
Abstract
The antiobesity effect of ice plant (IP) (Mesembryanthemum crystallinum), a salt-resistant African plant, has recently attracted increased attention. IP is rich in pinitol, which lowers blood sugar, and myo-inositol, which prevents fatty liver disease. Furthermore, IP can potentially prevent or reduce the symptoms of metabolic syndrome. However, the details of the physiological mechanisms and mechanisms of action of IP are unclear. A previous study by our group demonstrated the capability of IP extract to prevent adipogenesis in 3T3-L1 preadipocytes. In this study, we analyzed the physiological function of IP extract on lipolysis in 3T3-L1 cells and the underlying mechanisms of this process. We found that the release of glycerol from cells treated with IP extract increased in an IP dose-dependent manner. IP extract exhibited cytotoxic activity at concentrations above 4 mg/mL. Real-time polymerase chain reaction and western blotting showed that IP extract downregulated peroxisome proliferator-activated receptor (PPAR-)γ, hormone-sensitive lipase (HSL), and adipose triglyceride lipase (ATGL) in a concentration-dependent manner, but did not affect HSL-Ser563, HSL-Ser660, or perilipin phosphorylation. Although the cAMP-dependent protein kinase A (PKA)-specific inhibitor H89 did not affect IP extract-induced lipolysis, the extracellular signal-regulated kinase (ERK1/2) inhibitor U0126 significantly abrogated IP extract-activated glycerol release. Furthermore, IP extract strongly enhanced ERK1/2 phosphorylation at the concentrations used in the study. These results suggest that IP extract augments lipolysis by enhancing ERK phosphorylation.
Introduction
L
In recent years, plant-derived physiologically active substances (phytochemicals) have been associated with the reversal and prevention of obesity. The efficacy and mechanisms of action of phytochemicals vary enormously, and each phytochemical can exert a range of physiological actions at various sites of action; caffeine and ephedrine enhance energy consumption, capsaicin and berberine inhibit lipid formation, and flavonoids and conjugated linoleic acid promote lipolysis. 10
In this study, we focused on the antiobesity effect of ice plant (IP) (Mesembryanthemum crystallinum). IP, which is native to Africa, is a salt-resistant plant that produces substances useful for controlling the damage by salt and ultraviolet light. IP contains numerous functional substances, including myo-inositol, which acts to prevent fatty liver disease, and pinitol, which lowers blood sugar and increases insulin sensitivity, as well as the antioxidants β-cyanins, proline, pantothenic acid, β-carotene, and other flavonoids. 11 –16 Previous studies have shown that crude extracts of IP inhibit the differentiation of mouse 3T3-L1 preadipocytes. 17
This study aimed to determine the lipolysis-related physiological functions of IP extract and to elucidate the mechanisms of action of IP by using 3T3-L1 adipocytes.
Materials and Methods
Materials
Dried powder made from IP grown in indoor planters (Tsuburina; Nihon Advanced Agri Co., Shiga, Japan) was used in the experiments. IP powder was dissolved in distilled water and passed through a filter paper. The supernatant was filter sterilized, and the IP samples thus obtained were stored at −80°C. The concentrations of IP are expressed as the weight of IP dissolved in 1 mL of culture medium. H89 and U0126 were purchased from Cayman Chemical (Ann Arbor, MI), dissolved in DMSO, and stored at −80°C.
Cell culture
Mouse 3T3-L1 preadipocytes (Institute of Physical and Chemical Research Cell Bank, Tsukuba, Ibaraki, Japan) were cultured in DMEM high glucose (Dulbecco's modified minimum essential; 4500 mg/L glucose [pH 7.3]) (Sigma-Aldrich, St. Louis, MO) containing 10% fetal bovine serum (Sanko Junyaku, Tokyo, Japan), 100 U/mL penicillin (Wako, Osaka, Japan), and 100 μg/mL streptomycin (Meiji Seika, Tokyo, Japan). Cells were cultured in either 24-well plates at 5 × 104 cells/well or 60-mm dishes at 3 × 105 cells/dish and maintained in a 5% CO2 incubator at 37°C (NAPCO, Amityville, NY). After the cells reached confluence, they were cultured for 2 days in DMEM high glucose culture medium containing DMI (1 μM dexamethasone (Sigma-Aldrich), 500 μM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), and 10 μg/mL insulin (Wako)). After incubation with DMI for 2 days, the cells were cultured for an additional 2 days in DMEM high glucose containing 5 μg/mL insulin. The culture medium was then exchanged for DMEM high glucose with no additives. 3T3-L1 adipocytes were used at day 10 for further experiments.
Glycerol release assay
The amount of glycerol released from adipocytes into the culture medium was analyzed using an Etest Wako kit (Wako). Absorbance at 600 nm was measured using a SpectraMax microplate reader (SpectraMax 190; Molecular Devices Corporation, Sunnyvale, CA) and converted to the amount of glycerol. Total protein was measured using a BCA Protein Assay Kit (Reducing Agent Compatible; Wako).
Cell viability assay
Thiazolyl Blue Tetrazolium Bromide (MTT; Sigma-Aldrich) was used to measure the cytotoxicity of IP. 18 Briefly, 10-day-old 3T3-L1 adipocytes, cultured in 24-well plates, were incubated with IP extract (0–5 mg/mL) for 48 h. MTT (5 mg/mL) was added to the culture medium at a ratio of 1:10, and the cells were cultured for an additional 3 h in the CO2 incubator. The culture medium was then removed, formazan was dissolved in 0.04 mol/L HCl/isopropanol, and the absorbance was measured at 570 nm.
Real-time polymerase chain reaction analysis
RNA extraction was performed according to the acid guanidinium–phenol–chloroform method. 19 After treatment with DNase I (TaKaRa Bio, Otsu, Shiga, Japan), M-MLV reverse transcriptase (TaKaRa Bio) was used to synthesize cDNA. Real-time polymerase chain reaction (PCR) was performed using the KAPA SYBR FAST qPCR Kit (Kapa Biosystems, Boston, MA). cDNA was amplified using 40 cycles of 3 min at 95°C, 3 s at 95°C, and 20 s at 60°C. The following primers were used: β-actin (5′-AGATGTGGATCAGCAAGCAGG-3′ and 5′-AACGCAGCTCAGTAACAGTCC-3′), peroxisome proliferator-activated receptor γ (PPARγ) (5′-AAACTCTGGGAGATTCTCCT-3′ and 5′-TGGCATCTCTGTGTCAAC-3′), HSL (5′-GCTGTCTGAAGGCTCTGAGTTGC-3′ and 5′-AAGACCACATCGCCCACAGC-3′), and adipose triglyceride lipase (ATGL) (5′-TGCTGGAGGCCTGTGTGGAA-3′ and 5′-TCAGGGACATCAGGCAGCCACT-3′). β-actin was used as an internal control.
Western blot analysis
3T3-L1 adipocytes were harvested in RIPA buffer (50 mmol/L Tris-HCl [pH 7.4], 150 mmol/L NaCl, 0.1% SDS, 0.5% Na-deoxycholate, 1% NP40, 1 mmol/L EDTA, 2 mmol/L NaF, and 1 mmol/L Na3VO4). After protein transfer, the membranes were blocked for 1 h in TBS-T buffer containing 2% bovine serum albumin. Then, primary antibodies (anti-HSL563, anti-HSL660, anti-HSL, anti-ATGL, anti-perilipin, anti-pERK, anti-ERK, and anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Cell Signaling Technology, Danvers, MA), dissolved in TBS-T buffer, were added for overnight incubation. The LumiGLO reagent (Cell Signaling Technology) was used for protein detection. The protein bands were analyzed using chemiluminescence (EZ-Capture MG; ATTO, Tokyo, Japan). The band intensity was analyzed using ImageJ analysis software.
Statistical analysis
Results are expressed as mean ± SD and statistical analysis was performed using one-way or two-way analysis of variance (ANOVA) followed by the Neuman–Keuls post hoc test, or the Student's unpaired t-test. P values below .05 were considered significant.
Results
Lipolytic effect of IP extract
The physiological effects of IP extract on lipid storage in differentiated adipocytes were analyzed. Ten days after differentiation was induced, 3T3-L1 adipocytes were cultured in the presence of IP extract (0–1.5 mg/mL). After incubation with IP for 12 or 24 h, the amount of glycerol released into the culture medium was measured (Fig. 1A). The addition of IP extract increased the glycerol content of the culture medium in a time- and concentration-dependent manner. Thus, IP extract promotes lipolysis in differentiated adipocytes. Next, we investigated the effect of IP extract on cell viability. Adipocytes differentiated for 10 days were treated with IP extract (0–5 mg/mL) for 48 h, and cell viability was measured using an MTT assay (Fig. 1B). Cell viability decreased significantly at IP extract concentrations at or above 4 mg/mL. However, cell viability was largely unaffected at IP extract concentrations at or below 3 mg/mL. Therefore, subsequent experiments were performed using IP extract concentrations below 3 mg/mL.
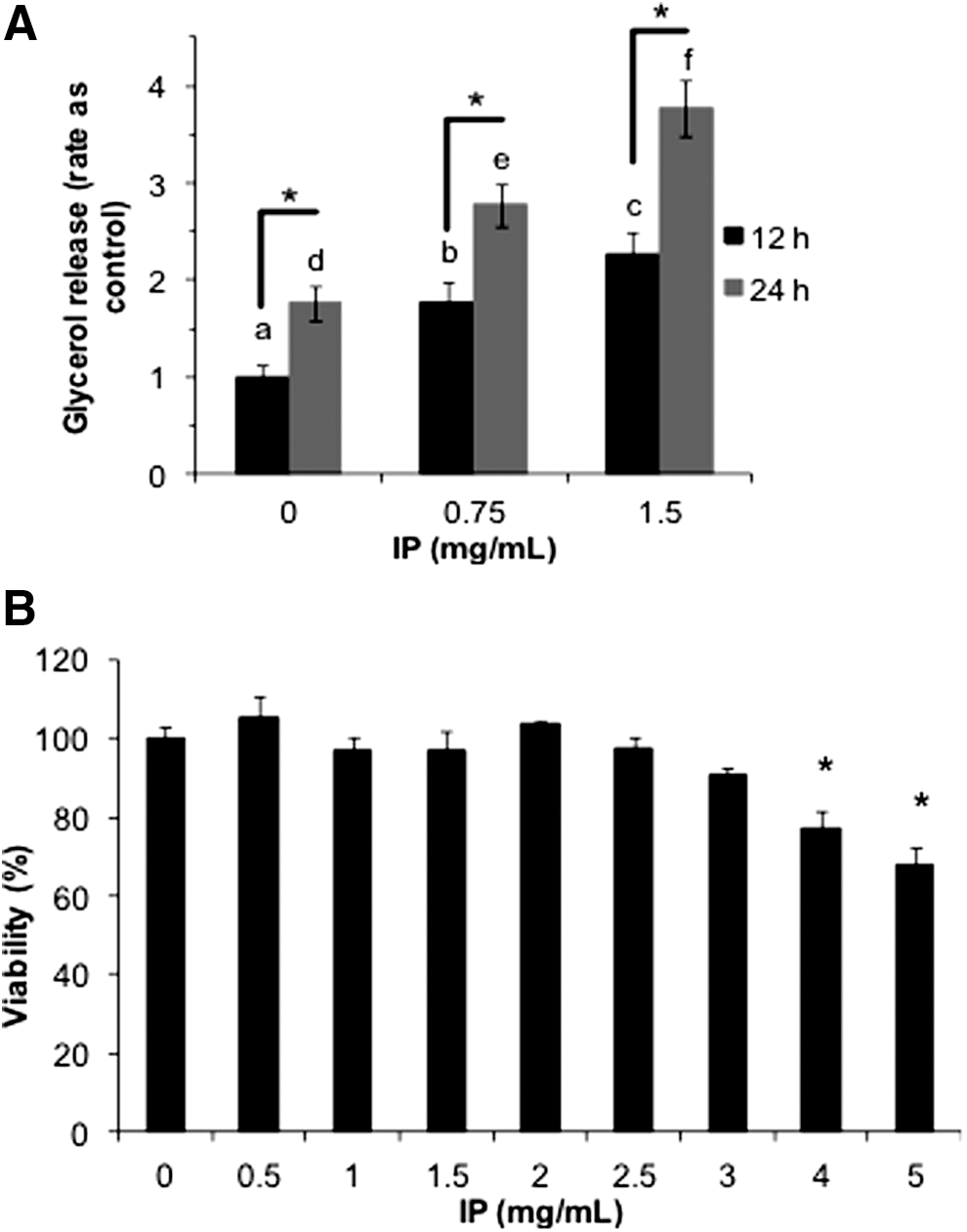
Effect of ice plant (IP) extract on adipocyte lipolysis and viability. 3T3-L1 preadipocytes were cultured for 10 days after induction with DMI (1 μM dexamethasone (Sigma-Aldrich), 500 μM 3-isobutyl-1-methylxanthine (Sigma-Aldrich), and 10 μg/mL insulin (Wako)). IP extract (0–1.5 mg/mL) was added, and after treatment for 12 or 24 h, absorption at 600 nm was measured to quantify the glycerol content of the culture media. The glycerol content was standardized according to the total protein content. The graph shows values relative to the control. Results are representative of three independent experiments with n = 3 for each. The error bars indicate SD. For statistical analysis, two-way ANOVA, followed by the Newman–Keuls post hoc test was used. a, b, and c indicate significant difference at P < .05 between different treatments at 12 h; d, e, and f indicate significant difference at P < .05 between different treatments at 24 h. *Shows significant difference at P < .05 among treatments at the same concentration
IP extract induced changes in PPARγ, ATGL, and HSL gene expression
PPARγ is involved not only in lipid formation, but also in regulating the expression of the HSL and ATGL genes, which are responsible for lipolysis. 20 Figure 2 shows the mRNA expression levels of PPARγ, ATGL, and HSL, which were dramatically lower in the growth media with added IP extract. Moreover, western blot analysis showed lower ATGL protein expression (Fig. 3A).

Effect of IP extract on gene expression. 3T3-L1 cells were differentiated for 10 days. Then, IP extract (0–1.5 mg/mL) was added, and the cells were incubated for 24 h before gene expression was investigated using real-time polymerase chain reaction. The graph shows the values relative to the control. Results are representative of three independent experiments. The error bars indicate SD.
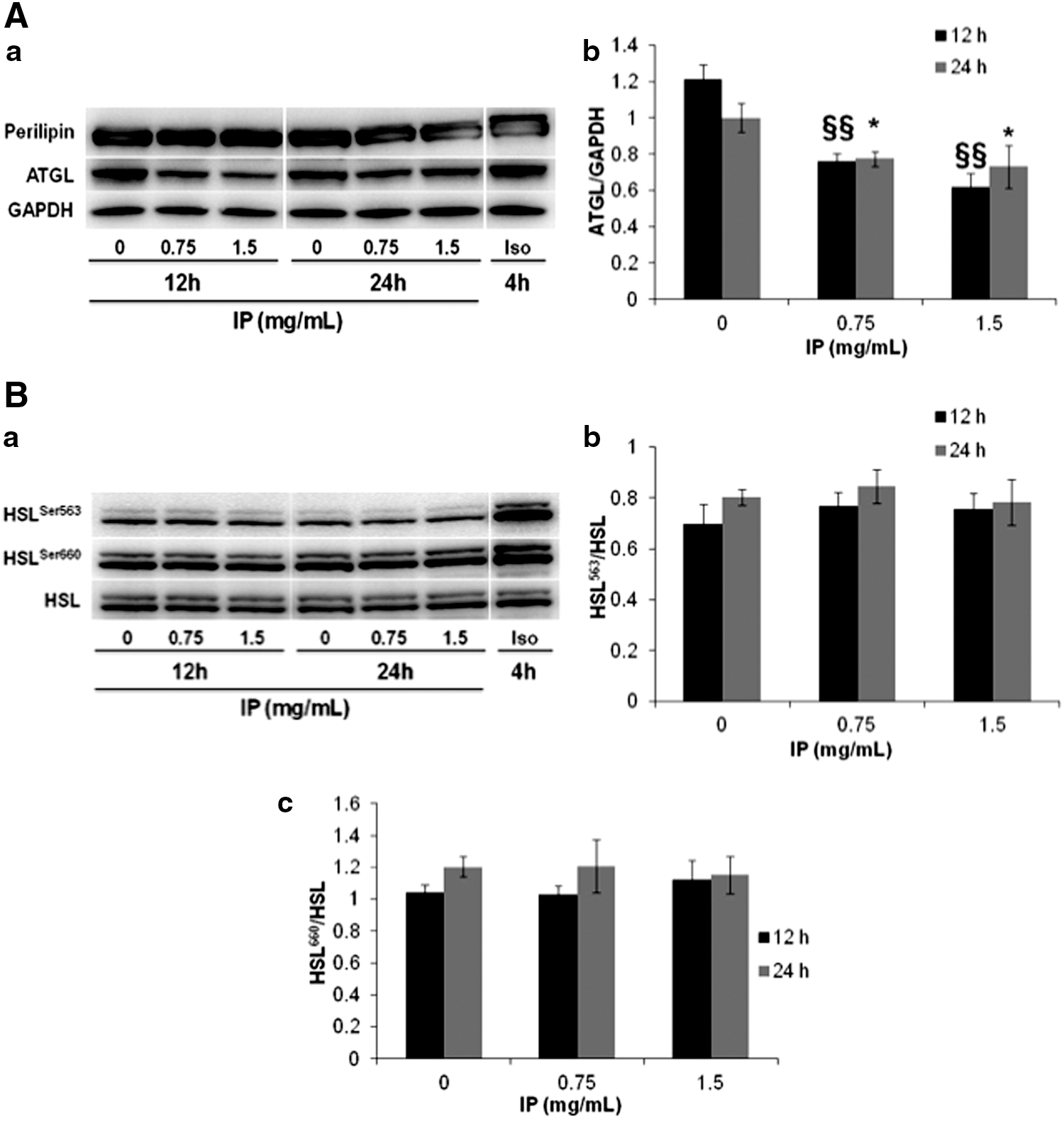
Effect of IP extract on adipose triglyceride lipase (ATGL) expression, perilipin phosphorylation, and hormone-sensitive lipase (HSL) phosphorylation. 3T3-L1 cells were cultured for 10 days after induction with DMI. Then, IP extract (0–1.5 mg/mL) or isoproterenol (30 nmol/L) was added, and the cells were incubated for 12 or 24 h. ATGL expression, perilipin phosphorylation
IP extract has no effect on HSL and perilipin phosphorylation
HSL activity is strictly controlled by phosphorylation. cAMP-activated PKA phosphorylates Ser563, Ser659, and Ser660 of HSL, whereupon HSL hydrolyzes TAG and DAG. 3,21 Figure 3 shows the western blot analysis of perilipin phosphorylation and HSL phosphorylation (Ser563 and Ser660 residues). IP extract did not enhance the phosphorylation of perilipin (Fig. 3A). Additionally, no significant difference was observed in HSL phosphorylation at the Ser563 and Ser660 residues after IP extract addition (Fig. 3B). In contrast, isoproterenol, which was used as a positive control, clearly increased the amount of the perilipin isoform at 67 kDa, reflecting an increase in phosphorylation and activation, and enhanced the phosphorylation of HSL at both sites (Ser563 and Ser660).
Suppression of IP lipolysis by the ERK1/2 inhibitor U0126
HSL enzyme activity is regulated by phosphorylation; HSL can be activated by phosphorylation through the PKA and ERK1/2 pathways. Therefore, we investigated the effect of PKA and ERK1/2 pathway inhibitors (H89 and U0126, respectively) on the lipolytic effect of IP extract. H89 is a selective inhibitor of PKA, while U0126 selectively inhibits MAPK/ERK1/2 kinase (MEK), which activates ERK1/2. Differentiated adipocytes were cultured for 1 h in the presence of either H89 (10 μM) or U0126 (50 μM). IP extract (0–1.5 mg/mL) was then added to the adipocytes. After incubation for 12 and 24 h, the glycerol content of the culture medium was measured. The lipolysis-enhancing effect of IP extract was not reduced by H89 (Fig. 4A). However, U0126 significantly inhibited IP extract-induced lipolysis. Consistent with the inhibitor assay, western blot analysis indicated that IP extract strongly induced the phosphorylation of ERK1/2 kinase in a concentration-dependent manner (Fig. 4B). This IP extract-induced phosphorylation was effectively attenuated by the U0126 inhibitor. These data indicate that the ERK1/2 phosphorylation pathway is involved in IP extract-enhanced lipolysis.

Effect of protein kinase A (PKA) inhibitor (H89) and ERK inhibitor (U0126) on IP extract-stimulated lipolysis
P < 0.01 versus the IP extract (1.5 mg/mL) treatment in each group was regarded as statistically significant (Student's t-test)
Discussion
In this study, we showed that IP extract enhances lipolysis in mature adipocytes, and is not cytotoxic (Fig. 1). We also showed that IP extract is involved in lipolysis by enhancing ERK1/2 kinase phosphorylation (Fig. 4).
ATGL expression is increased by PPAR agonists and glucocorticoids, whereas insulin decreases its expression. Recent findings showed that mTOR complex 1 (mTORC1)-dependent signaling reduces ATGL expression. 22 Inversely, activation of FoxO1 by SIRT1-induced deacetylation activates lipolysis by increasing ATGL expression. 23 However, the abundance of ATGL and HSL expression does not always correlate with cellular lipolytic activity. 24 Thus, cellular lipase expressions are inadequate as indicators of enzyme activities. In fact, IP extract used at 1.5 mg/mL decreases the expression of PPARγ, ATGL, and HSL expressions.
HSL is normally present in the cytoplasm. 25 In this study, HSL was phosphorylated by PKA at three residues (Ser563, Ser659, and Ser660) or by ERK1/2 at one residue (Ser600), after which it localizes to lipid droplets. 9,26 After localization to lipid droplets, the hydrolytic activity of HSL on TAG and DAG increases. 2 ERK1/2 is a serine/threonine kinase that is stimulated by cytokines such as insulin and tumor necrosis factor α (TNFα), growth factors, oxidative stress, and other factors, and it is activated through MEK. 27 H89, which was used in this study, selectively suppresses PKA, while U0126 directly inhibits MEK activity. Our results show that although the IP extract has a negative effect on the expression of HSL, it has no effect on the phosphorylation and activation of perilipin and HSL at Ser563 and Ser660. In agreement with this finding, H89 did not attenuate IP extract-induced lipolysis. Between the inhibitors used, only U0126 had the capability to abolish IP extract-enhanced lipolysis; IP extract enhanced the phosphorylation of ERK1/2 in a concentration-dependent manner, and this phosphorylation was significantly attenuated by U0126.
HSL activity can also be affected by inhibitory pathways. The Ser656 residue of HSL can be phosphorylated by AMP-activated protein kinase (AMPK), which inhibits HSL activity. 25 Previous studies have demonstrated the existence of cytokines, such as TNFα, resistin, interleukin 6, leptin, and ciliary neurotrophic factor, that inhibit as well as activate AMPK, and the regulation of lipolysis by these cytokines was the focus of a previous study. 28 AMPK is also an important factor in the regulation of lipolysis, and it would be interesting to determine whether IP is involved in regulating AMPK activity.
Interestingly, a recent report has demonstrated that mTORC1 is also associated with lipolysis. 22 Ras homolog enriched in brain (Rheb) acts on mTOR as a positive regulatory factor. 3T3-L1 cells that have undergone transformation to enable the stable expression of Rheb release smaller amounts of glycerol. The mTOR inhibitor rapamycin enhances TAG hydrolysis and fatty acid release in 3T3-L1 cells. 29 Another study demonstrated the positive effect of Trans-10, cis-12 CLA on lipolysis through convergent increase of mTOR and ERK1/2 activation in human adipocytes. 30,31 Recently, it was reported that the control of ERK1/2 phosphorylation is closely related to the Ca2+/calmodulin-dependent protein kinases II (CaMKII), and inhibition of CaMKII blunted ERK1/2 phosphorylation and glycerol release. 32 It is possible that IP extract, which acts to suppress adipocyte differentiation and enhance lipolysis, is involved in the mTOR signaling pathway or in CaMKII phosphorylation. We are currently investigating the mechanism of action of IP extract on the mTOR and CaMKII signaling pathways.
In this study, we showed that IP extract acts on differentiated mature adipocytes to enhance the hydrolysis of lipids. As mentioned above, IP has been found to contain a large amount of pinitol, myo-inositol, as well as antioxidant compounds. Two independent studies related to lipid metabolism demonstrated that pinitol has no effect on adipogenesis in 3T3-L1 preadipocytes, whereas, myo-inositol increased lipid accumulation and reduced basal lipolysis in the same cell line. 33,34 For this reason, future studies are needed to isolate and identify the functional substances endowed with antiadipogenic and lipolytic effects of IP extract, as well as in vivo analysis of their bioavailability. It is possible to expand on the results of this study and examine the use of IP as an effective functional food to prevent and treat metabolic syndrome.
Footnotes
Acknowledgment
This work was supported by a grant for Science and Education from the University of Tsukuba.
Author Disclosure Statement
No competing financial interests exist.