
Abstract
Stamens of Nelumbo nucifera Gaertn have been used as a Chinese medicine due to its antioxidant, hypoglycemic, and antiatherogenic activity. However, the effects of kaempferol, a main component of N. nucifera, on obesity are not fully understood. We examined the effect of kaempferol on adipogenesis and fatty acid oxidation signaling pathways in 3T3-L1 adipocytes. Kaempferol reduced cytoplasmic triglyceride (TG) accumulation in dose and time-dependent manners during adipocyte differentiation. Accumulation of TG was rapidly reversed by retrieving kaempferol treatment. Kaempferol broadly decreased mRNA or protein levels of adipogenic transcription factors and their target genes related to lipid accumulation. Kaempferol also suppressed glucose uptake and glucose transporter GLUT4 mRNA expression in adipocytes. Furthermore, protein docking simulation suggests that Kaempferol can directly bind to and activate peroxisome proliferator-activated receptor (PPAR)-α by forming hydrophobic interactions with VAL324, THR279, and LEU321 residues of PPARα. The binding affinity was higher than a well-known PPARα agonist fenofibrate. Consistently, mRNA expression levels of PPARα target genes were increased. Our study indicates while kaempferol inhibits lipogenic transcription factors and lipid accumulation, it may bind to PPARα and stimulate fatty acid oxidation signaling in adipocytes.
Introduction
O
Nelumbo nucifera Gaertn. (Nymphaeaceae), commonly called lotus, is widely distributed throughout Eastern Asia, and it has a long history of use as food and Chinese medicine. Among 33 compounds isolated from N. nucifera leaves, liriodenine and pronuciferine showed antiobesity effect by inhibiting pancreatic lipase activity and adipocyte differentiation. 9 Also, N. nucifera leaf flavonoids may ameliorate hyperlipidemia and type II diabetes mellitus by inhibiting porcine pancreatic lipase, α-amylase, and α-glucosidase. 10,11 However, it is still not fully understood how kaempferol as a main component of N. nucifera regulates adipocyte metabolism. Our study examined effects of kaempferol from N. nucifera on signaling pathways of adipogenesis, glucose uptake, and fatty acid oxidation in adipocytes.
Materials and Methods
Materials
Kaempferol was isolated from the methanolic extract of N. nucifera stamens described in Lim et al. 12 Dulbecco's modified Eagle's medium (DMEM), penicillin-streptomycin mixture, 0.25% tripsin-ethylenediaminetetraacetic acid (EDTA), TRIzol reagent, calf serum, and fetal bovine serum (FBS) were purchased from Gibco BRL (Gaithersburg, MD, USA). Dexamethasone, 3-isobutyl-1-methylxanthine, insulin, GPO-trinder, and troglitazone were purchased from Sigma Chemical (St. Louis, MO, USA). CellTiter 96 AQueous Nonradioactive Cell Proliferation Assay kit was purchased from Promega (Madison, WI, USA). Polyclonal antibodies against PPARγ, C/EBPα, SCD-1, FAS, and SREBP-1c were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Protein standard marker was purchased from Amersharm Pharmacia (Piscataway, NJ, USA) and the enhanced chemiluminescence (ECL) detection kit was purchased from Perkin Elmer Life Science (Waltham, MA, USA).
Cell culture and treatments
Murine 3T3-L1 preadipocytes (ATCC, Rockville, MD, USA) were cultured in DMEM with 4.5 g/L glucose, 10% calf serum, penicillin/streptomycin (100 U/mL penicillin and 100 μg/mL streptomycin in 0.85% saline), and 100 μM sodium pyruvate. Two days postconfluence, 3T3-L1 preadipocytes were differentiated into adipocytes by DMEM containing 10% FBS, 100 μM 3-isobutyl-1-methylxanthine, 250 nM dexamethasone, and 170 nM insulin (MDI media). On day 3, the medium was replaced with DMEM containing 10% FBS and 170 nM insulin (INS media). The medium was changed every 2 days until performing experiments. Cells were cultured in the absence (dimethyl sulfoxide [DMSO] alone) or in the presence of kaempferol (dissolved in DMSO) at concentrations and periods indicated in the figure legends. During the period of kaempferol exposure, the medium was changed every 2 days for both controls and treated cells.
Cell proliferation assay
Seventy to eighty percent confluent 3T3-L1 preadipocytes were seeded in 96-well plates at a density of 10,000 cells/well. Vehicle or kaempferol (0–40 μM) was added to culture medium and incubated for 24 h. Colorimetric proliferation assay (CellTiter 96 AQueous Nonradioactive Cell Proliferation Assay kit; Promega) was performed based on the manufacturer's manual.
Triglyceride assay
Cells were washed twice with phosphate-buffered saline (PBS) and collected by scraping with a cell scraper into 50 mM sodium phosphate buffer (pH 7.4) containing 2 mM EDTA and 2 M NaCl. The harvested cells were sonicated two times for 5 s at 40 W with the microtip of a Branson model 250 Sonifier (Branson Ultrasonics, Danbury, CT, USA). After centrifugation at 19,300 g for 10 min at 4°C, triglyceride (TG) content was analyzed using a commercial kit (GPO-trinder; Sigma Chemical) according to the manufacturer's instructions.
RNA isolation and real-time PCR
Total RNA was isolated from 3T3-L1 cells using TRIzol reagent (Life Technologies, Carlsbad, CA, USA) with DNase I according to the manufacturer's instructions. Reverse transcription and polymerase chain reaction (PCR) were performed with QuantiTect SYBR Green PCR kit according to the manufacturer's instructions (Qiagen, Inc., Valencia, MD, USA) on an ABI Prism 7900 Sequence Detection System (Life Technologies). Post-PCR melting curves confirmed the specificity of single-target amplification. The quantification of each mRNA was normalized by the expression levels of β-actin, and all experiments were repeated five to seven times. Primers used in this study were as follows: PPARγ, 5′-CCCTGGCAAAGCATTTGTAT-3′ (forward) and 5′-GAAACTGGCACCCTTGAA AA-3′(reverse); C/EBPα, 5′-TTACAACAGGCCAGGTTTCC-3′(forward) and 5′-AA CTCCAGTCCCTCTGGGAT–3′(reverse); liver X receptor-α (LXRα), 5′-TCTGCAATTGAGGTCATGCT (forward) and 5′-TGCAGAGAAGATGCTGATGG-3 (reverse); SCD-1, 5′-GAACTT ACAAGGCTCGGCTG-3′ (forward) and 5′-AGACATGTCCAGTTTTCCGC-3′ (reverse); GLUT4, 5′-ATCTTGATGACCGTGGCTCT-3′ (forward) and 5′-TAGC ATCCGCAACATACTGG-3′(reverse); FAS, 5′-GCTGTGCTTGCAGCT TACTG-3′ (forward) and 5′-TGTCCTCAGAGT TGTGGCAG-3′ (reverse); SREBP-1c, 5′–TCATGCCCTCCATAGACACA-3′ (forward) and 5′-AGCTCAAAGACCTGGTGGTG-3′ (reverse); RXRα, 5′-CCTAGGTGGCTTGATGTGGT-3′ (forward) and 5′-AGCCATTGTCCTGTTCAACC-3′ (reverse); AP2, 5′-TGCTCGAGTTTGGTC-TGAAC-3′ (forward) and 5′-CTCTGATCAGTCGCCTCCTC-3′ (reverse); and β-actin, 5′-CTGACTGACTA-CCTCATGAAGATCCT-3′ (forward) and 5′-CTTAATGTCACG CACGATTTCC-3′ (reverse).
Immunoblot analysis
Protein samples from 3T3-L1 cells were prepared using a lysis buffer (50 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1% Nonidet-40, 1% Tween-20, 0.1% SDS, 1 mM Na3VO4, 10 μg/mL leupeptin, 50 mM NaF, and s1 mM phenylmethylsulfonyl fluoride). Total proteins (40 μg) were separated by SDS-PAGE and transferred into a nitrocellulose membrane. The membrane was blocked with 5% nonfat dried milk in Tris-buffered saline with Tween-20 (TBST; 10 mM Tris–HCl, pH 8.0, 150 mM NaCl, and 0.06% Tween-20) for 2 h and incubated for 1 h with primary antibody in TBST containing 5% nonfat dried milk. The blots were treated with horseradish peroxidase-conjugated secondary antibody in TBST containing 5% nonfat dried milk for 1 h and detected using an ECL detection kit.
Glucose uptake
Glucose uptake was analyzed by measuring the uptake of 2-deoxy-
Docking simulation of kaempferol and PPARα
Because of automated docking capability, Dock6 was used for the in silico protein–ligand docking simulation. The three-dimensional structure of PPARα was used in the crystal structure of Homo sapiens (PDB ID: 1K7L). As a docking pocket, predefined binding site of PPARα was used. Docking simulations were performed between PPARα and kaempferol or fenofibrate, a well-known PPARα agonist. To prepare compounds for docking simulation, (1) two-dimensional structures were converted into three-dimensional structures, (2) charges were calculated, and (3) hydrogen atoms were added using the ChemOffice program (
Statistical analysis
All measurements were performed independently and repeated at least three times. Data were expressed as the mean ± SD. Data were analyzed using the analysis of variance procedure of the Statistical Analysis System (SAS Institute, Cary, NC, USA). Significant differences among groups were determined using Duncan's multiple range tests. Differences were considered significant if P < .05.
Results
Effect of kaempferol on TG accumulation during adipocyte differentiation
We examined whether the kaempferol induces cytotoxicity in 3T3-L1 cells. The concentrations of kaempferol used in this study did not show cytotoxicity up to 40 μM (Fig. 1). To investigate the effects of kaempferol on cellular TG accumulation, 3T3-L1 cells were treated with various concentrations of kaempferol for 7 days. Kaempferol treatment significantly reduced cellular TG content in a concentration-dependent manner (2.5–40 μM) (Fig. 2). To determine the time-dependent effect of kaempferol on TG accumulation, 3T3-L1 cells were differentiated in the absence or presence of 5 and 40 μM of kaempferol for different time periods. Cellular TG content was not changed until 3 days of the treatment. However, TG content was significantly reduced from day 5 with both 5 and 40 μM of kaempferol treatment and the suppressive effects were increased in a time-dependent manner (Fig. 3). These data indicate that kaempferol is a strong compound that inhibits lipid accumulation in adipocytes.
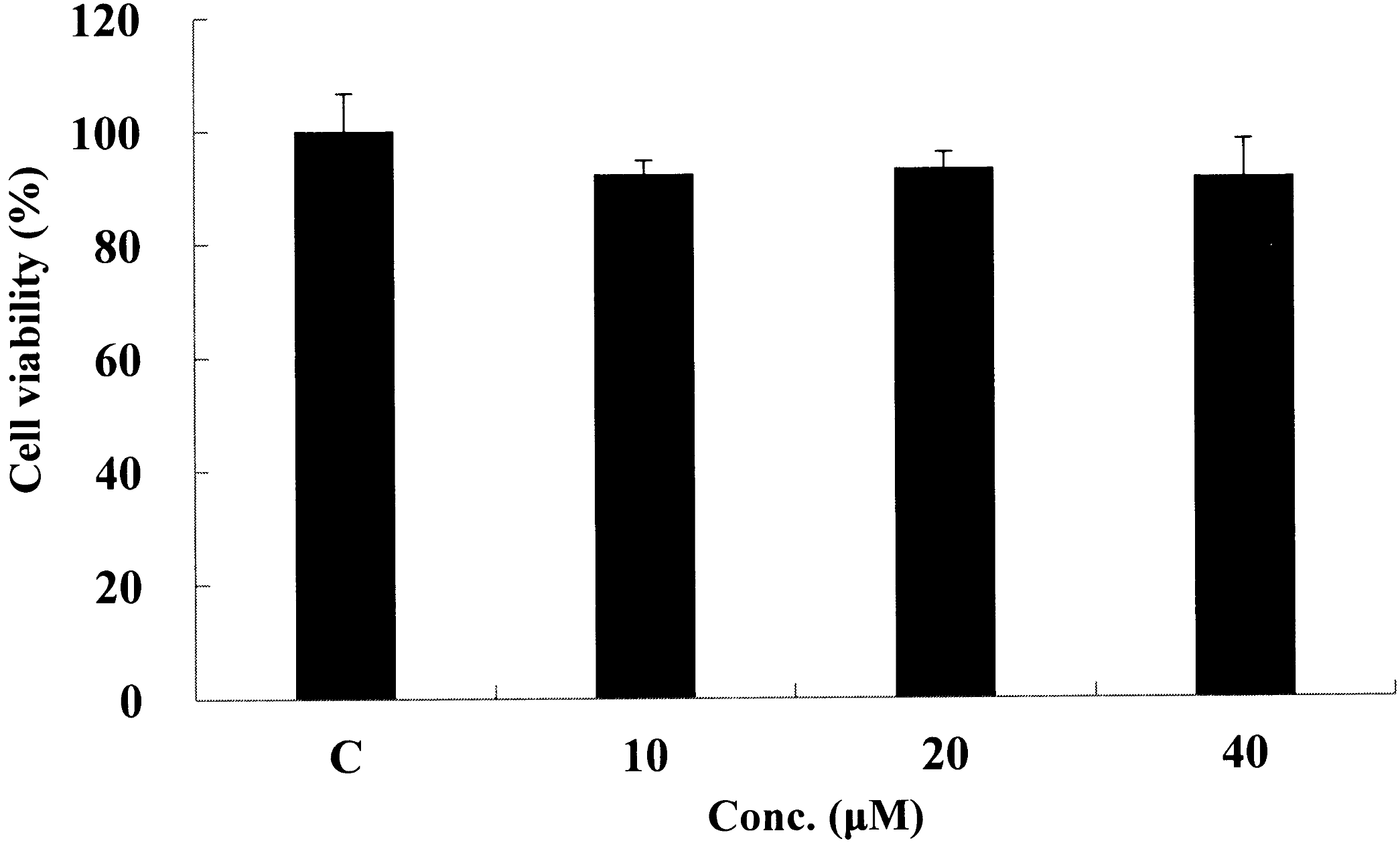
Effect of kaempferol on cytotoxicity of 3T3-L1 cells. Seventy to eighty percent confluent 3T3-L1 cells were treated with kaempferol (10–40 μM) for 24 h. Cell viability was determined by MTT assay.
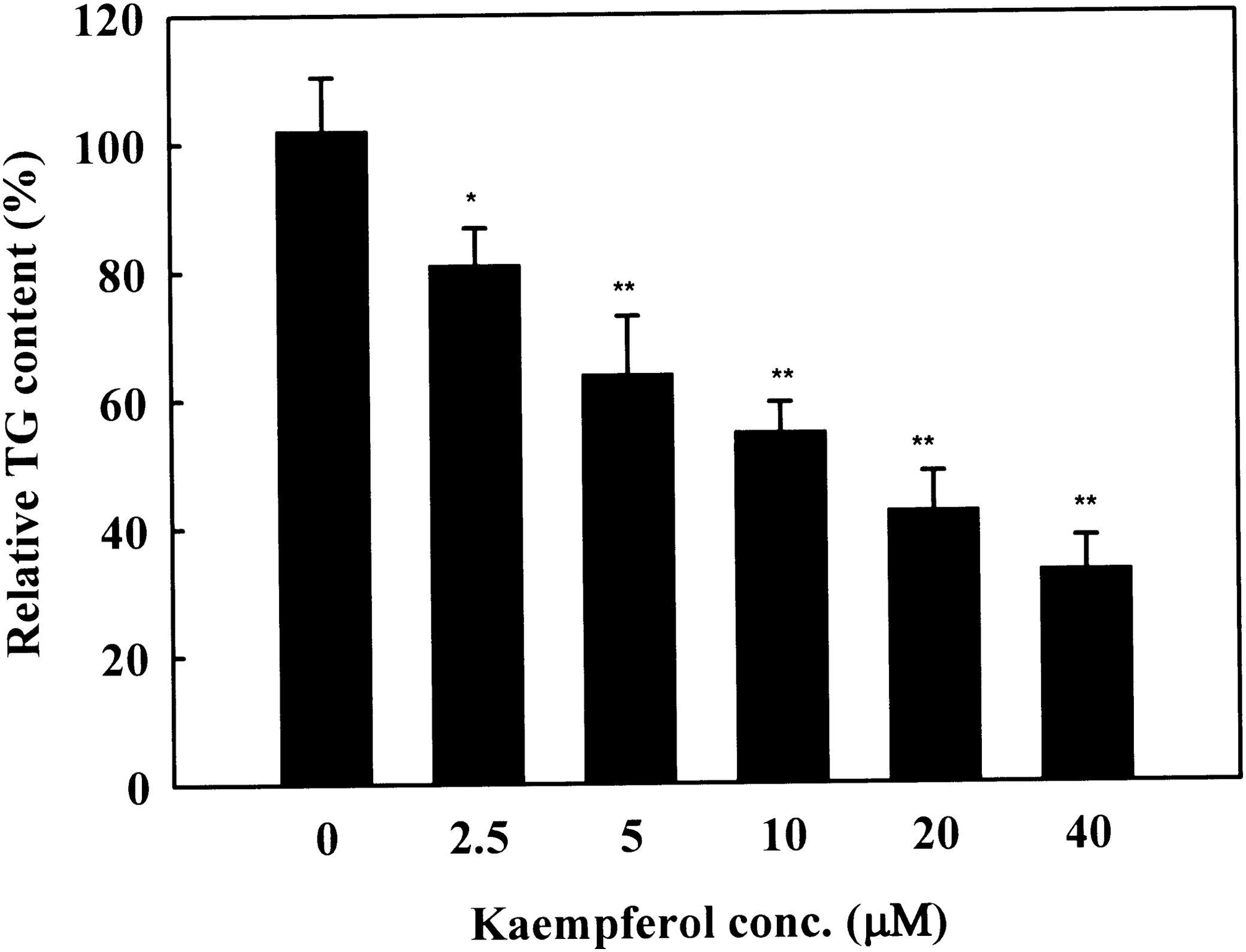
Effect of kaempferol on triglyceride (TG) accumulation during adipogenesis of 3T3-L1. Two-day postconfluence, 3T3-L1 preadipocytes (day 0) were stimulated for 3 days with MDI medium containing kaempferol. On day 3, the MDI medium was replaced with INS medium containing kaempferol. Medium was changed every 2 days and the cells were harvested on day 9 for TG analysis. Results represent the mean ± SD. *P < .05 and **P < .01 indicate significant differences from control cells.
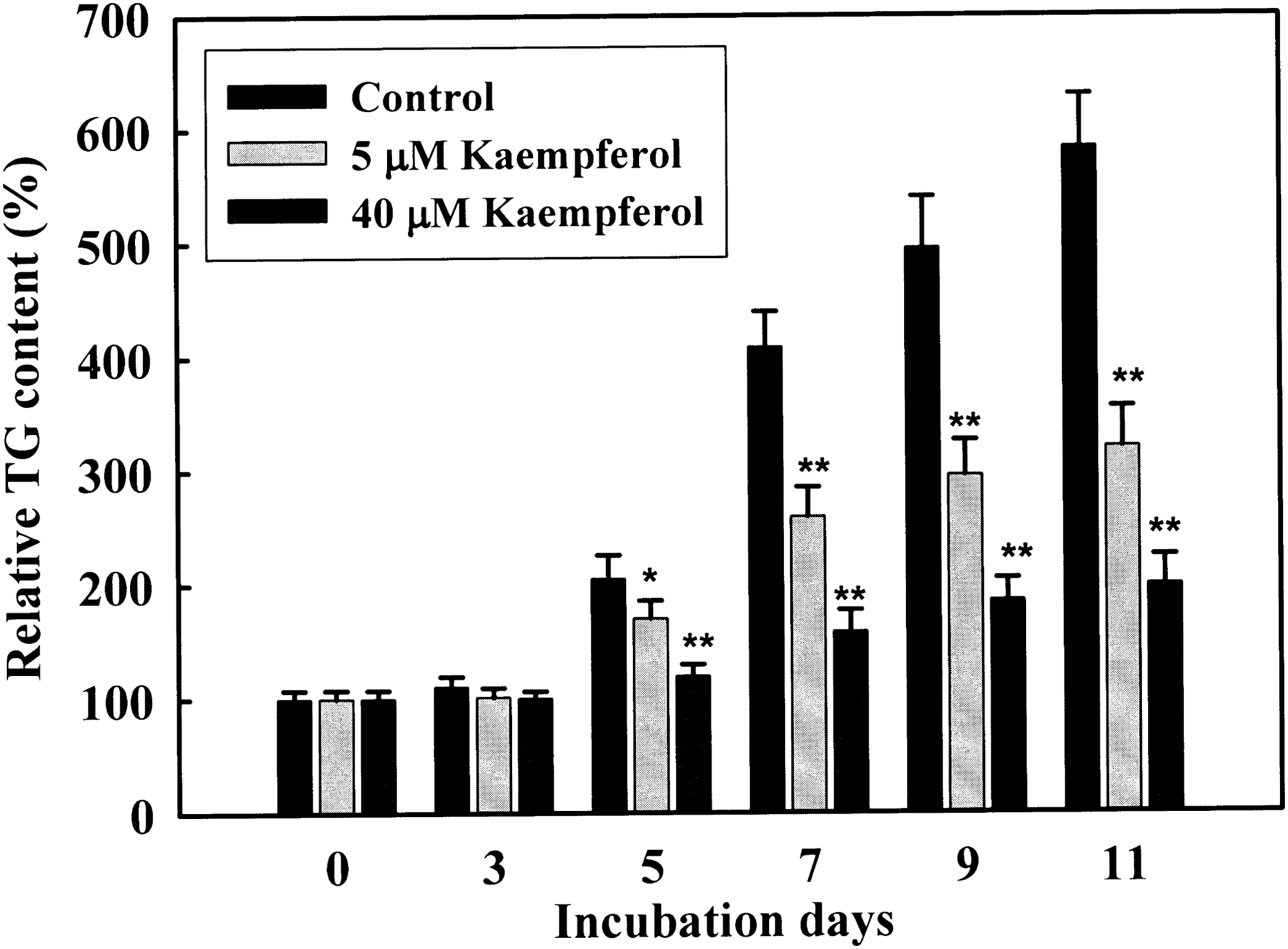
Time-dependent effect of kaempferol on TG accumulation during 3T3-L1 adipogenesis. Two-day postconfluence, 3T3-L1 preadipocytes (day 0) were stimulated with MDI medium containing 5 and 40 μM kaempferol for 3 days. On day 3, MDI medium was replaced with INS medium containing 5 and 40 μM kaempferol. Medium was changed every 2 days. Cells were harvested at day 0, 3, 5, 7, 9 and 11 for TG analysis. Results represent the mean ± SD. *P < .05 and **P < .01 indicate significance differences from control cells.
To determine whether kaempferol-mediated inhibition of TG accumulation was reversible, 3T3-L1 cells were initially cultured from day 0 to day 7 in the absence (control) or the presence of 5 μM kaempferol, approximating the circulating concentration of polyphenols attained after a polyphenol-rich diet. 14 A kaempferol-treated group was divided into two groups in which the cells were further cultured with or without kaempferol to examine whether TG accumulation is reversible. When kaempferol was treated continuously until day 13, cellular TG content was markedly reduced for the entire period of the study (P < .01, Fig. 4). However, the withdrawal of kaempferol restored TG accumulation to levels comparable to the control group by day 13 (Fig. 4), indicating that kaempferol-induced TG reduction was spontaneously reversible. This result suggests that kaempferol regulates TG accumulation in a direct and rapid manner.
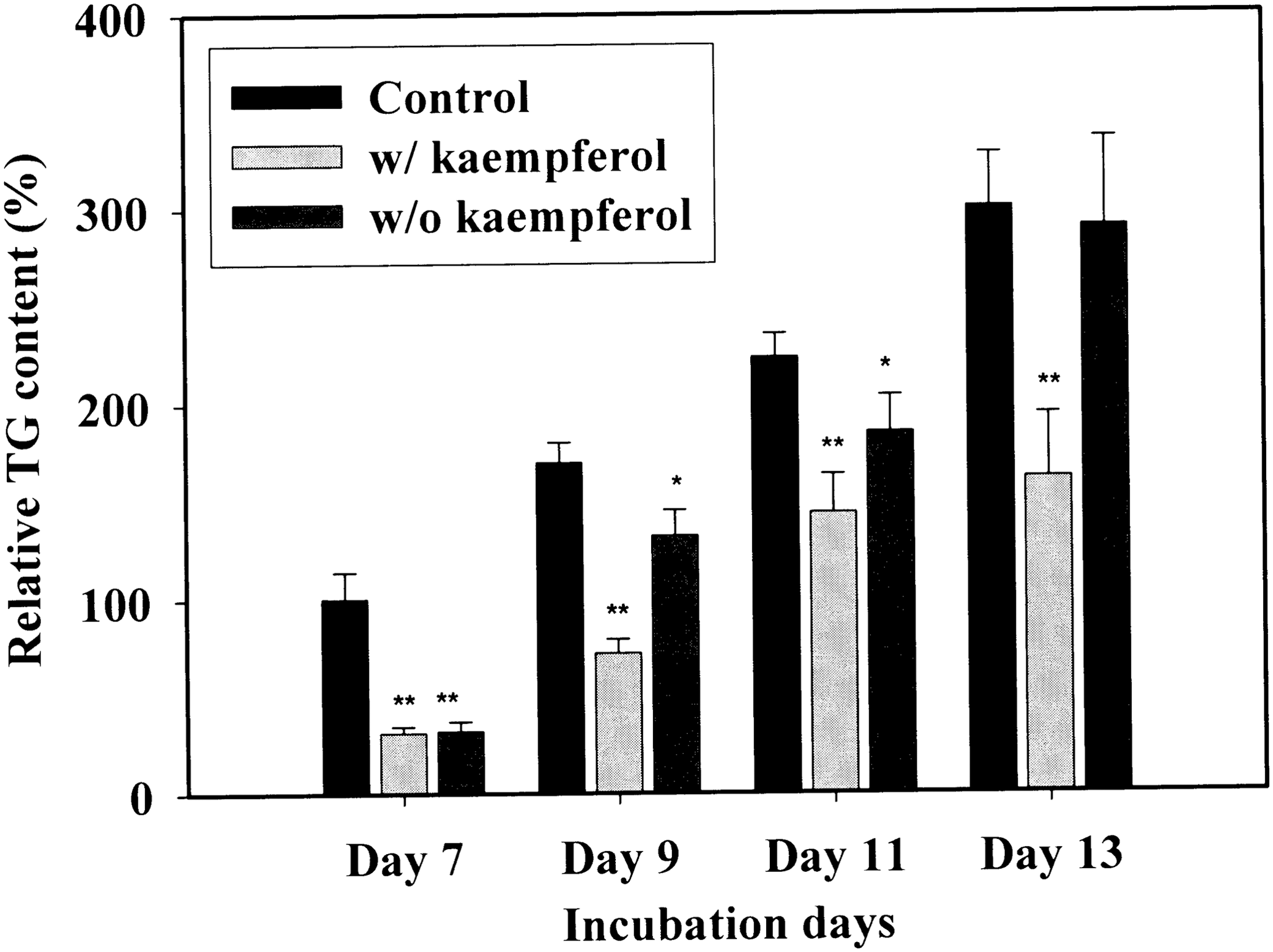
Spontaneous reversibility of TG accumulation in 3T3-L1 cells after kaempferol withdrawal. Two-day postconfluence, 3T3-L1 preadipocytes (day 0) were stimulated with MDI medium containing kaempferol (10 μM) for 3 days. On day 3, the MDI medium was replaced with Dulbecco's modified Eagle's medium (DMEM)/10% fetal bovine serum (FBS) culture medium containing 10 μg/mL insulin. Medium was changed at differentiation day 3 and 5. On day 7, control cells were maintained without kaempferol for an additional 6 days; cells exposed to kaempferol were divided in two groups: kaempferol-treated (10 μM) and kaempferol-withdrawal groups. Cell extracts were prepared on the indicated days and tested for TG content. Results represent the mean ± SD. *P < .05 and **P < .01 indicate significant differences from control cells.
Effect of kaempferol on TG accumulation in mature 3T3-L1 adipocytes
To examine whether kaempferol inhibits lipid accumulation via suppressing lipolysis, 3T3-L1 cells were differentiated for 7 days by MDI treatment without kaempferol. On day 7, mature adipocytes were divided into three groups, which included an untreated group through all culture periods, a treated group with 5 μM kaempferol over the next 6 days, and a treated group with 0.5 μM troglitazone, a PPARγ agonist, over the next 6 days. TG content of cells was measured at differentiation day 7, 9, 11, and 13. The PPARγ agonist increased lipid accumulation even in mature adipocytes (Fig. 5). However, kaempferol did not show any inhibitory effect on TG accumulation in mature adipocytes (Fig. 5). This result indicates that the suppression of TG accumulation by kaempferol may be through suppressing adipocyte differentiation process rather than inhibiting lipid breakdown.

Effect of kaempferol on TG accumulation in mature 3T3-L1 adipocytes. 3T3-L1 cells (day 0) were stimulated with MDI medium for 3 days. On differentiation day 3, the MDI medium was replaced with DMEM/10% FBS culture medium containing 10 μg/mL insulin. At differentiation day 7, the cells were divided into three groups, which include a control group, kaempferol-treated group (10 μM) from differentiation day 7, and troglitazone-treated group (0.5 μM) through all period. TG content of cells was analyzed at indicated time points. Results represent the mean ± SD. *P < .05 and **P < .01 indicate significant differences from control cells.
Effects of kaempferol on adipogenic transcription factors
To explore the mechanism of the kaempferol-driven lipid reduction, we measured mRNA and protein expression levels of adipogenic transcription factors. 3T3-L1 cells were treated with various concentrations of kaempferol for 7 days and mRNA expression levels of adipogenic transcription factors C/EBPα, PPARγ, and RXRα were analyzed by real-time PCR. During adipocyte differentiation, the mRNA expression levels of C/EBPα, PPARγ, RXRα, LXRα, and SREBP-1c were suppressed in a dose-dependent manner up to 10 μM (P < .01, Fig. 6a). However, no further inhibitory effect was found in higher concentrations (20–40 μM) (Fig. 6a). Consistently, the protein level of C/EBPα, PPARγ, RXRα, LXRα, and SREBP-1c (68 kDa active form) was markedly reduced by kaempferol treatment (Fig. 6b). These data suggest that kaempferol broadly inhibit adipogenic transcription factors and thus contribute to the inhibition of TG accumulation in adipocytes.
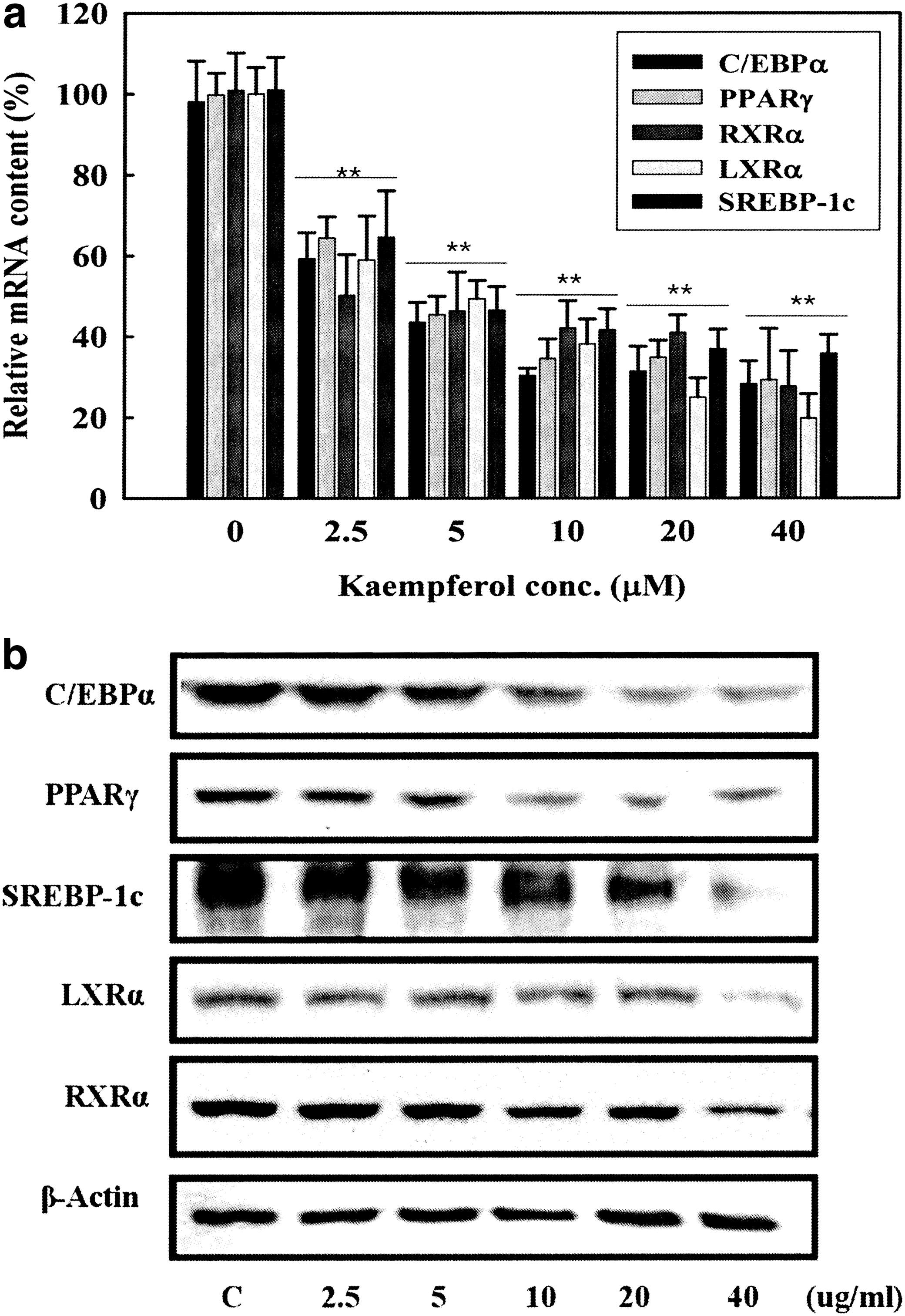
Dose-dependent effect of kaempferol on mRNA and protein expression levels of adipogenic transcription factors. Two-day postconfluence, 3T3-L1 cells (day 0) were stimulated with MDI medium containing kaempferol for 3 days. On differentiation day 3 and 5, the medium was replaced with DMEM/10% FBS culture medium containing 10 μg/mL insulin and indicated concentrations of kaempferol. On day 7,
Effects of kaempferol on enzymes related to lipid accumulation
To ensure functional consequence of kaempferol-driven inhibitory effect on adipogenic transcription factors, we analyzed the effect of kaempferol on mRNA and protein expression levels of their target enzymes including SCD-1, FAS, and ACCα related to de novo fatty acid synthesis. Kaempferol treatment reduced mRNA levels of SCD-1, FAS, and ACCα in a dose-dependent manner (Fig. 7a). In parallel with this, the protein levels of SCD-1, FAS, and ACCα were reduced by kaempferol treatment (Fig. 7b). These data confirmed the inhibitory effect of kaempferol on adipogenic pathway(s) during adipocyte differentiation.
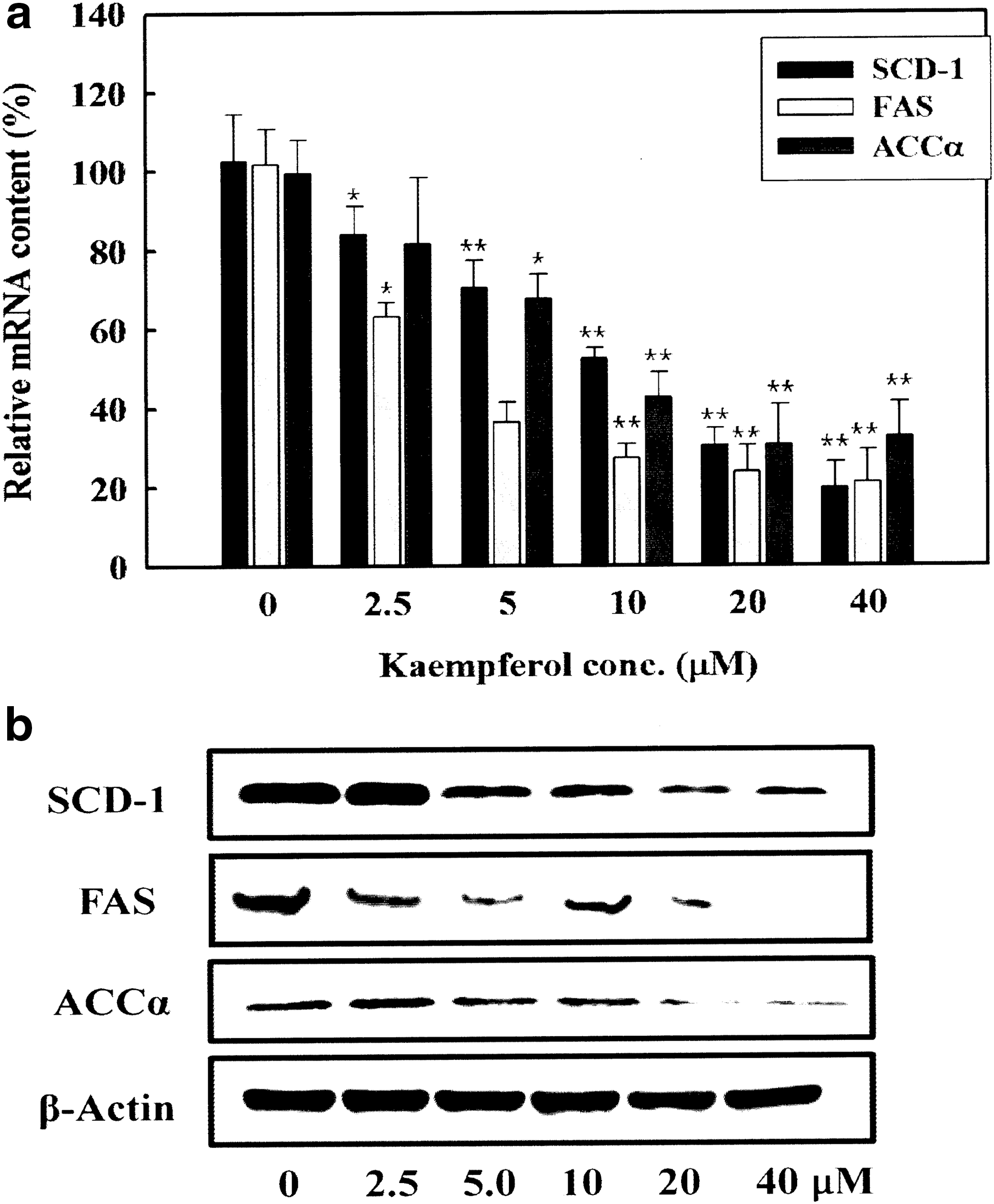
Dose-dependent effect of kaempferol on mRNA and protein expression levels of genes related to lipid accumulation. Two-day postconfluence, 3T3-L1 cells (day 0) were stimulated with MDI medium containing kaempferol for 3 days. On differentiation day 3 and 5, the medium was replaced with DMEM/10% FBS culture medium containing 10 μg/mL insulin and indicated concentrations of kaempferol. On day 7,
Effects of kaempferol on glucose uptake
In response to insulin, glucose uptake is an important feature of adipocytes to accumulate TG. Glucose uptake is primarily mediated by glucose transporter GLUT4. To examine whether the kaempferol plays a role in regulating glucose uptake in adipocytes, GLUT4 mRNA expression was measured. Kaempferol notably inhibited GLUT4 mRNA expression levels (Fig. 8a). To examine whether the reduction of GLUT4 mRNA expression by kaempferol inhibits glucose uptake in adipocytes 2-deoxy-

Dose-dependent effect of kaempferol on GLUT4 mRNA expression and glucose uptake. Two-day postconfluence, 3T3-L1 preadipocytes (day 0) were stimulated with MDI medium containing kaempferol for 3 days. On differentiation day 3 and 5, the medium was replaced with DMEM/10% FBS culture medium containing 10 μg/mL insulin and indicated concentrations of kaempferol. On day 7,
Effects of kaempferol on PPARα and its target genes
It has been reported in an animal study that kaempferol produced effects similar to fenofibrate, a well-known PPARα agonist, such as reducing fat mass, ameliorating blood lipid profile, and so on. 15 The study showed that kaempferol decreased lipogenic transcription factors and increased PPARα target gene expression in liver of high fat-fed rats. To examine whether kaempferol is able to activate PPARα in adipocytes, we applied protein docking simulation to measure binding affinity between kaempferol or fenofibrate and PPARα. Interestingly, kaempferol can directly bind to PPARα by forming hydrophobic interactions with VAL324, THR279, and LEU321 residues of PPARα (Fig. 9a, b). The binding affinity of kaempferol with PPARα (−33) was slightly higher than that of fenofibrate (−30.8) (Fig. 9b), indicating that kaempferol may directly regulate PPARα activity in a similar manner as fenofibrate. To confirm this, we measured mRNA expression levels of PPARα target genes related to fatty acid oxidation including CPT1 and ACO. Kaempferol increased mRNA expression levels of CPT1 and ACO in a dose-dependent manner (Fig. 9c, d), suggesting that kaempferol may activate PPARα-mediated fatty acid oxidation signaling.

Binding capacity of kaempferol on PPARα. Computational structure prediction for docking simulation between PPARα and kaempferol.
Discussion
In this study, we found that kaempferol directly suppressed glucose uptake and lipid accumulation in adipocytes at least partially by inhibiting key transcription factors related to adipocyte differentiation. Our study showed that intracellular lipid accumulation in 3T3-L1 cells was significantly decreased in dose and time-dependent manners (Fig. 2). The low concentration (5 μM) comparable to the concentration achieved by a polyphenol-rich meal 14 was enough for preventing lipid accumulation during adipocyte differentiation. Furthermore, our study showed that kaempferol may directly bind to and activate PPARα and increase its target gene expression. Therefore, kaempferol may be a novel molecule for obesity intervention.
Various transcription factors are associated with adipocyte differentiation and lipid accumulation. PPARγ, 16 LXRα, 4 SREBP-1c, 17 and C/EBPα 3 are well-known transcription factors for increasing lipid accumulation in adipocytes. We examined the roles of kaempferol in regulating these transcription factors and our data showed that kaempferol reduced gene and protein expression levels of PPARγ, LXRα, SREBP-1c, and C/EBPα in a concentration-dependent manner. Consistently, mRNA and protein expression of their downstream genes such as FAS, SCD-1, and ACCα were significantly decreased by kaempferol treatment (Fig. 7). These data suggest that the inhibitory effect of kaempferol on lipid accumulation in adipocytes is probably mediated by suppressing a broad range of adipogenic transcription factors rather inhibiting a specific pathway. However, mechanisms underlying kaempferol-mediated inhibition of adipogenic transcription factors are unclear. An in vitro study using adipocytes indicated that kaempferol suppressed PPARγ transcriptional activity. 18 Furthermore, it has been suggested that transcriptional coactivator with PDZ-binding motif (TAZ) plays an important role in regulating PPARγ transcriptional activity. 19 TAZ has been shown to stimulate osteoblast differentiation and suppress adipocyte differentiation from mesenchymal stem cells by activating runt-related transcription factor 2 and inhibiting PPARγ transcriptional activity, respectively. 19 Interestingly, kaempferol activated TAZ and thereby suppressed PPARγ transcriptional activity, 19 suggesting that TAZ may be a target of kaempferol to inhibit transcriptional activity of PPARγ.
TG content in adipocytes is a balance between lipid accumulation and lipid breakdown. Although we did not directly examine the effect of kaempferol on lipolysis, it is unlikely that kaempferol increases lipolysis to reduce TG content because kaempferol treatment in differentiated adipocytes did not decrease TG content (Fig. 5). Therefore, we assumed that the inhibitory effect of kaempferol on TG content is probably through suppressing the early stage of TG accumulation in adipocytes.
Adipocytes are able to remove glucose from blood to store excessive energy as TGs. To investigate whether kaempferol also plays a role in regulating glucose uptake, we measured mRNA expression levels of GLUT4, a major glucose transporter in adipocytes, and glucose uptake activity. Interestingly, kaempferol notably decreased glucose uptake in a concentration-dependent manner (Fig. 8b). Consistently, GLUT4 mRNA expression was markedly reduced by kaempferol treatment (Fig. 8a). These data indicate that kaempferol suppresses substrate availability for TG synthesis in adipocytes. However, a contrary result was also reported. Kaempferol treatment in adipocytes increased glucose uptake without changing adipogenic activity. 20 We assume that the opposite results may come from different experimental conditions. While kaempferol was treated from day 9 of adipocyte differentiation in the other study, it was treated before adipocyte differentiation (day 0) in our study. Because GLUT4 expression was regulated by adipogenic transcription factors including PPARγ, it is likely that the strong inhibitory effect on adipogenic transcription factors by earlier treatment of kaempferol (day 0) also inhibit GLUT4 gene expression and thereby reduce glucose uptake in our experimental conditions.
Proper lipid accumulation in adipocytes is essential to maintain systemic insulin sensitivity and prevent fatty liver because ectopic lipid accumulation in other tissues such as liver and skeletal muscle is closely associated with insulin resistance and fatty liver. 21 Although kaempferol appears to reduce lipid accumulation in adipocytes, it also increases insulin sensitivity and reduces fatty liver in a high fat-fed condition. 15,22 Our study revealed that kaempferol may bind to and activate PPARα and stimulate mRNA expression of genes related to fatty acid oxidation (Fig. 9a, d). Although further studies are necessary, it is likely that kamepferol reduces lipid accumulation in adipocytes, but excess lipids may not go to the circulation and not be stored in other tissues such as liver and skeletal muscle because of PPARα-mediated increase in fatty acids oxidation in adipocytes. Therefore, the increase in PPARα signaling may contribute to kaempferol-mediated decrease in lipid accumulation in adipocytes and reduced metabolic syndrome in high fat-fed animals. 15,22
In conclusion, our study provides evidence that treatment with physiologically relevant levels of kaempferol efficiently decreases TG accumulation in adipocytes presumably by inhibiting a wide range of adipogenic transcription factor expression and substrate supply for TG synthesis and by increasing PPARα-mediated fatty acid oxidation signaling.
Footnotes
Acknowledgment
This work was supported by a Research Grant of Pukyong National University (2013).
Author Disclosure Statement
No competing financial interests exist.