
Abstract
Low-grade chronic inflammation (metaflammation) is a major contributing factor for the onset and development of metabolic diseases, such as type 2 diabetes, obesity, and cardiovascular disease. Nicotinamide riboside (NR), which is present in milk and beer, is a functional vitamin B3 having advantageous effects on metabolic regulation. However, the anti-inflammatory capacity of NR is unknown. This study evaluated whether NR modulates hepatic nucleotide binding and oligomerization domain-like receptor family, pyrin domain containing 3 (NLRP3) inflammasome. Male, 8-week-old KK/HlJ mice were allocated to the control or NR group. NR (100 mg/kg/day) or vehicle (phosphate-buffered saline) was administrated by an osmotic pump for 7 days. Glucose control, lipid profiles, NLRP3 inflammasome, and inflammation markers were analyzed, and structural and histological analyses were conducted. NR treatment did not affect body weight gain, food intake, and liver function. Glucose control based on the oral glucose tolerance test and levels of serum insulin and adiponectin was improved by NR treatment. Among tested lipid profiles, NR lowered the total cholesterol concentration in the liver. Histological and structural analysis by hematoxylin and eosin staining and transmission electron microscopy, respectively, showed that NR rescued the disrupted cellular integrity of the mitochondria and nucleus in the livers of obese and diabetic KK mice. In addition, NR treatment significantly improved hepatic proinflammatory markers, including tumor necrosis factor-alpha, interleukin (IL)-6, and IL-1. These ameliorations were accompanied by significant shifts of NLRP3 inflammasome components (NLRP3, ASC, and caspase1). These results demonstrate that NR attenuates hepatic metaflammation by modulating the NLRP3 inflammasome.
Introduction
N
Inflammation is a defense mechanism by which the body recovers from injuries and is accompanied by heat, redness, pain, and swelling. Inflammation is considered a hallmark of obesity, type 2 diabetes, nonalcoholic fatty liver disease (NAFLD), and cardiovascular disease. 8 Metabolic inflammation differs from classic inflammation and is characterized by a low-grade and chronic state. It is termed metaflammation and is considered harmful. 8 Metaflammation affects the whole body or metabolic tissues such as the adipose tissue, muscle, and liver. 9
The inflammatory response is controlled by the coordinated network of various proinflammatory and anti-inflammatory cytokines. Interleukin-1 (IL-1) is a proinflammatory cytokine, and inflammasomes regulate the maturation and secretion of IL-1. 10 Inflammasomes are multiprotein complexes consisting of nucleotide-binding oligomerization domain-like receptor (NLR) family member, adaptor proteins like apoptosis-associated speck-like protein containing a caspase activation and recruitment domain (ASC), and caspase 1. 11,12 Caspase1 activation and IL-1 signaling have been suggested to be involved in the pathogenesis of alcoholic liver disease and NAFLD. 13 The NLRP3 inflammasome is the best characterized inflammasome and this inflammasome activation affects many metabolic disorders. 14 Previous studies have suggested the relationship of the NLR family, pyrin domain containing 3 (NLRP3) and NAFLD. NLRP3 knockin mice present severe liver inflammation, fibrosis, and hepatocyte pyroptosis. 15 In addition, NLRP3 inflammasome-associated proteins are increased in patients with nonalcoholic steatohepatitis (NASH) compared with non-NASH NAFLD patients. 16 These reports indicate that the regulation of the NLRP3 inflammasome may be an effective strategy for delaying the onset and progression of NAFLD.
Recently, it was reported that NR supplementation increases energy expenditure and improves the oxidative metabolism in a high-fat diet-induced obese mouse model. 17 Oral NR treatment for 16 weeks can delay the progression of mitochondrial myopathy in mice by promoting mitochondrial biogenesis and preventing mitochondrial ultrastructure abnormalities. 18 NR administration enhances the motor performance of genetic mitochondrial disease mice by increasing the mitochondrial function. 19 Dietary NR treatment for 12 weeks in an Alzheimer's mouse model can improve cognitive deterioration by increasing NAD+ in the cerebral cortex and peroxisome proliferator-activated receptor gamma coactivator 1-alpha expression in the brain. 20
However, it has not yet been reported whether NR affects systemic and hepatic inflammation as well as the NLRP3 inflammasome. Therefore, we tested the hypothesis that NR treatment improves hepatic metaflammation by modulating the NLRP3 inflammasome in an obese and diabetic rodent model.
Materials and Methods
Animals
Male KK/HlJ mice were provided by the Jackson Laboratory (Bar Harbor, ME, USA) and were fed a standard diet (D12450B; Research Diets, New Brunswick, NJ, USA). Male, 8-week-old KK/HlJ mice were allocated to the control or NR group. NR (100 mg/kg/day; ChromaDex Corporate, Irvine, CA, USA) or vehicle (phosphate-buffered saline) was administrated using an ALZET micro-osmotic pump 1007D (DURECT Corporation, Cupertino, CA, USA) for 7 days. All mice had free access to water and food and were maintained under a controlled temperature of 20°C±5°C with a 12-h light/12-h dark cycle. All procedures were approved by the Animal Care and Use Committee of Chonnam National University.
Sample collection
Mice were fasted for 12 h before sacrifice and anesthetized with Zoletil/Rompun (2:1). Blood was collected by cardiac puncture. After blood collection, the liver, epididymal fat, and subcutaneous fat depots were carefully dissected, weighted, and stored at −80°C until further analysis.
Oral glucose tolerance test
The oral glucose tolerance test (OGTT) was performed on mice before and after treatment. After 16 h of fasting, glucose solutions were given through the stomach gavage (2 g/kg body weight). Blood glucose levels were measured at 0, 15, 30, 60, 90, and 120 min after the glucose challenge using a Glucocard X-Meter (Arkray, Kyoto, Japan).
Biochemical analysis
Serum and liver triglyceride concentrations were determined using kits from Sigma-Aldrich (St Louis, MO, USA). Commercially available ELISA kits were used to measure adiponectin (AdipoGen, Incheon, Korea), insulin, leptin (Crystal Chem, Downers Grove, IL, USA), IL-6, and tumor necrosis factor-alpha (TNF-α; R&D Systems, Minneapolis, MN, USA). The aspartate aminotransferase (AST) activity, alanine aminotransferase (ALT) activity (Biovision, Milpitas, CA, USA), and total cholesterol concentration (Wako Pure Chemical Industries, Richmond, VA, USA) were measured using commercial kits.
Histological analysis
Livers were fixed overnight in 10% (v/v) neutral buffered formalin (Sigma-Aldrich), dehydrated in a graded series of alcohol washes, cleared in toluene, and embedded in paraffin. Using a microtome, 5-μm sections were prepared, collected on slides, and stained with hematoxylin and eosin (H&E).
Transmission electron microscopy analysis
Freshly isolated liver tissues from mice were fixed with 2% glutaraldehyde/2% paraformaldehyde in a 0.1 M cacodylate buffer for 2 h. The samples were processed and thin sections (80 nm) were cut on a Reichert Ultracut E microtome. Sections were viewed at 80 kV with a JEM-1400 TEM (JEOL, Tokyo, Japan). Micrographs were taken (15–20 per sample) by random sampling (magnification: 10,000–25,000×).
Real-time polymerase chain reaction
Total RNA was extracted from the mouse liver using the PureLink RNA Mini kit (Invitrogen, Carlsbad, CA, USA). cDNA was synthesized using the SuperScript III First-Strand Synthesis System (Applied Biosystems, Foster City, CA, USA) following the manufacturer's instructions. mRNA expression levels of various genes were determined using the StepOnePlus real-time PCR system (Life Technologies, Carlsbad, CA, USA), Power SYBR Master Mix (Applied Biosystems), and a gene-specific primer (Bioneer, Daejeon, Korea). Individual reactions for target and 18S were carried out separately with negative controls lacking cDNA. The conditions used were 95°C for 10 min, followed by 40 cycles of denaturation (95°C for 10 sec), annealing (Tm [°C] for 15 sec), and extension (72°C for 60 sec). The cycle number for the threshold of detection was determined by StepOne Software (Life Technologies). mRNA expression of each target was normalized to that of the 18S gene and expressed as the fold change relative to controls.
Statistical analysis
Data are expressed as mean±standard error of the mean. Statistical significance was determined by a Student's t-test. SPSS Statistics 22 (SPSS, Inc., Chicago, IL, USA) was used for statistical tests. P<.05 was considered statistically significant.
Results
Body weight, food intake, and tissue weights
NR did not affect the food intake or body weight (Table 1), and no signs of toxicity were observed during the period of the experiment. In addition, NR had no effect on the food intake (Table 1). The subcutaneous fat (% body weight) in NR mice was 20% lower than in control mice. The weights of epididymal fat and liver tissues were not altered by NR (Table 1).
Data are expressed as mean±SEM (n=9–10 per group).
P<.05 versus control.
BW, body weights; NR, nicotinamide riboside; SEM, standard error of the mean.
Changes of metabolic parameters in NR-treated mice
Activities of ALT and AST were similar in both groups (Table 2), which suggests that NR treatment did not impair liver function. Total cholesterol concentrations in livers were significantly lower with NR administration (Table 2). NR-treated mice had 40% lower hepatic total cholesterol compared with the control (P=.02). Serum total cholesterol and triglyceride concentrations were not significantly different between the control and NR groups (Table 2).
Data are expressed as mean±SEM (n=9–10 per group).
P<.05 versus control.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; TC, total cholesterol; TG, triglyceride.
Effects of NR treatment on glucose control
At baseline, the accumulative area under the curve (AUC) for glucose responses during OGTT was similar in all groups (Fig. 1A, B). One week of NR treatment significantly decreased the accumulative AUC (Fig. 1C, D), suggesting improved glucose control in the rodent model of type 2 diabetes. Serum insulin concentrations were significantly lower in NR-treated mice when compared with the control mice (30% decrease; Fig. 2A). Also, serum adiponectin was significantly higher in the NR group (30% increase; Fig. 2B). One week of NR treatment had no effect on serum leptin levels (Fig. 2C).
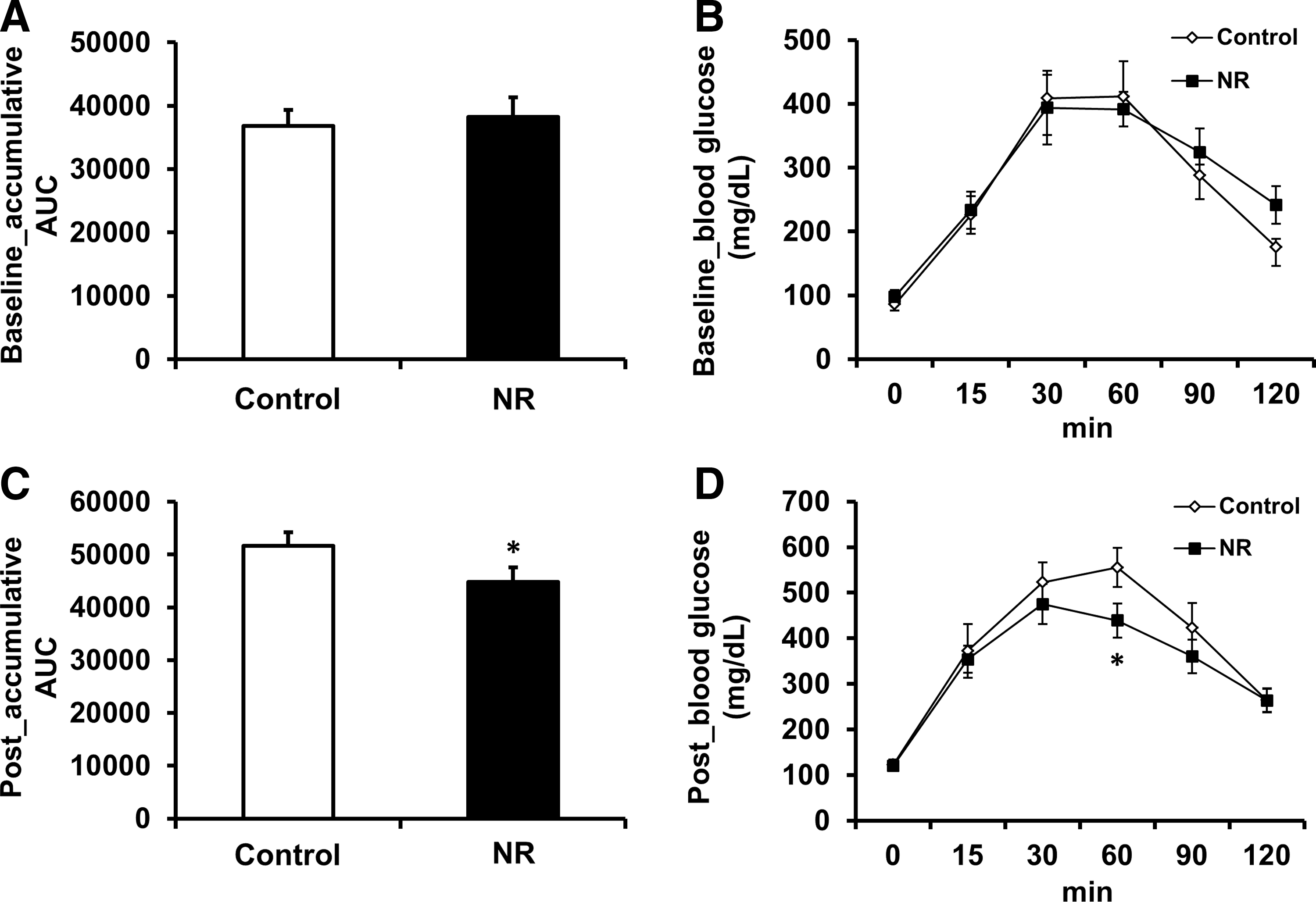
Effect of nicotinamide riboside (NR) on glycemic control during the oral glucose tolerance test (OGTT). At baseline,

Effect of NR treatment on
Effects of NR on hepatic morphology
H&E-stained liver sections demonstrated that the NR-treated mice had healthy appearing hepatocytes compared with the control mice (Fig. 3A). Transmission electron microscopy images showed that NR rescued disrupted cellular integrity of mitochondria in the livers of obese and diabetic KK mice (Fig. 3B). In addition, appearances of cell nuclei in the NR-treated mice were more normal than the control mice.
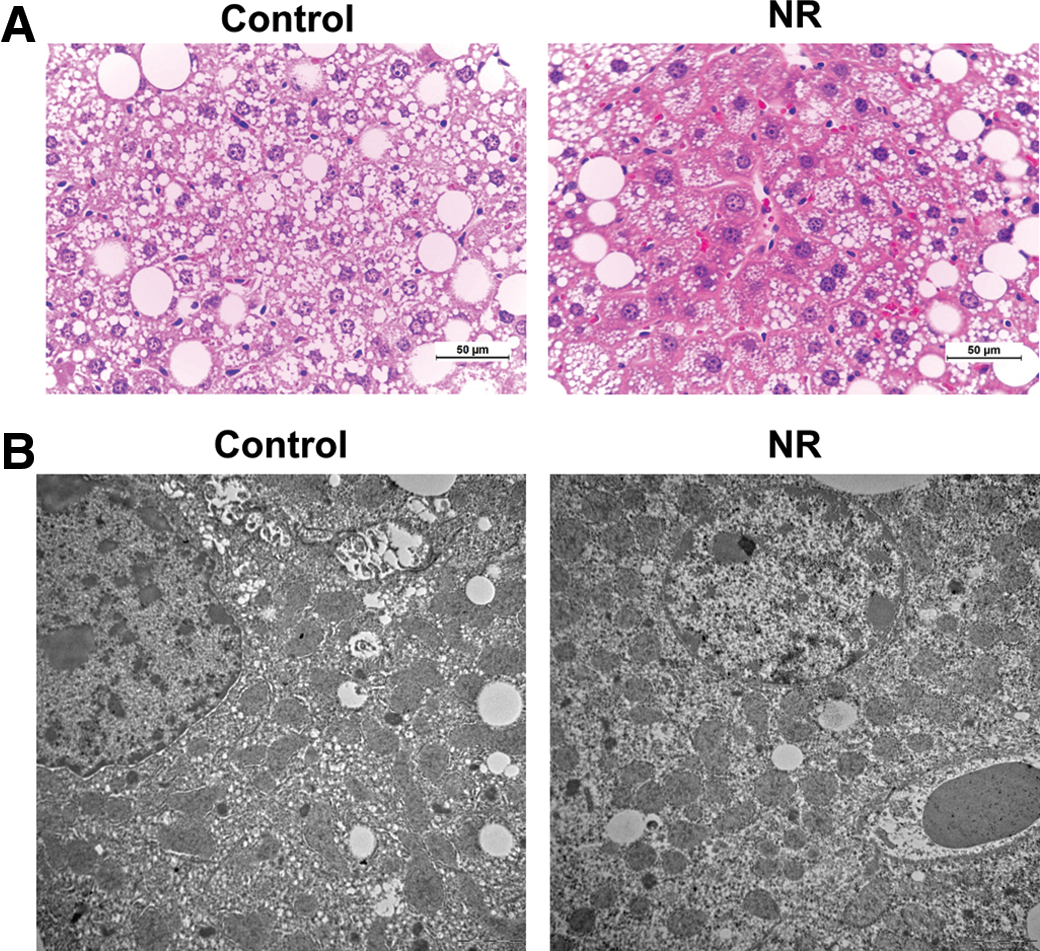
Representative photographs of
NR downregulates NLRP3 inflammasome and ameliorates hepatic metaflammation
The gene expression level of NLRP3 was markedly decreased in NR-treated mouse livers compared with controls (Fig. 4A). Also, mRNA expression levels of ASC and caspase 1, which are NLRP3 inflammasome components, were significantly lower in mice receiving NR treatment (Fig. 4B, C). These alterations lead to lower IL-1 in the NR group (Fig. 4D). The mRNA expressions of the proinflammatory markers TNF-α and IL-6 were significantly decreased in NR-treated mouse livers by 58% and 56%, respectively (Fig. 5A, B). These gene expression data were confirmed by the protein analysis results by ELISA (Fig. 5C–E). The findings suggest that NR treatment ameliorates hepatic metaflammation.

Effect of NR on nucleotide binding and oligomerization domain-like receptor family, pyrin domain containing 3 (NLRP3) inflammasome in the liver. mRNA expression of

Effect of NR on inflammatory markers. mRNA expression of
Discussion
We investigated whether NR ameliorates hepatic metaflammation and whether this improvement might be causally linked to the altered expression and/or activation of the NLRP3 inflammasome in vivo. NR treatment (100 mg/kg body weight/day) did not cause any negative effects to the mice. Nicotinic acid at pharmacological doses is often limited because of the side effects that include hepatic toxicity. 21 Hepatic total cholesterol levels were lower in NR-treated mice, even though serum total cholesterol levels were not affected. Nicotinic acid has been used for the treatment of dyslipidemia and cardiovascular diseases. 22,23 The liver is a major organ for the production and secretion of apo B, its associated lipids, very low-density lipoprotein, and low-density lipoprotein particles. Nicotinic acid influences plasma triglyceride and the secretion of apo B-containing lipoproteins in the liver. 24 In HepG2 cells, nicotinic acid inhibits microsomal diacylglycerol acyltransferase (DGAT), which catalyzes the formation of triglyceride and is a key rate-limiting enzyme in triglyceride synthesis. 25 DGAT2 knockout mice have lower triglyceride levels, 26 suggesting that nicotinic acid decreases triglyceride synthesis by inhibiting the expression and activities of DGAT and that the reduced availability of DGAT in hepatocytes increases apo B degradation and subsequently decreases the secretion of very low-density lipoprotein and low-density lipoprotein particles. Altered expression and/or activity of DGAT are suggested as the underlying mechanisms for the amelioration of cholesterol profiles by NR. However, this improvement did not continue to triglyceride levels, which may be due to the short-term treatment period.
In this study, NR ameliorated glucose control in obese and diabetic mice, accompanied by increased adiponectin and decreased insulin levels. Similar effects were observed in a high-fat diet-induced obesity mouse model. 17 Adiponectin is an adipokine with insulin-sensitizing and anti-inflammatory properties. 27,28 Decreased adiponectin levels have been documented in obese, insulin-resistant, and type 2 diabetes patients. 29,30 The increase in adiponectin levels is one of the therapeutic targets for insulin-sensitizing and antidiabetic drugs given to insulin-resistant individuals. 31 A meta-analysis report suggested that higher adiponectin levels are associated with a lower risk of developing type 2 diabetes across diverse populations. 32 In mice, adiponectin administration reverses insulin resistance and weight gain. 33
NR reduced the expression levels of the NLRP3 inflammasome complex, including NLRP3, ASC, and caspase1, and leads to the lower IL-1 levels. These alterations ameliorated hepatic metaflammation by changing the levels of the proinflammatory cytokines, IL-6 and TNF-α. Inflammasomes are involved in the regulation of innate immunity responses. Danger signals, including pathogen-associated molecular patterns, danger-associated molecular patterns, and environmental irritants, activate the NLRP3 inflammasome. 34,35 The interaction of the pyrin domain of ASC with the pyrin domain of NLRP3 induces the assembly of the NLRP3 inflammasome. The interaction of a dual caspase activation and recruitment domain of ASC with pro-caspase1 forms a functional inflammasome complex, and the cleavage with caspase1 leads to the maturation and secretion of the proinflammatory cytokine IL-1β. 10 Recently, the inflammasome function has been implicated in many common conditions, such as type 2 diabetes, obesity, and atherosclerosis. 36 –38 In the present study, 1 week of NR treatment resulted in lower levels of NLRP3, ASC, and caspase1. The NR-mediated inhibition of the NLRP3 inflammasome activation suggests that NR ameliorates glucose control and decreases hepatic cholesterol, in part, by the modulation of the NLRP3 inflammasome and the maturation of IL-1β. In addition, NR may improve hepatic metaflammation by suppressing IL-6 and TNF-α secretion. Moreover, inflammasome activation promotes T-cell responses. 39 Recent studies in rodent and human suggest that the NLRP3 inflammasome and T-helper 1 cell shift in the T-cell population in adipose tissue contribute to inflammation and insulin resistance. 40 The effect of NR-mediated inhibition of NLRP3 inflammasome activation on T cell and other inflammatory factors will be interesting to study.
This study has several limitations. First, 1 week of NR treatment may not have been sufficient to demonstrate significant changes of the levels of serum glucose and triglyceride in the liver and serum. Second, although no side effects of NR were observed in this study, the safety of the long-term treatment with NR should be considered. Further investigations are needed to clarify the detailed mechanisms underlying the modulation of the NLRP3 inflammasome by NR and the effects of long-term NR treatment on metaflammation and its related diseases. The present study supports the hypothesis that NR treatment improves hepatic metaflammation by modulating NLRP3 inflammasome in a mouse model of obesity and type 2 diabetes. These findings demonstrate that NR has potential hepatic anti-inflammatory activity and suggest the possibility of NR as a therapeutic agent against hepatic metaflammation and its related diseases.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2014R1A1A2A16055328) and the Korean Diabetes Association (S.J.Y., 2013). The funder had no role in the study design, data collection, analysis and interpretation, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.