
Abstract
Korean red ginseng (KRG), a highly valuable medicinal herb in oriental societies, has biological activity similar to that of Panax ginseng. Recently, it has been discovered that the biological activities of red ginseng can vary according to heating and steaming processes under different conditions that change the principal components of KRG and result in changes in biological activity. This study evaluated and compared the effects of high temperature- and high pressure-treated red ginseng (HRG) and commercial red ginseng (RG) on β-oxidation in C2C12 myotubes. HRG enhanced the phosphorylation levels of AMP-activated protein kinase (AMPK) and acetyl-CoA carboxylase (ACC), but RG did not affect the phosphorylation of AMPK in C2C12 myotubes. HRG also promoted the nuclear translocation of forkhead box protein O1 (FoxO1), and the translocation exerted an increase in the protein expression of adipose triglyceride lipase (ATGL). As a consequence, HRG increased the mRNA expression level of carnitine palmitoyltransferase 1 (CPT-1) compared to the control. Taken together, our results indicated that HRG promotes the lipolysis of triglycerides and mitochondrial β-oxidation of fatty acids in C2C12 myotubes, suggesting that alterations to the principal components by high temperature and pressure may positively influence the nutraceutical functions of HRG.
Introduction
K
The biological activities of KRG have been shown to include regulation of the AMP-activated protein kinase (AMPK) signaling pathway. 5,6 AMPK is an enzyme that acts as an energy sensor and a metabolic master switch in response to changes in cellular energy charge. 7 AMPK consists of heterotrimeric complexes composed of catalytic α subunits and regulatory β and γ subunits. Among the activation domains of the α subunit kinase, the phosphorylation at threonine-172 activates AMPK. 8 Activated AMPK beneficially influences the body by increasing fatty acid oxidation and glucose uptake. 9,10 In addition, AMPK regulates the translocation of forkhead box protein O1 (FoxO1) from the cytosol to the nucleus and subsequently increases the adipose triglyceride lipase (ATGL) expression. 11
However, it is necessary to investigate whether changes to the components by the different conditions during KRG production process actually lead to the change in biological activities at the molecular level. Therefore, we tested how high temperature and high pressure influence the formation of various derivatives from red ginseng, but also how the alterations affect biological activity such as β-oxidation through AMPK signaling pathway in muscle using C2C12 muscle cells.
Materials and Methods
Reagents
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), horse serum (HS), penicillin–streptomycin (P/S), phosphate-buffered saline (PBS), and trypsin-EDTA were purchased from Gibco (Gaithersburg, MD, USA). Bradford reagent, Oil Red O, palmitate (PA), and bovine serum albumin (BSA) were obtained from Sigma (St Louis, MO, USA). Antibodies against AMPKα, phospho-AMPK (p-AMPK) α Thr172, acetyl-CoA carboxylase (ACC), and phospho-ACC (p-ACC)ser79 and β-actin were acquired from Cell Signaling Technology (Beverly, MA, USA). Antibody against lipoprotein lipase (LPL) was obtained from Abcam (Cambridge, MA, USA).
Preparation of high temperature- and high pressure-treated red ginseng extract
Five-year-old ginseng and commercial red ginseng (RG) were purchased from ginseng grocery store (Geumsan, Korea). High temperature- and high pressure-treated red ginseng (HRG) was produced from the 5-year-old ginseng according to the method developed by the Korea Food Research Institute. In brief, 5-year-old ginseng roots were cleaned and first dried at 50°C, and then they were steamed at high temperature (140°C) and high pressure (3 kg/cm2) for 20 min. The steamed ginseng was dried at 50°C until the moisture content reached below 15%. HRG and RG were ground to 60 mesh in a mill (Cyclotec™ 1093; FOSS Co., Kyoto, Japan) and refluxed with 5 volumes of hot water at 80°C for 3 h. After reflux, HRG and RG extracts were filtered two times and concentrated at 55°C under vacuum pressure. Then, the concentrates were freeze dried and stored at −20°C until used.
Analysis of ginsenoside contents in HRG and RG
Each 300 mg sample of HRG and RG extract was dissolved in 50% methanol solution and centrifuged at 2300 g for 10 min, and then the supernatants were concentrated at 50°C. The concentrates were dissolved in the mixture of distilled water and acetonitrile at a ratio of 8:2 and filtered with a Millipore membrane filter (0.45 μm) before high-performance liquid chromatography (HPLC) analysis. The analysis of ginsenosides was conducted using JASCO HPLC (Tokyo, Japan) equipped with MD-2010 Plus multi-wavelength detector. The dissolved sample was separated on a SunFire™ C18 column (5 μm, 4.6 × 250 cm; Waters, Wexford, Ireland). The mobile phase was composed of two solvents: solvent A was distilled water and solvent B was acetonitrile.
Cell culture
C2C12 skeletal muscle cell was obtained from the American Type Culture Collection (Manassas, VA, USA). The C2C12 myoblasts were maintained in DMEM containing FBS (10%) and P/S (1%), and cultured in a humidified atmosphere of 5% CO2 at 37°C. For inducing the differentiation into myotubes, the medium was replaced with DMEM containing HS (10%) and P/S (1%) when the cells reached 80–90% confluency in each well. The medium was refreshed every other day until the cells fully differentiated into myotubes. The fully differentiated cells were maintained in the differentiation DMEM containing 0.5 mM PA and fatty acid-free BSA (2%) for 12 h and then exposed to samples for 24 h.
Cell viability
The viabilities of myoblasts and myotubes were examined by the XTT Assay Kit. First, the C2C12 myoblasts were seeded in a 96-well plate and then exposed to HRG and RG for 24 h. For myotube's viability, the myoblasts were differentiated into myotubes in DMEM with 10% HS and 1% P/S for 6 days after roughly 80% confluence. After fully differentiated, the myotubes were preincubated in the differentiation media containing PA (0.5 mM) and fatty acid-free BSA (2%) for 12 h and then treated with the samples for 24 h. All the cells after the treatment with the samples for 24 h were further incubated after adding the mixture containing XTT reagent and PMS reagent to each well. After 4 h, the absorbance was measured at 490 and 660 nm. The growth medium alone was used as a blank. The specific absorbance of the sample is expressed as follows: Specific
Oil Red O staining
To measure the lipid content in C2C12 myotubes, the cells were stained with Oil Red O as described by Blumberg et al. 12 with slight modifications. After 6 days, the fully differentiated C2C12 myotubes were washed with PBS (pH 7.4) twice and fixed with 10% formaldehyde for 1 h. Then the added formaldehyde was removed and washed with 60% isopropanol and completely dried. An Oil Red O stock solution was made by mixing 2 g of Oil Red O powder with 100 mL of isopropanol and the stock solution was mixed with distilled water at a ratio of 60:40. The cells were stained with the mixture for 15 min at room temperature, washed four times with distilled water, and completely dried. The red dye in the cells was eluted with isopropanol and the absorbance was measured by a microplate reader (Molecular Devices, Sunnyvale, CA, USA) at 490 nm.
RNA isolation and semi-quantitative real-time polymerase chain reaction
Total RNA was extracted from C2C12 myotubes with TRIzol reagent (Invitrogen, Carlsbad, CA, USA). RNA with OD260/OD280 ratios higher than 2.0 was used for semi-quantitative reverse transcription–polymerase chain reaction (RT-PCR). One microgram of total RNA was employed for the production of cDNA and the sequence of the oligonucleotide primer was as follows: carnitine palmitoyltransferase 1 (CPT-1) sense (5′-ACT CCT GGA AGA AGA AGT TCA-3′) and antisense (5′-AGT ATC TTT GAC AGC TGG GAC-3′); β-actin sense (5′-AGG CTG TGC TGT CCC TGT ATG C-3′) and antisense (5′-ACC CAA GAA GGA AGG CTG GAA A-3′). The PCR products were run on 1.5% agarose gels, stained with ethidium bromide, and photographed. The expression levels were quantified by Carestream MI SE software.
Western blotting
Cells were lysed using protein lysis buffer (50 mM Tris-HCl, 1 mM EDTA, 1 mM EGTA, 150 mM NaCl, 1% Triton X-100, 0.1% sodium dodecyl sulfate [SDS], 0.25% sodium deoxycholate, 1 mM PMSF, 1 mM benzamidine, 5 μg/mL of aprotinin, 5 μg/mL of leupeptin, 5 μg/mL of pepstatin, 1 mM sodium orthovanadate, 1× phosphatase inhibitor cocktail II, 1× phosphatase inhibitor cocktail III, 1 mM sodium fluoride, 1 mM sodium pyrophosphate, β-mercaptoethanol, and 1 mM β-glycerophosphate). After that, lysates were clarified by centrifugation at 20,800 × g for 20 min at 4°C and the supernatants were transferred to new tubes. The concentration of each lysate was determined using Bradford reagent and equal amounts (50 μg) of proteins from lysates were separated through 6% or 10% SDS-polyacrylamide gel electrophoresis and transferred to PVDF membranes. The membranes were blocked by 3% BSA for 30 min at room temperature and incubated overnight with 3% BSA containing the indicated primary antibody at 4°C. On the following day, the membranes were incubated with horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. Detection of each protein was performed using an enhanced chemiluminescence kit (SuperSignal™ West Pico Chemiluminescent Substrate; Thermo Scientific, West Palm Beach, FL, USA). The proteins were detected using ChemiDoc™ imaging system (Bio-Rad, Hercules, CA, USA) and quantified using Image Lab™ software. After quantifying all proteins detected, the expression level of each target protein was normalized with β-actin.
Statistical analysis
Data are expressed as the mean ± standard deviation. The data were analyzed by one-way analysis of variance conducted by Duncan's test at a significance level of P < .05 (version 9.2; SAS Institute, Inc., Cary, NC, USA). Mean without common letters is significantly different.
Results
The contents of ginsenosides in HRG and RG
The ginsenoside contents of HRG and RG were analyzed by HPLC. As shown in Table 1 and Figure 1, the content of most ginsenosides, including Re, Rb1, Rb2, Rb3, and Rd, was higher in RG than those of HRG. Among the ginsenosides, however, ginsenoside Rh1(R) was only detected in HRG and not RG. Moreover, the amount of ginsenoside Rg1 in HRG (3.33 ± 0.12 mg/g) was higher than that of RG (2.41 ± 0.14 mg/g). Although the content of most ginsenosides in HRG were lower than that of RG, it is well known that heat-treated ginseng is more pharmacologically active than raw ginseng, as the heating process causes changes in the composition and ratio of ginsenosides, thereby enhancing the biological activity of the ginseng. 13 Furthermore, we previously reported that ginsenoside Rg1 promotes the AMPK signaling pathway in muscle cells. 14 Therefore, we questioned whether changes in the composition and ratio of ginsenosides are correlated with changes in physiological activity involved in lipolysis and β-oxidation through regulation of AMPK signaling pathway.
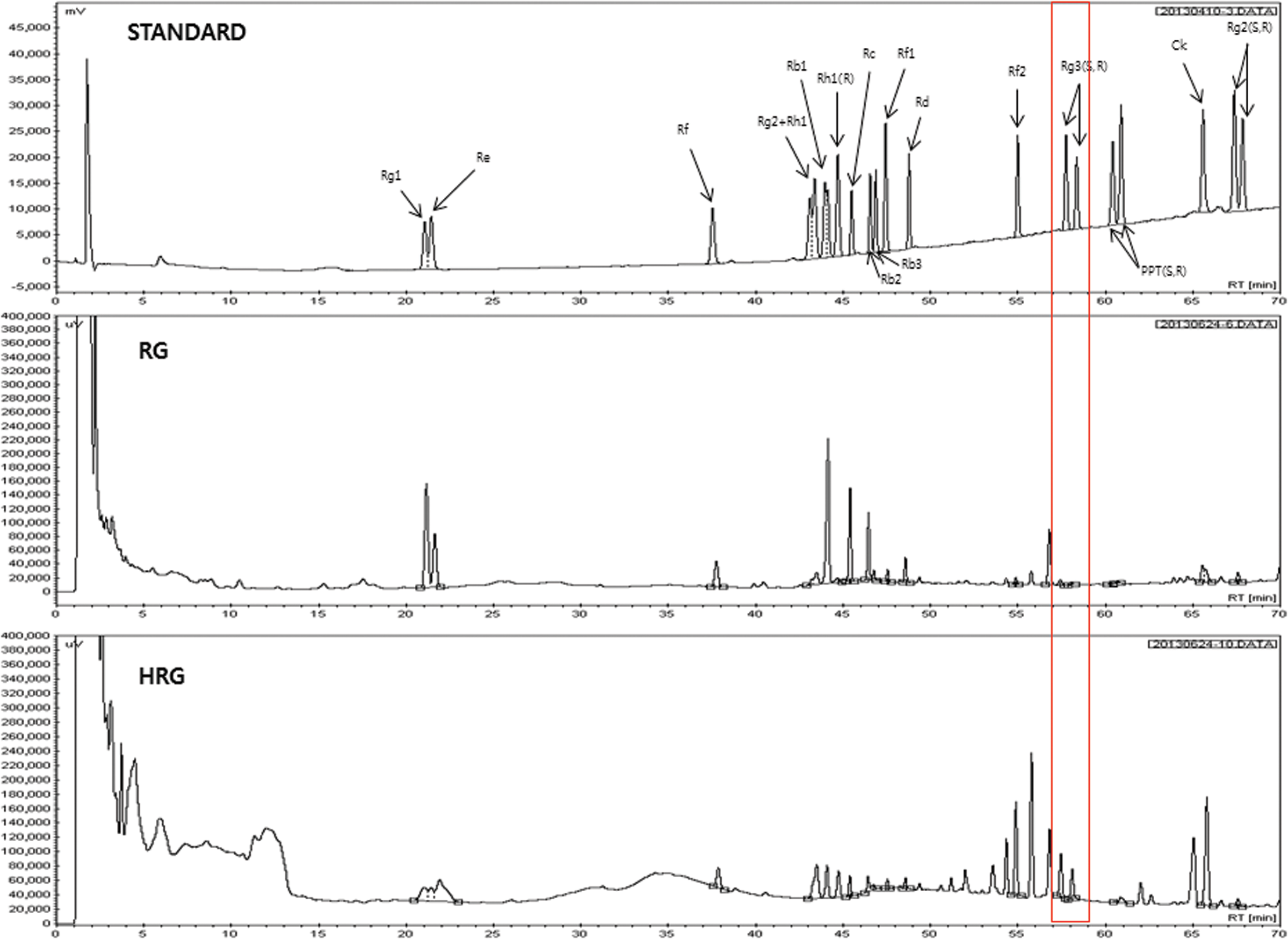
High-performance liquid chromatography chromatogram of ginsenosides standard, HRG and RG. The significance of the red line box is ginsenoside Rg3 (S, R) which is a major active compound in HRG. HRG, high temperature- and high pressure-treated red ginseng; RG, commercial red ginseng. Color images available online at
Values are mean ± standard deviation (n = 3).
RG, commercial red ginseng; HRG, high temperature- and high pressure-treated red ginseng.
Effect of HRG on cell viability
Since HRG and RG both significantly prevented oxidative stress resulting from reactive oxygen species production during preadipocyte differentiation and also markedly inhibited H2O2-induced oxidative stress in myotubes at 200 μg/mL in our previous report, 15 we used the concentration (200 μg/mL) and tested whether the concentration is appropriate for this study (Fig. 2). As we expected, HRG and RG did not show any cytotoxicity to the cells we used at 200 μg/mL. Therefore, further studies were conducted with the concentration of 200 μg/mL.
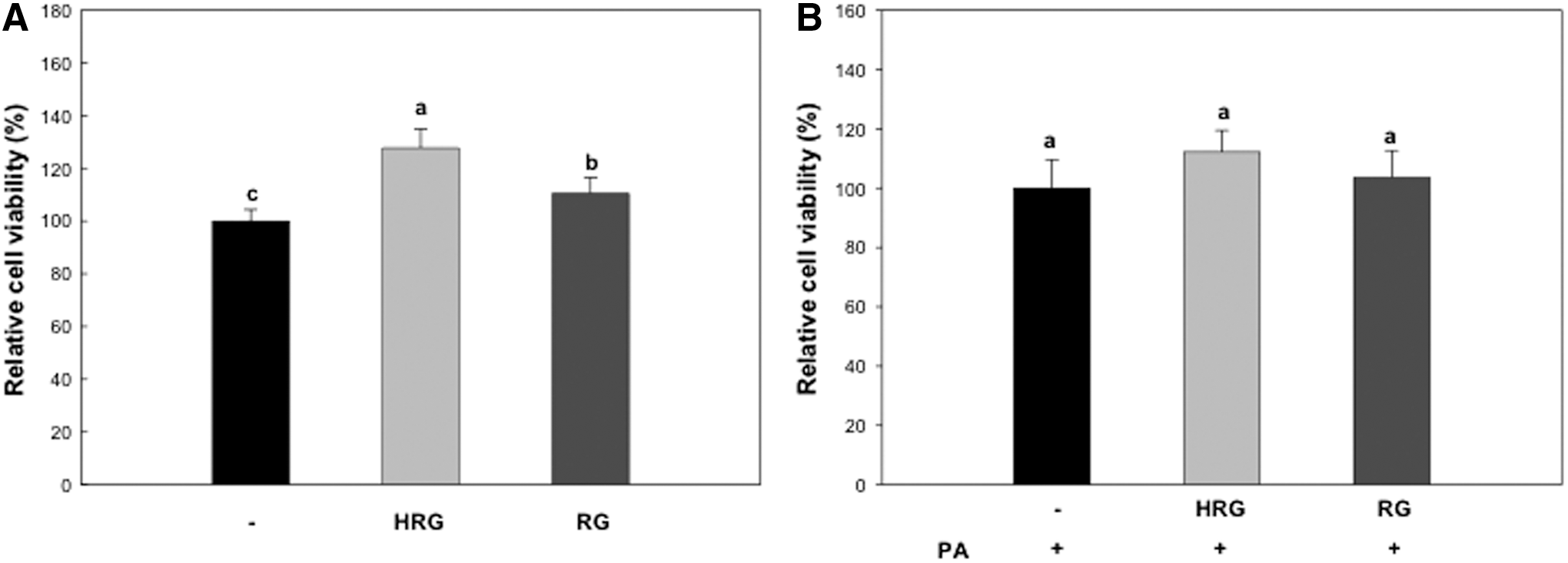
Effect of HRG and RG on cell viability.
Effect of HRG on the phosphorylation of AMPK and ACC
To investigate whether HRG and RG regulates β-oxidation through the AMPK signaling pathway, fully differentiated C2C12 myotubes were exposed to PA (0.5 mM) because PA is known to induce lipid accumulation in myotubes (Richmond et al. 16 ). Fully differentiated C2C12 myotubes were preincubated with 0.5 mM PA for 12 h and then exposed to HRG or RG for 24 h. The expression and phosphorylation levels of AMPK and ACC were determined by western blot analysis. As shown in Figure 3, HRG increased the phosphorylation of AMPK and also upregulated the phosphorylation of ACC, a key downstream target related to β-oxidation. However, RG did not influence the phosphorylation of AMPK or ACC.
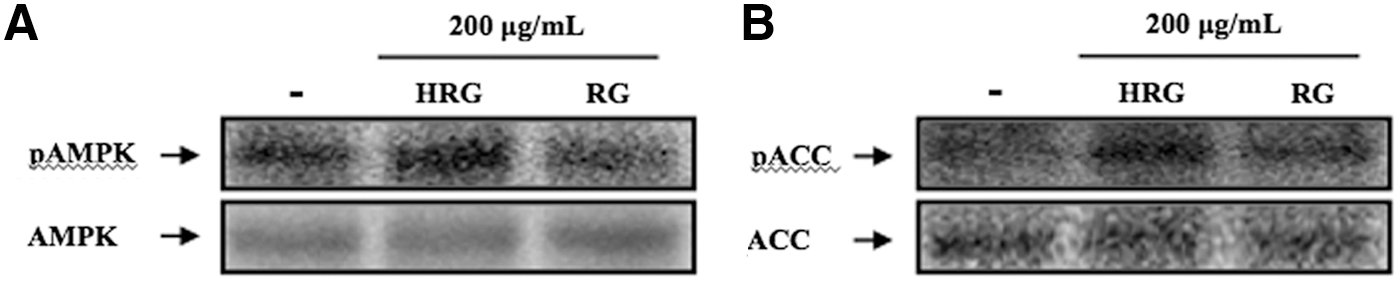
Effect of HRG and RG on the phosphorylation of AMPK
Effect of HRG on the mRNA expression of CPT-1 and lipolysis
We also investigated the effects of HRG and RG on the mRNA expression of CPT-1 and triglyceride contents in myotubes. As shown in Figure 4, HRG treatment significantly enhanced the mRNA expression of CPT-1 compared to the control. PA treatment induced lipid accumulation in C2C12 myotubes. HRG significantly decreased the lipid accumulated in C2C12 myotubes compared to the PA-treated cells. Treatment of RG also increased the mRNA expression levels of CPT-1 compared to the control. However, RG significantly reduced the triglyceride contents in myotubes when compared to the PA-treated cells. These results indicate that HRG and RG could enhance mRNA expression of CPT-1 and inhibit lipid accumulation in C2C12 myotubes.
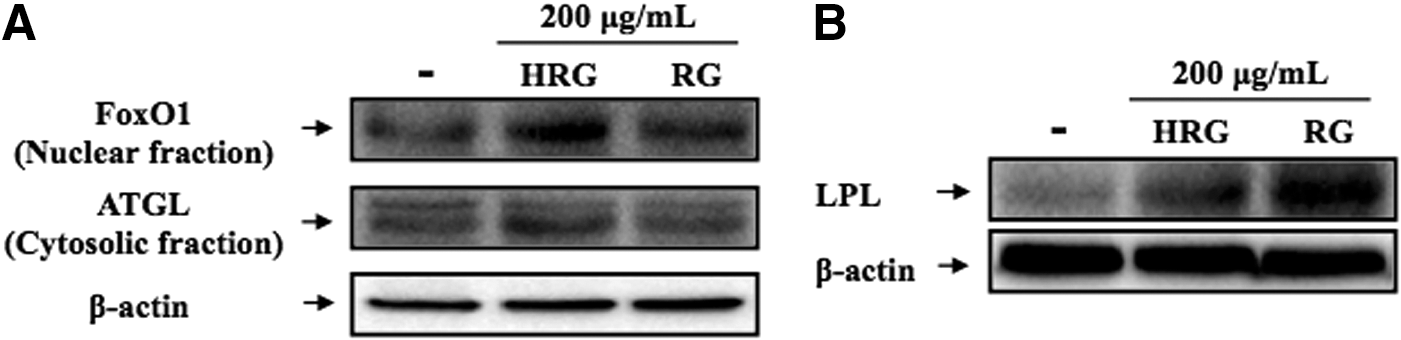
Effect of HRG and RG on FoxO1 translocation, ATGL
Effect of HRG on FoxO1 translocation, ATGL, and LPL
To investigate whether HRG and RG exerts a lipid-lowering effect through regulating lipolytic enzymes, the protein levels of ATGL and LPL were examined by western blotting (Fig. 5). HRG and RG increased the translocation level of FoxO1 from cytosol to nucleus, suggesting that HRG and RG could regulate the translocation of FoxO1 (Fig. 5A). HRG enhanced the expression level of ATGL compared to the control, but RG did not affect the expression level of ATGL. 16 In addition, HRG and RG upregulated the expression level of LPL compared to the control (Fig. 5B). In particular, RG remarkably increased the LPL protein expression compared to the HRG.

Effect of HRG and RG on the mRNA expression of CPT-1
Discussion
KRG, a popular medicinal herb, has various biological activities such as improving exercise capacity and attenuating oxidative stress. 17,18 It was recently suggested that the differences in pharmacological activities between red ginseng and raw ginseng might result from changes in their constituents during the heating and steaming process, and the process might be one of the major elements to promote the enhancement of red ginseng's biological activity. 4 Moreover, the high pressure used to produce red ginseng from ginseng exerts a major positive influence on appearance, total reducing sugars, and amino acid contents, as well as ginsenosides in red ginseng, suggesting that these changes may give rise to changes in biological properties of the nutraceutical. 4,19 In this present study, the high temperature and high pressure conditions led to primary components attributable to a difference in pharmacological activities (Table 1). According to several studies, red ginseng attenuates insulin sensitivity, diabetes, and exercise-induced muscle damage. 18,20,21 Ginseng is also reported to modulate lipid metabolism, although the mechanism of its action remains unknown. Therefore, we investigated whether high temperature and high pressure conditions can lead to changes in the bioactives of red ginseng, but also contribute to its biological effect on β-oxidation involving in AMPK activation using C2C12 muscle cells.
AMPK is an enzyme that plays a role in cellular energy homeostasis and regulates fatty acid β-oxidation in skeletal muscle. 22 AMPK increases the phosphorylation of ACC, which inhibits the generation of malonyl-CoA and subsequently activates the CPT-1 in mitochondria. 23 Recently, it was shown that red ginseng can regulate oxidative stress in hepatocytes and free fatty acids in skeletal muscle and liver tissues through the AMPK signaling pathway. 6,24,25 In the present study, HRG induced the phosphorylation of AMPK and also phosphorylated ACC. In addition, HRG remarkably increased the mRNA expression of CPT-1, thereby significantly decreasing the accumulated lipids in C2C12 myotubes, suggesting that free fatty acids released from lipids in C2C12 myotubes were oxidized by β-oxidation (Fig. 4). However, AMPK and ACC were not phosphorylated by RG treatment, but increased the mRNA expression of CPT-1 slightly (Fig. 4B). According to Lee et al., 5 KRG contributed to improving insulin sensitivity and attenuating diabetes in fatty rats and C2C12 muscle cells. These findings are thought to result from the differences in ginsenoside constituents as well as phenolic compounds, especially maltol in red ginseng, suggesting that the biological activities of red ginseng may differ depending on the components in red ginseng and the conditions that are utilized to make red ginseng.
RG significantly decreased lipid accumulation in C2C12 cells. Therefore, we investigated whether HRG and RG modulate lipolytic protein, such as ATGL and LPL. Inoue et al. 26 reported that red ginseng modulated serum TG content and concomitant cholesterol by increasing LPL activity. Indeed, RG significantly increased the LPL protein levels in C2C12 myotubes when compared to HRG-treated C2C12 myotubes. It has been reported that the translocation of FoxO1 protein into the nucleus induced by the phosphorylation of AMPK regulates the ATGL protein expression level. 11 In addition, LPL in skeletal muscle is also regulated through the activation of AMPK. 27 In this study, HRG increased the protein level of LPL and also upregulated the ATGL protein level by translocating FoxO1 into the nucleus, suggesting that HRG may increase LPL, FoxO1 translocation, and ATGL by modulating AMPK phosphorylation. RG, however, did not influence the FoxO1 translocation and ATGL protein level, but remarkably increased the LPL protein level in C2C12 myotubes without the activation of AMPK. Therefore, these results indicate that HRG produced under high temperature and high pressure conditions can modulate lipid metabolism through the activation of AMPK and lipolytic proteins, which are involved in β-oxidation, suggesting that the difference in ginsenoside profiles may be a result of heating conditions utilized to make red ginseng and may lead to the changes in biological activity between RG and HRG.
Footnotes
Author Disclosure Statement
No competing financial interests exist.