
Abstract
The dopamine transporter (DAT) protein, a component of the dopamine system, undergoes adaptive neurobiological changes from drug abuse. Prevention of relapse and reduction of withdrawal symptoms are still the major limitations in the current pharmacological treatments of drug addiction. The present study aimed to investigate the effects of essential oils extracted from Elsholtzia ciliata, Shinchim, Angelicae gigantis Radix, and Eugenia caryophyllata, well-known traditional Korean medicines for addiction, on the modulation of dopamine system in amphetamine-treated cells and to explore the possible mechanism underlying its therapeutic effect. The potential cytotoxic effect of essential oils was evaluated in PC12 rat pheochromocytoma cells using cell viability assays. Quantification of DAT, p-CREB, p-MAPK, and p-Akt was done by immunoblotting. DAT was significantly reduced in cells treated with 50 μM of amphetamine in a time-dependent manner. No significant toxicity of essential oils from Elsholtzia ciliata and Shinchim was observed at doses of 10, 25, and 50 μg/mL. However, essential oils from A. gigantis Radix at a dose of 100 μg/mL and E. caryophyllata at doses of 50 and 100 μg/mL showed cytotoxicity. Treatment with GBR 12909, a highly selective DAT inhibitor, significantly increased DAT expression compared with that of amphetamine only by enhancing phosphorylation of mitogen-activated protein kinases (MAPK) and Akt. In addition, essential oils effectively induced hyperphosphorylation of cyclic-AMP response element-binding protein (CREB), MAPK, and Akt, which resulted in DAT upregulation. Our study implies that the essential oils may rehabilitate brain dopamine function through increased DAT availability in abstinent former drug users.
Introduction
D
Dopaminergic neurons and their projections have been implicated as a key player in addiction. 4 Acute heroin administration elevates dopamine (DA) in the nucleus accumbens, 5 while chronic opioid users demonstrated downregulation of dopamine transporter (DAT) 4,6 and DA receptors, 7 including decreased dopaminergic activity with impairment of tyrosine hydroxylase. 8
DAT protein, as a component of the dopamine nerve terminal, is located primarily on the presynaptic membranes of dopaminergic neurons and modulates the reuptake of dopamine from the synaptic cleft. 4 There is considerable evidence for the DA system to have adaptive neurobiological changes from long-term drug abuse; decreased DAT levels were caused by the chronic use of methamphetamine 9 –11 and morphine. 12 Even imaging methods have been used to confirm the changes of DAT function in rhesus monkeys during acute morphine injection 13 and to assess DAT uptake function in heroin users. 14 Collectively, these findings are consistent with the hypothesis suggesting that an effective treatment for drug addiction may be achieved if the DA function is restored.
Transcription factors such as cyclic-AMP response element-binding protein (CREB) that bind to regulatory regions of genes and thereby increase or decrease the transcription of those genes have been implicated in mediating drug addiction at the gene expression level in the brain. 1 Addiction to a psychostimulant such as methamphetamine is accompanied by stimulation of DA receptors through transcriptional effects of psychostimulants, followed by increases in protein kinase A activity, phosphorylation of CREB (p-CREB), and subsequent CREB-mediated transcription. 15 CREB is regulated by several pathways, including protein kinase B (PKB/Akt). Thus, it is possible that the cellular signalling pathways through which drugs of abuse activate a given transcription factor in brain can be causally linked to such activation of that transcription factor's target genes and to specific behavioral aspects of addiction. 1 One such group of signaling pathways involved in drug addiction from the cellular stress response includes the family of mitogen-activated protein kinases (MAPK). Addiction-related behaviors were suppressed when MAPK isoforms were deleted using a gene knockout approach, 16 implying that mechanisms underlying increased risk for drug addiction might involve MAPK signal transduction pathways.
There are some medications for the treatment of drug addiction that target the alleviation of withdrawal symptoms, craving, and relapse. However, pharmacological treatments that are aimed at reducing the severity of withdrawal symptoms are opiates with undesirable side effects such as abuse potential. Moreover, there is not much choice of drugs that can reduce the drug craving and prevent relapse. Considering the side effects of synthetic medications, natural plants can be alternative sources of new antiaddiction medications due to their more tolerable and less toxic properties. Therefore, the current study was conducted to search for reliable alternatives to treat addiction.
Three different ingredients of traditional Korean medicine for addiction were selected to observe their effects in cell culture. Elsholtzia ciliata Hylander is a valuable bioactive source of natural antioxidants. 17 In particular, the water extract of E. ciliata is found to possess anti-inflammatory properties, which are MAPK dependent. 18 Angelicae gigantis Radix has traditionally been used for inflammatory diseases. An ethanol extract of the root of A. gigantis showed considerable antiallergic activity on human mast cells. 19 Reported pharmacological activities of A. gigantis Radix include inhibitory effects on osteoclast formation, 20 angiogenesis, 21 and immune-enhancing effects. 22 Eugenol isolated from the cortex of Eugenia caryophyllata Thunb. is found to possess anti-inflammatory and anticancer properties by suppressing cyclooxygenase and subsequent prostaglandin production. 23
These three natural plants have been used as traditional herbal medicines in the treatment of neurological disorders in Korea. However, few studies have evaluated their potential as antiaddiction medications. The effects of these herbs on p-CREB, p-MAPK, and p-Akt are currently unknown. In this experiment, rat pheochromocytoma (PC12) cell line was used to investigate the hypothesis that modulation of the dopamine system by herbs is a possible mechanism of their therapeutic actions.
Materials and Methods
Preparation of the essential oil
All medicinal plants were purchased from the Omni Herb Company (Seoul, Korea). An essential oil was prepared in the same way as described previously. 24 One gram each of E. ciliata, A. gigantis Radix, and E. caryophyllata was pulverized and extracted once with 2 L of n-hexane at room temperature for 48 h and then filtered. The filtrate was evaporated at 80°C to remove hexane, which was further eliminated under vacuum for 5 min at room temperature to give a clear, pale brown essential oil. An additional special essential oil called Shinchim was prepared by using 28 medicinal herbs (Table 1). Each extract was solubilized in ethanol and filtered through a 0.2-μm filter for cell culture usage. The use of Shinchim as a therapeutic agent for various cerebrovascular diseases has appeared in Dongui Bogam, a Korean medical book first published in 1613 and one of the classics of oriental medicine today. According to Dongui Bogam, Shinchim can be used for Qi-tonifying, phlegm dispersing, and activation of blood circulation which is an important principle for treatment of brain-associated diseases.
UPLC-ESI-MS analysis
In the current study, an ultraperformance liquid chromatography coupled with electrospray ionization mass spectrometry (UPLC-ESI-MS) method was applied for investigating the pharmacochemistry of each extract. Decusin (Sigma, St. Louis, MO, USA) and eugenol solutions were prepared by dissolving sample (10 μg) in methanol (1 mL), and each essential oil (50 μg) was dissolved in ethanol (1 mL). The UPLC system (Waters Corporation, Milford, MA, USA) was equipped with a quaternary pump, vacuum degasser, and a diode-array detector. The chromatographic condition was as follows: UPLC™ BEH C18 column (1.7 μm, 2.1 × 50 mm); mobile phase: a linear gradient system of
Cell culture
PC12 cells were purchased from American Type Culture Collection (Rockville, MD, USA). Cells were cultured in the RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% horse serum (HS; Invitrogen), 5% fetal bovine serum (Invitrogen), 100 unit/mL penicillin, and 100 μg/mL streptomycin (Gibco-BRL, Rockville, MD, USA) with further addition of trypsin-EDTA (Gibco-BRL). The cells were maintained at 37°C in 5% CO2 and plated at a density of 1 × 106 cells/well on poly-L-lysine-coated six-well plates (Corning Costar Corp., Cambridge, MA, USA). Cells were pretreated with the indicated concentrations of each essential oil for 30 min before exposure to amphetamine (50 μM) or GBR 12909 (Tocris Bioscience, Ellisville, MO, USA) for 24 h.
Cell viability assay
Cell viability was determined by the 3-(4,5-dimethylthizaol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Abcam, Cambridge, MA, USA) assay. PC12 cells were seeded in 12-well plates at a density of 5 × 103 cells/well, stabilized at 37°C for 16 h in the RPMI 1640 medium supplemented with 0.5% HS, and cultured with the indicated concentrations of essential oils for 24 h. The medium was removed and the cells were incubated with 2 mg/mL of MTT solution. After incubation for 4 h at 37°C and 5% CO2, the supernatants were removed and dimethyl sulfoxide (Sigma) was added. The reactants were measured in terms of optical density (OD) at 580 nm with a microplate reader (UV max; Molecular Devices, Sunnyvale, CA, USA). The ODs were converted into percentages using the following formula: Cell viability (%) = OD sample/OD negative control × 100. Negative control cells were treated with RPMI 1640 alone.
Quantification of DAT, p-CREB, p-MAPK, and p-Akt by Western blot analysis
PC12 cells were maintained in a serum-free medium with essential oils at 30 μg/mL for 24 h. Cells were then washed twice with ice-cold phosphate-buffered saline and lysed in a lysis buffer containing 62.5 mM Tris-HCl, pH 6.8, 2% sodium dodecyl sulfate, 20% glycerol, 10% 2-mercaptoethanol, and protease inhibitors. After incubation for 30 min on ice, cell lysates were centrifuged and protein concentrations were determined using the bicinchoninic acid method (Pierce, Rockford, IL, USA). Samples of cell lysates containing 30 μg of total protein were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes (Amersham Pharmacia Biotech, Buckinghamshire, United Kingdom) by electroblotting. After blocking with 5% skim milk in Tris-buffered saline (50 mM Tris-HCl, pH 7.6, 150 mM NaCl, 0.1% Tween-20) for more than 30 min, the membranes were incubated for 16 h at 4°C with primary antibodies (anti-DAT, anti-p-CREB, anti-p-MAPK, anti-p-Akt, and β-actin) and further incubated with horseradish peroxidase-conjugated secondary antibodies for 1 h. The membranes were visualized by an enhanced chemiluminescence system (ECL kit; Pierce). Band images were obtained by using Molecular Imager ChemiDoc XRS+ (Bio-Rad Laboratories, Hercules, CA, USA) and the band intensity was analyzed by Image Lab™ software version 2.0.1 (Bio-Rad).
Data analyses
All statistical analyses were conducted with SPSS (ver. 19; SPSS, Inc., Chicago, IL, USA). Values are expressed as mean ± SD. Data were analyzed by one-way ANOVA and then differences among means were analyzed using Tukey–Kramer's multiple comparison test. Statistical significance was accepted at a P-value < .05.
Results
Establishment of UPLC profile and identification of phytochemicals by MS
To establish the standard chromatogram for each extract, we adopted the UPLC system. Gradient elution systems are the way of choice to obtain entire chromatographic profiles of the phytochemicals from natural resources. The identification of major peaks of chromatogram was performed by UPLC-ESI-MS. The identified peaks of each extract were recorded in Figure 1: piperine and pipericine from Shinchim, decursin from A. gigantis, eugenol from E. caryophyllata, or elsholziaketone from E. ciliata.
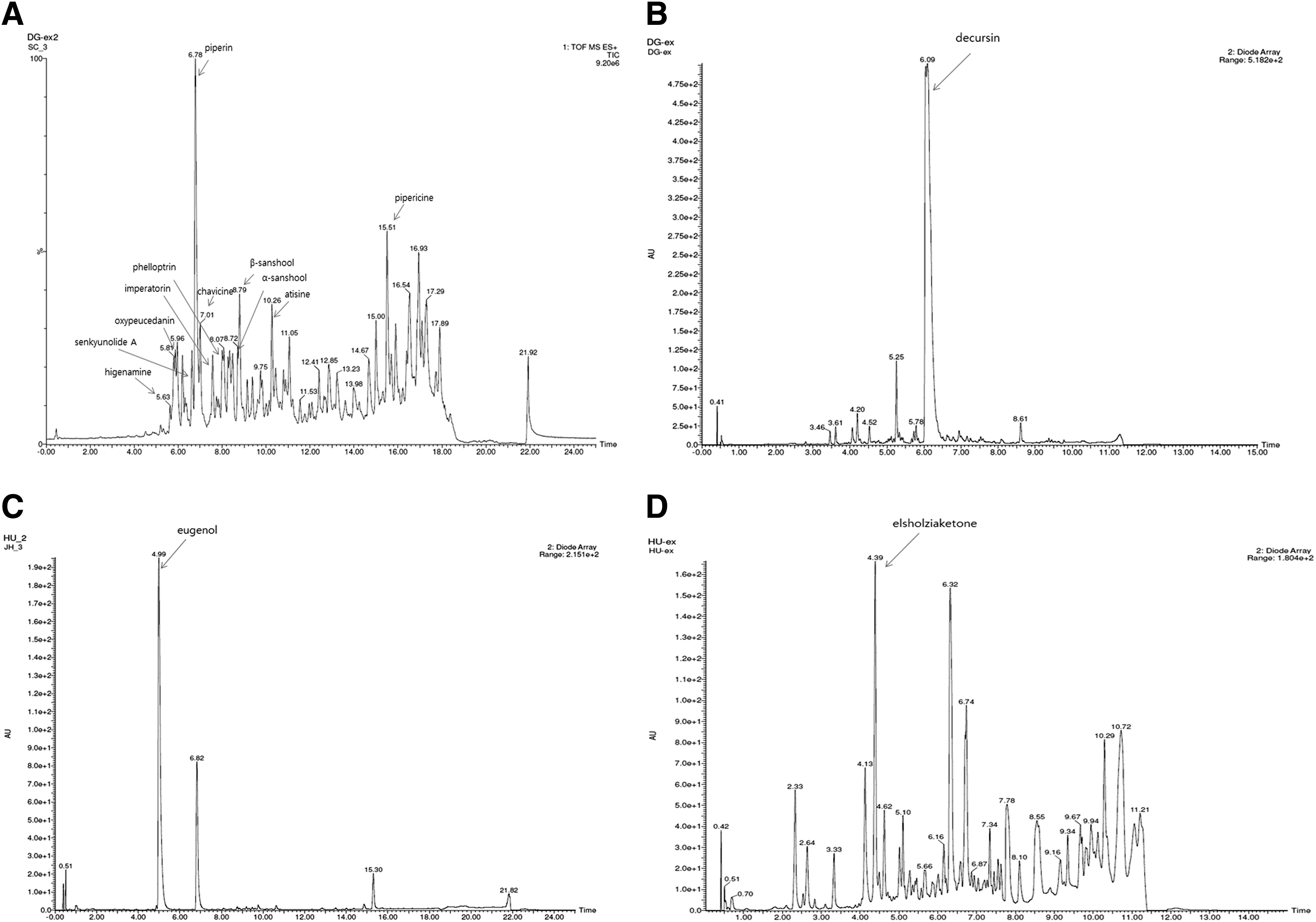
UPLC chromatogram and ESI-MS spectra of peaks from each extract.
Amphetamine decreases DAT expression
We analyzed the expression level of DAT by Western blot. The DAT level was standardized with β-actin as a loading control. DAT was significantly lower in cells treated with 50 μM of amphetamine in a time-dependent manner (Fig. 2).

Amphetamine decreases dopamine transporter (DAT) expression level in PC12 cells. Serum-starved PC12 cells were stimulated with 50 μM of amphetamine for 15, 60, or 240 min, and whole cell lysates were electrophoresed in sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and analyzed by immunoblotting with anti-DAT and β-actin antibodies. The intensity of bands was quantitated by densitometry from four independent experiments and the amounts of DAT were normalized versus β-actin. C, unstimulated control cells; **P < .01 versus control.
Effect of essential oils on cell viability
The potential cytotoxic effect of essential oils was investigated by determining their effects on the viability of PC12 cells. Cells were treated for 24 h with essential oils at concentrations of 10, 25, and 50 μg/mL of E. ciliata. For Shinchim, A. gigantis Radix, and E. caryophyllata, doses of 25, 50, and 100 μg/mL of essential oils were used (Fig. 3). No significant toxicities of essential oils from E. ciliata and Shinchim were observed at the indicated concentrations. However, essential oils from A. gigantis Radix at a dose of 100 μg/mL and E. caryophyllata at doses of 50 and 100 μg/mL were shown to elicit cytotoxicity. Accordingly, the doses of essential oils, which were overtly cytotoxic to the cells were not used for further investigation. Amphetamine at a dose of 50 μM was shown not to be toxic to PC12 cells.
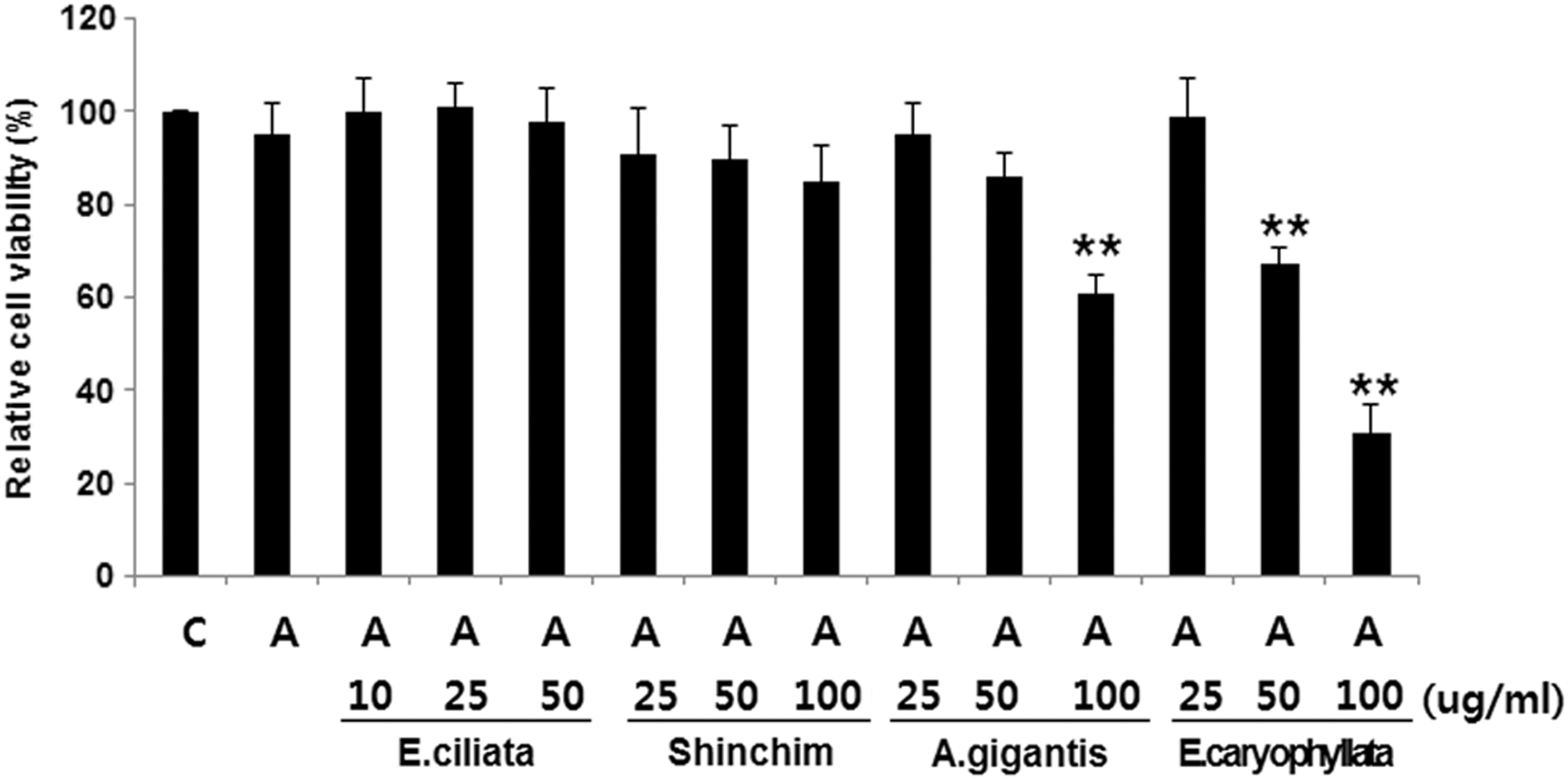
Dose-dependent cytotoxic effect of essential oils and amphetamine in PC12 cells. Serum-starved PC12 cells were stimulated with 50 μM of amphetamine and indicated concentrations of essential oils from E. ciliata, Shinchim, A. gigantis Radix, and E. caryophyllata. Cell viability was measured by the MTT assay. Data expressed as percentage of control are mean ± SD of six separate experiments. C, unstimulated control cells; and A, amphetamine-treated cells, *P < .05 versus A.
DAT inhibitor reverses DAT degradation and causes phosphorylation of MAPK and Akt
To investigate the effect of DAT inhibitor on the amphetamine-induced DAT degradation and phosphorylation of MAPK and Akt in PC12 cells, GBR 12909, a potent and highly selective DAT inhibitor, was utilized. The amphetamine-treated group exhibited a decreased DAT level by half compared to the control group, while treatment with either 5 or 10 μM of GBR 12909 increased DAT expression 2.4 and 2.8-fold, respectively, compared with that of amphetamine only (Fig. 4A). Both the sample immunoblot and the mean data obtained from densitometric analysis indicate that the activities of both kinase phosphorylations were enhanced in PC12 cells, irrespective of doses of GBR 12909 used (Fig. 4B). In the amphetamine-treated group, there was no change in the expression of p-MAPK or p-Akt compared with the control group.
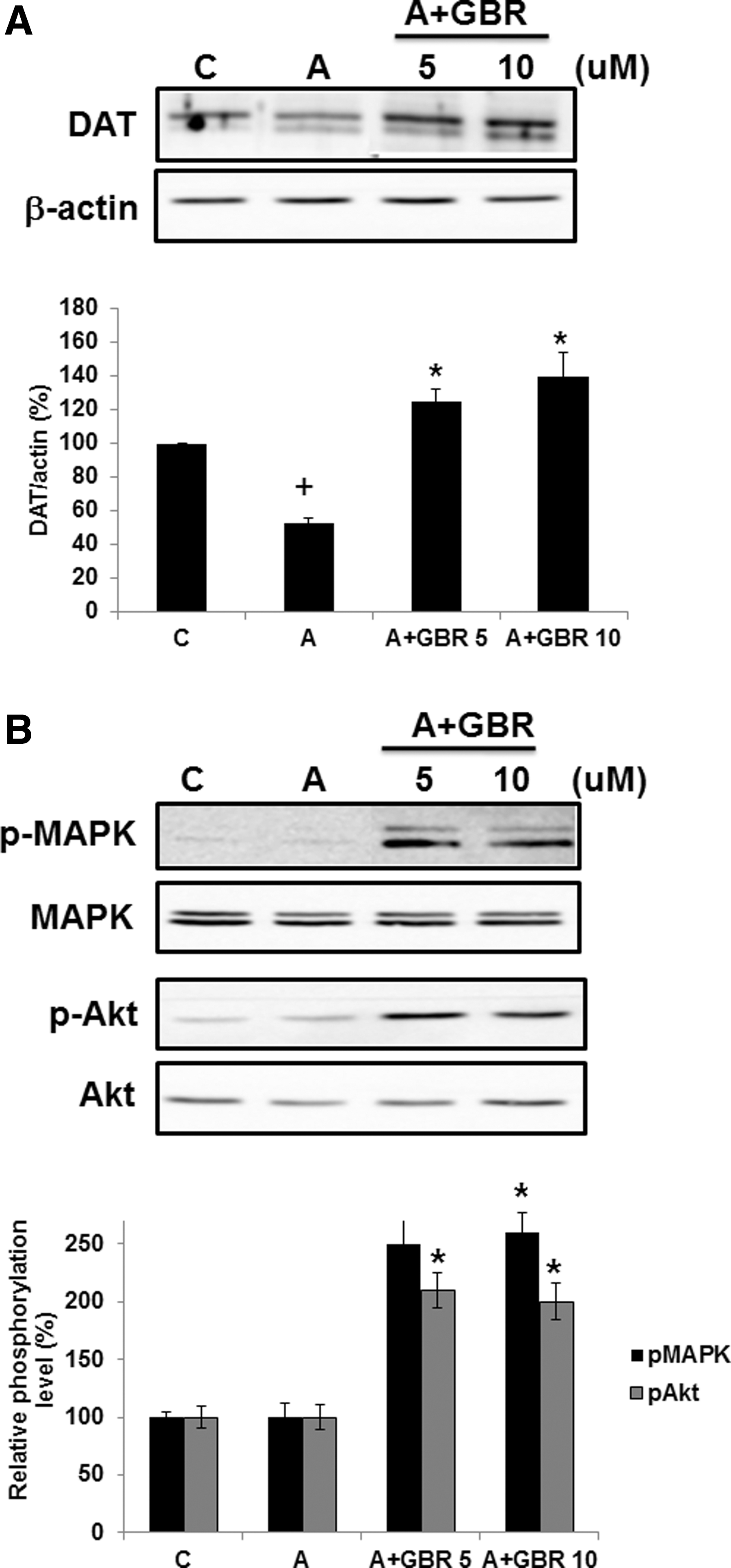
DAT inhibitor, GBR 12909, upregulates DAT expression
Essential oils induce increased DAT expression
The effects of essential oils on DAT expression in the presence of amphetamine are illustrated in Figure 5. We found that essential oil treatment increased DAT levels in PC12 cells in a dose-dependent manner compared to the amphetamine-treated control.
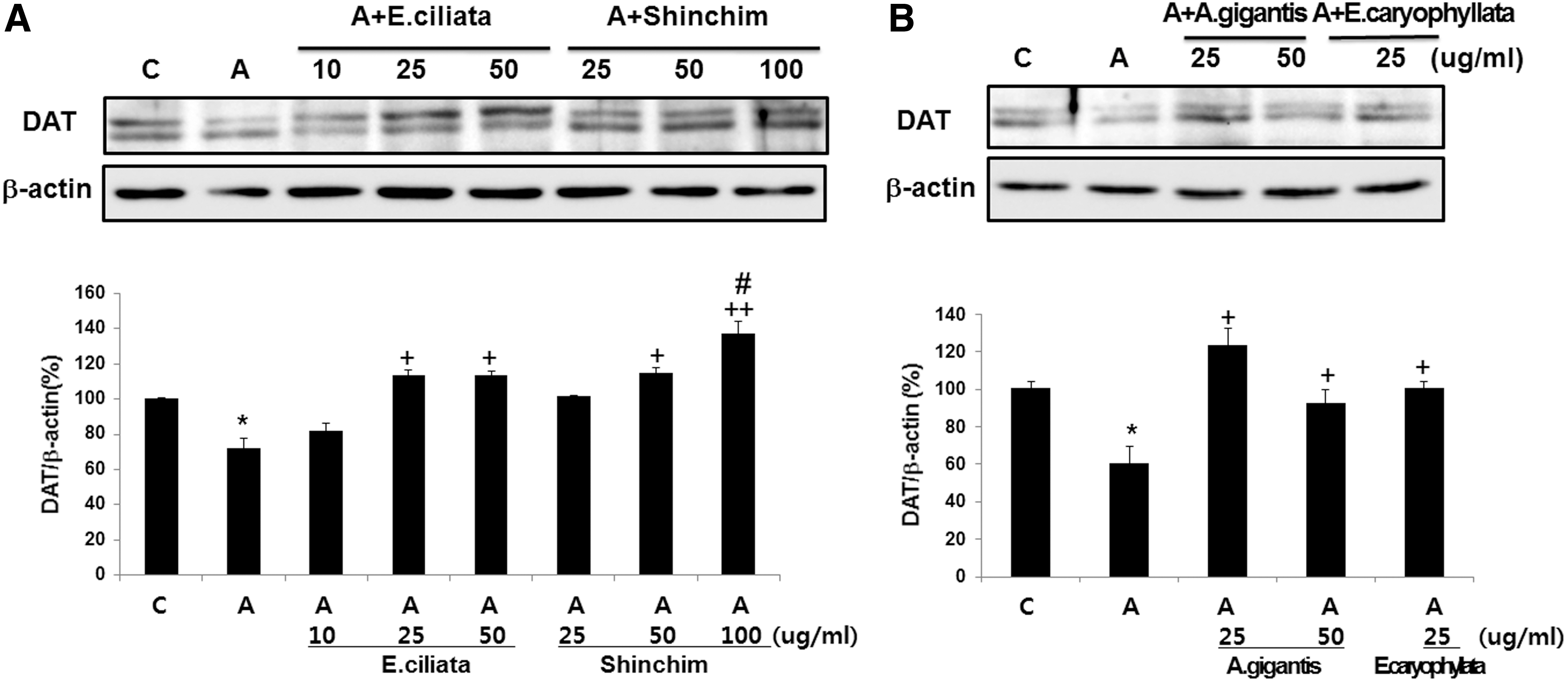
Essential oils induce an increase in DAT expression. Serum-starved PC12 cells were treated with 50 μM of amphetamine for 24 h and indicated doses of essential oils from E. ciliata, Shinchim
Essential oils induce phosphorylation of CREB, MAPK, and Akt
The effects of essential oils on phosphorylation of CREB, MAPK, and Akt in the presence of amphetamine are shown in Figure 6. No phosphorylation was observed from the administration of E. ciliata essential oil in PC12 cells. Treatment of Shinchim essential oil was accompanied by a significantly increased phosphorylation of both CREB and MAPK. Strong CREB phosphorylation was only observed in PC12 cells treated with A. gigantis Radix and E. caryophyllata.

Essential oils induce phosphorylation of cyclic-AMP response element-binding protein (CREB), mitogen-activated protein kinases (MAPK), and Akt. Serum-starved PC12 cells were treated with 50 μM of amphetamine for 24 h and indicated doses of essential oils from E. ciliata, Shinchim, A. gigantis Radix, and E. caryophyllata were added
Discussion
The present study demonstrates the modulation of the dopamine system as a mechanism underlying the therapeutic action of the essential oils extracted from medicinal plants in alleviating addiction by upregulation of DATs. This would explain the inverse relationship between DAT levels and addiction; as the number of reuptake proteins rises, the dopamine would decline, leading to a lower level of addiction. In addition, essential oils effectively induced hyperphosphorylation of CREB, MAPK, and Akt, implying that changes in the phosphorylation state into hyperphosphorylation exert a therapeutic effect on opioid addiction. Furthermore, amphetamine-induced DAT decrease was reversed by blocking the DAT with the selective inhibitor vanoxerine (GBR 12909), suggesting that a mechanism of such action is related to the enhancement of kinase activities of MAPK and Akt.
The restoration of the DA system is believed to be effective in the treatment of opioid addiction. Literature data indicate that repeated drug intake exerts a catalytic activity on dysregulated DA function. Decreased DAT level has been found in patients addicted to opioids 4,6,14,25 and methamphetamine. 9 –11 Chronic administration of opioids to rodents has caused a decrease in DAT, 12 DA, 26,27 tyrosine hydroxylase, 28 and DA receptors. 29 From these findings, we would speculate that opioid addiction can be treated by targeting pathologies of dopaminergic disorders. In this study, we examined the effects of administration with medicinal plant essential oils on the DAT level using an amphetamine-treated cell culture. Reduced DAT level in amphetamine-treated PC12 cells of the current study is in agreement with the findings of previous imaging studies in heroin-dependent patients 4,6,14,25 and in morphine-injected rhesus monkeys, 13 in which exposure to addictive substances was shown to be associated with a decrease in the striatal DAT level. However, a postmortem study 8 and another imaging study 30 found no difference in striatal DAT levels between heroin users and control subjects. This discrepancy might be due to the introduction of a detoxification protocol in the study design before the first imaging session 4,14,25 or not. 30 The subject numbers could be another explanation. Interestingly, those study findings that decreased DAT levels in heroin users tended to have a large sample size, whereas studies reporting no difference in DAT levels tended to have small number of enrolled subjects.
Together, the above studies suggest that a successful treatment of opioid addiction can be achieved through the reversal of reduced DAT levels. Furthermore, the development of effective opioid addiction treatment strategy should include reducing the intensity of withdrawal symptoms. The implementation of abstinence might affect the readiness of dopamine cells to respond in humans. In cocaine users, the DAT density increases significantly after acute cocaine discontinuation. 31 Both heroin users 4 and methamphetamine/methcathinone users 32 tend to show low level of DAT despite the long period of abstinence. This may imply that a decreased DAT availability in the brains of drug addicts is long lasting and does not readily recover during abstinence, although the final outcome of DAT levels can be varied across different types of drugs of abuse. It is, therefore, noteworthy that DAT degradation induced by amphetamine was reversed by DAT inhibitor in the current study. The recovery of DAT density by GBR 12909 was stimulated upon phosphorylation of two protein kinases, MAPK and Akt. Moreover, normalization of DAT levels with the treatment of medicinal plant essential oils is in accordance with prescription of 15 herbs to heroin-dependent subjects. 4 A different type of herbal formula restored a decreased DAT availability to normal levels in a rat model of morphine dependence. 33
Among several brain regions, the nucleus accumbens is seen as important for memory of positive emotional states where an altered functioning of nerve cells can lead to addiction. A large body of literature has shown that CREB in the nucleus accumbens is activated by repeated drug exposure through negative feedback system, whereby CREB is responsible for reducing the user's sensitivity to the rewarding effects of drugs. 1 Our study evaluated the signaling pathways underlying the effects of medicinal plant essential oils on pheochromocytoma cells. In this study, we show that the expression of CREB phosphorylation is upregulated through the MAPK/Akt signaling pathways. This provides new information about the inhibitory effect of amphetamine on essential oil-induced upregulation of the MAPK/Akt signaling pathways, which may underlie or contribute to defective CREB signaling.
Among the components of each extract, eugenol and decursin were identified (Fig. 1). Although eugenol and decursin have neuroprotective and antiapoptotic effects in the brain, 34,35 it has not been reported that they are effective interventions for drug addiction or dopamine regulation. Thus, both phytochemicals are worth studying for DAT regulation and addition.
In conclusion, the results of this study demonstrate that DAT availability is lower in amphetamine-treated cells compared to controls. We showed that DAT upregulation was induced by medicinal plant essential oils. Importantly, we documented that the increases in DAT from the treatment of essential oils are mediated through the hyperphosphorylation of CREB, MAPK, and Akt. This is an important result, because it implies that the essential oils may rehabilitate brain DA function through increased DAT availability in abstinent drug users. Although more data are required to pinpoint the exact mechanism underlying this effect of essential oils on the DA system, our study nonetheless provides biological evidence of the efficacy of medicinal plant essential oils in the treatment of drug addiction.
Footnotes
Acknowledgment
This work was supported by the Dongguk University Research Fund of 2012.
Author Disclosure Statement
No competing financial interests exist.