzymatic digestion have been tested in this study. The carrageenan-induced model in mice using native LZ or modified LZ has been applied. It was observed that LZ denatured by heat treatment at pH 6.0 presented 39.47% of inhibition of paw edema when administered at 30 mg/kg. LZ denatured with DL-dithiothreitol (DTT) presented a significant result of 42.10% inhibition of paw edema when administered at 30 mg/kg of animal weight. Modified LZ showed anti-inflammatory capacity comparable with the activity of the positive control dexamethasone. A classical model of acetic acid-induced abdominal writhing tests in mice was used to assess anti-nociceptive activity of native LZ and denatured heat treatment LZ and denatured chemical agent LZ. Finally, hydrolyzed native LZ presented 48% of inhibition of abdominal writhing in mice. Modified LZ with heat, chemical, and hydrolysis presented anti-inflammatory and anti-nociceptive activities independently of their natural enzymatic activity. These novel data point out the potential use of denatured and digested LZ as therapeutic agents and offer alternatives to the use of NSAIDs. LZ can be a natural source of anti-inflammatory and anti-nociceptive agents.
Introduction
I
Nonsteroidal anti-inflammatory drugs (NSAIDs) include a wide variety of medicines that are commonly used for the treatment of inflammatory-related diseases and pain. Although these compounds have potent activity, long-term administration is required to treat chronic diseases, which represents a risk for the patient since these drugs can cause side effects, particularly at the gastrointestinal level such as gastric and duodenal ulcers. 2 Therefore, it is important to find new anti-inflammatory agents with no or reduced side effects. Along that line, naturally occurring agents with high effectiveness and that are harmless are desirable as an alternative to chemical therapeutic agents. 3
Lysozyme (LZ) (E.C.3.2.17, LZ), with a single polypeptide chain consisting 129 amino acids, is a globular basic protein widespread in nature and characterized by hydrolyzing the β-1-4 linkage of the peptidoglycan backbone, which counts for its antimicrobial activity. 4 –6 Besides its antimicrobial properties that are exploited within the food and pharmaceutical industries where LZ is included as a preservative in various food items as an active principle in different drug preparations, 7,8 LZ has been reported to have antiviral, immunomodulating, immunostimulating, antitumoral, 9 anti-inflammatory, 10 antioxidant, 11 and anti-nociceptive properties, 12 among others. There are promising results on the anti-inflammatory activity of LZ both in vitro, LZ-ameliorated IL-8 production in Caco-2 cells stimulated with lipopolysaccharide, 13 and in vivo, LZ attenuated inflammation in a porcine model of dextran sodium sulfate-induced colitis. 10 On the other hand, relatively little is known about the anti-nociceptive activity of LZ. 14
The modification of LZ to enhance its beneficial biological properties has been a considerable focus of past and current research, but mostly centered on the antimicrobial properties of LZ. 7 Interestingly, chemically or heat-denatured and catalytically inactive forms of LZ have shown strong antimicrobial activity even higher than the one of the native protein, 15 –17 suggesting that apart from its direct bacteriolytic action, LZ has structural motifs of biological importance embedded in its folded organization.
In the present work, we aimed to explore in vivo how different treatments, including heat and chemical denaturation, as well as hydrolysis, affect the anti-inflammatory and anti-nociceptive activities of LZ compared with that of the native protein, which, to the best of our knowledge, remains unexplored.
Materials and Methods
Materials
Hen egg white LZ (58,000 U/mL), pepsin (3440 U/mg, from porcine stomach mucosa), dexamethasone, indomethacin, DL-dithiothreitol (DTT), and iodoacetamide (IAM) were from Sigma-Aldrich Co. (Saint Louis, MO, USA). All other reagents were of analytical grade.
Thermal and chemical denaturation of LZ
Heat-induced inactivation of LZ (1 mg/mL) was performed by incubation at 90°C for 20 min in 10 mM potassium phosphate buffer at pH 6. The chemical denaturation of LZ was conducted by reduction of their disulfide bonds and reaction with IAM, as previously described. 18 Briefly, solution of 1 mg/mL of LZ in a Tris-HCl buffer solution (10 mM, pH 8) was incubated with 2 mM DTT at 30°C. Aliquots (10 mL) were withdrawn at 1.5 h and immediately allowed to react with three molar equivalents of IAM with respect to DTT at 30°C for 1 h in the dark.
In both treatments, insoluble aggregates were removed by centrifugation at 3000 g for 15 min. The supernatants obtained were used in the following experiments after determination of protein content according to the bicinchoninic acid assay, using the Pierce diagnostics commercial test (Rockford, USA). The samples, after being extensively dialyzed against distilled water at 5°C, were freeze-dried for further use.
Enzymatic hydrolysis of LZ
LZ was dissolved at 5 mg/mL in 10 mM potassium phosphate buffer 10 at pH 1.2 and pepsin was added for an enzyme-to-substrate ratio of 1:20. This mixture was shaken at 37°C for 1 h. The reaction was stopped by heating at 80°C for 15 min and the pH was adjusted at 7.0 by addition of 1 M sodium hydroxide to inactive pepsin. Sample was lyophilized and stored at −20°C.
Animal studies
In vivo experiments were conducted with 8-week-old male Swiss and BALB/c mice of 25–35 g body weight that were kept at 25°C in 12-h light–12-h dark cycles in the animal facility with water and food ad libitum for 7 days before assays. The studies were carried out in accordance with current guidelines for the veterinary care of laboratory animals and were performed under the consent and surveillance of Unicamp's Institute of Biology Ethics Committee for Animal Research (number 2206-2).
Anti-inflammatory activity of LZ
The anti-inflammatory activity of native or modified LZs was assessed by inducing paw edema in the mice by carrageenan (40 μL/paw, injection). The mice were divided into four groups (n = 6). The different groups were treated orally (p.o) with native and modified LZ (3, 10, and 30 mg/kg, p.o), dexamethasone (30 mg/kg, p.o), and vehicle control (0.9% NaCl, 0.1 mL/kg/p.o.). The administration of LZ and drugs was made 1 h before injection of 40 μL of 3% freshly prepared suspension of carrageenan in normal saline in the right hind paw subplantar of each mouse. The paw volume was measured initially and then at 0, 1, 4, 6, 24, 48, and 72 h after the administration of the carrageenan injection by using plethysmometer (Panlab, Barcelona, Spain). 19 Edematous responses were expressed as an increase in paw volume (in mL), calculated by subtracting the basal volume from the actual one, presented as mean ± standard error of mean (SEM) for each treatment group, and also expressed as the inhibition percentage of paw edema compared with the negative control (0.9% NaCl, 10 mL/kg/p.o.).
Anti-nociceptive activity of LZ
The writhing test in mice was carried out using the method of Santos et al. 20 The mice were divided into four groups (n = 6). The writhes were induced by intraperitoneal injection of 0.8% acetic acid (v/v). One dose of native or modified LZ (30 mg/kg b.wt.) was administered by oral gavage to groups of six animals each, 30 min before chemical stimulus (acetic acid). Sodium chloride 0.9% (10 mL/kg) was used as negative control and indomethacin (30 mg/kg) as positive control. The number of muscular contractions was counted over a period of 20 min after acetic acid injection. Contraction of the abdominal muscle concomitant with a stretching of the hind limbs in response to an intraperitoneal injection of acetic acid 0.8% (10 mL/kg) was measured.
The percentage of protection against acetic acid-induced writhing was calculated using the following equation: percentage of protection = (Wc − Wt)/Wc) × 100.
Wc is the mean value of number of writhes in the control group and Wt is the mean value of number of writhes in the test groups.
Statistical analysis
Results are presented as mean ± SEM (n = 5). The results were analyzed by one-way ANOVA, followed by Tukey's test. GraphPad Prism 4 software was used. Different letters show statistical difference from the control group (P < .05) ANOVA and Tukey's test.
Results and Discussion
Anti-inflammatory activity of LZ
Carrageenan-induced inflammation is an acute, nonimmune, and highly reproducible process useful to evaluate the activation of inhibitors of the inflammatory cascade by NSAIDs or novel drugs. The cardinal signs of inflammation are edema, hyperalgesia, and erythema, which develop immediately following subcutaneous injection, producing a maximum level of inflammation within the first 4–5 h. 21,22
The anti-inflammatory effect of the different preparations of LZ orally administered to mice was evaluated after 4 and 72 h (Table 1). In the test of carrageenan-induced paw edema, the positive control (dexamethasone) significantly (P < .05) reduced paw edema in mice at 4 and 72 h after carrageenan injection compared with the saline solution with percentages of inhibition of 34.4% and 47.4%, respectively. Mice treated with native and modified LZ between 3 and 30 mg/kg body weight significantly (P < .05) decreased paw edema at 4–72 h after carrageenan injection compared with the negative control of the saline solution (Table 1). After 4 h of administration, when the maximum inflammatory condition is reached, all the samples of LZ exerted anti-inflammatory activity in a manner comparable with that of dexamethasone (Table 1). Although at 4 h there is no clear dose/effect, after 72 h, the highest dose of thermally denatured and reduced LZ showed higher anti-inflammatory effect than doses of 3 and 10 mg/kg.
Values (calculated compared with control using one way-ANOVA, followed by Tukey's Test); P < .05. All values are mean of individual data obtained from six mice (n = 6).
Show statistical difference from control group (P < .05).
DTT, DL-dithiothreitol; LZ, lysozyme.
After 72 h of administration of dexamethasone, the anti-inflammatory effect of dexamethasone increased (47.4% of inhibition) compared with the earlier point in time (34.4% of inhibition). Such effect was not seen in the mice treated with native LZ, where the anti-inflammatory activity was just maintained through time or decreased in the case of the lowest dose. Likewise, the anti-inflammatory activity of hydrolyzed LZ was maintained or slightly decreased after 72 h of administration of hydrolysates, but not in a dose-dependent manner. On the other hand, the anti-inflammatory effect of chemically or heat-denatured LZ after 72 h of administration was dose dependent. The dose of 3 mg/kg being less active and with a decreased activity compared with that at 4 h, the 10 mg/kg dose maintained activity through time and the highest one (30 mg/kg) had an increased activity after 72 h (39.5% for heat-denatured LZ and 42.1% for chemically denatured LZ) similar to that exerted by dexamethasone. The whole time course of inflammation was tracked in the mice treated with chemically and heat-denatured LZ, as can be seen in Figure 1. Although during the acute phase of the inflammatory response (5–6 h) all dosages of denatured LZ had remarkable effects, only the highest dose (30 mg) had an effect that in fact increased over time, with a behavior comparable with that of dexamethasone.

Anti-inflammatory effect of heat-denatured LZ at 90°C at pH 6.0
It is of interest that the anti-inflammatory activity of denatured LZ compared with that of the hydrolyzed or native protein was clearly higher, particularly at 30 mg. These results suggest that the anti-inflammatory activity of LZ is independent of its lytic capacity since the lytic activity of these denatured preparations was negligible (data not shown), but related to regions embedded in the well-packed and stable structure of LZ, which includes four disulfide bridges along with six helix regions. The unfolding of the protein, due to heating at 90°C or by DTT treatment, exposed the hydrophobic core of the protein to the media. The unfolded form of LZ, hydrophobic and positively charged, could likely interact with some of the inflammatory mediators released at different levels of cascade neutralizing with subsequent reduction in paw edema formation. For example, arachidonic acid, which plays a pivotal role in both inflammation and pain, 23 could be a target for the action of denatured LZ especially. In support of this idea, bioengineered LZ is being produced to prevent sequestration of LZ by anionic biopolymers associated with inflammation as this interaction would decrease the lytic activity of LZ, compromising to a certain extent its antimicrobial activity. 24 Hence, mixtures of native and bioengineered LZ would be suitable in cases of infection and inflammation since the former typically precedes the latter.
Anti-nociceptive activity of LZ
The anti-nociceptive activity of native LZ and its modified forms are shown in Figure 2. The mice treated with indomethacin showed a decrease of 78% in the number of writhes, which was significantly higher than that obtained with mice treated with native LZ or any of the modified forms of LZ. No significant differences were found between the anti-nociceptive activity of LZ and chemically or heat-denatured LZ. Bianchi 12 reported anti-nociceptive activity of LZ in rats, but the dosages used were higher than the ones used in the present study. Noteworthy, the hydrolyzed LZ showed the highest anti-nociceptive activity of the different LZ preparations being able to inhibit up to 48% of the writhes. Therefore, it is not likely that the enzymatic activity of LZ plays any role in its anti-nociceptive capacity rather than the peptides or fragments released during digestion with pepsin, which was previously suggested in a rat model by Bruzzese et al. 14 In fact, it has been shown in vitro that intestinal caco-2 cells are capable of retaining, at the intracellular level, LZ peptides released during digestion up to 24 h after stimulation, which may facilitate its anti-nociceptive action for a prolonged time. 10 A profound characterization of the peptides or LZ fragments released by pepsin digestion has been recently published, 25 but unveiling the mechanisms underlying the anti-nociceptive activity of hydrolyzed LZ requires further investigation.
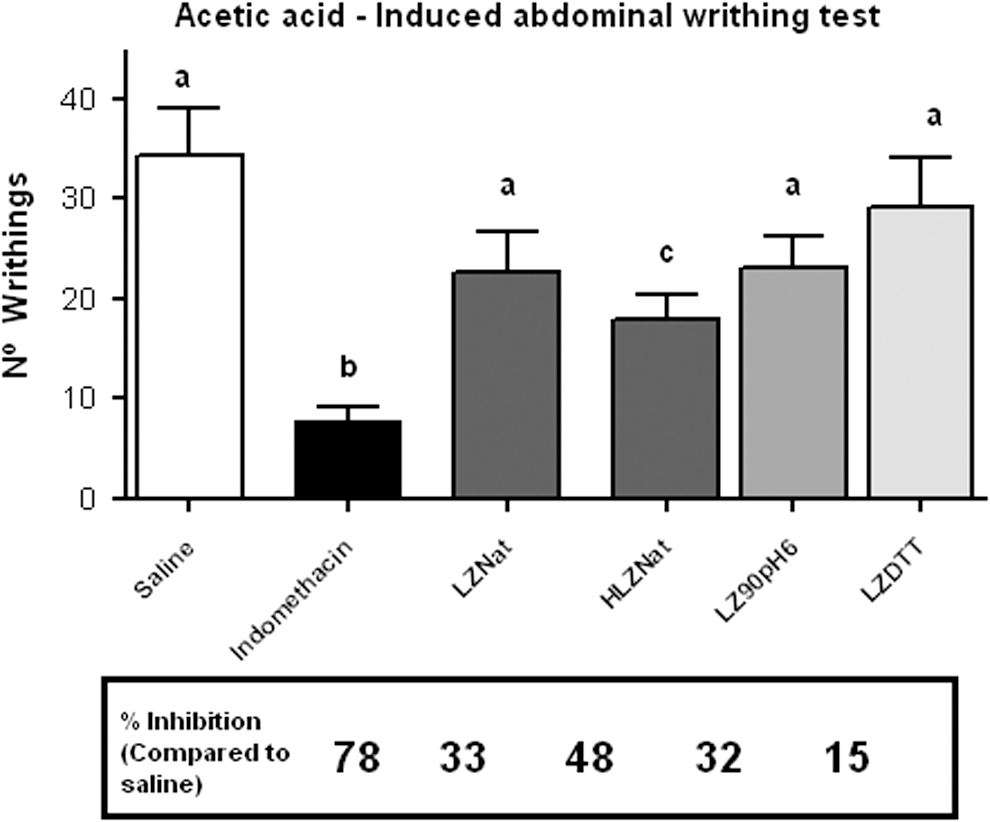
Analgesic effect of the native and modified LZ. Vertical bars represent mean ± SEM. Different letters are statistically different from the control group (P < .05) ANOVA and Tukey's test. LZNat, native LZ; HLZNat, Hydrolyzed native LZ; LZ90pH6, denatured at 90°C, pH6 Lysozyme; LZDTT, denatured LZ with DTT. Data inside the bars show percentage of inhibition.
Conclusions
Heat- and chemically denatured LZ showed strong anti-inflammatory capacity comparable with that of dexamethasone, a potent glucocorticoid. In addition, pepsin-digested LZ exerted remarkable anti-nociceptive activity. Denatured hen egg white LZ had more robust anti-inflammatory and paw edema-reducing properties than natïve enzyme, suggesting that these effects can be independent of their natural enzymatic function in its native form. Therefore, these novel data point out the potential use of denatured and digested LZ as therapeutic agents and offer alternatives to the use of NSAIDs. Further research needs to be done on the therapeutic mechanisms of LZ to optimize its beneficial effects against inflammation and pain.
Footnotes
Acknowledgments
This work was supported by projects, CYTED, AGL2011-24643, and Consolider-Ingenio FUN-C-Food CSD 2007-063, from Ministerio de Economía y Competitividad. The authors are participants in the FA1005 COST Action INFOGEST on food digestion. W. Carrillo acknowledges the Autonomous Community of Madrid for its support through a PhD grant. Special thanks are given to English editor Emilio Labrador.
Author Disclosure Statement
No competing financial interests exist.