
Abstract
This study investigated the effects of mangosteen on metabolic syndromes in high-fat (HF) diet-fed mice and the underlying mechanisms related to adipogenesis. Mangosteen-supplemented mice gained significantly less body weight, compared with the HF group. The levels were markedly elevated in HF mice for serum glutamate oxaloacetate transaminase, glutamate pyruvate transaminase, glucose, triglyceride, total cholesterol, low-density lipoprotein (LDL) cholesterol, and free fatty acid; whereas these levels were significantly lower in the 200 mg/kg of the mangosteen extract-treated group. The mangosteen extract did not modify high-density lipoprotein (HDL)-cholesterol, however, LDL-cholesterol was lower and HDL/LDL ratio was higher (9.4 vs. 3.7 in HF group). Furthermore, 200 mg/kg of mangosteen treatment activated the hepatic AMP-activated protein kinase and Sirtuin 1 in an in vivo system. Thus, the results of this study suggest that mangosteen extract exerts antiobesity effects by regulating energy metabolism and hepatic lipid homeostasis.
Introduction
T
Mangosteen (Garcinia mangostana L.) is a tropical fruit that contains xanthones, terpenes, anthocyanins, tannins, phenols, and multiple vitamins as biologically active compounds. 6 Numerous studies have been conducted regarding the various health benefits of mangosteen such as antioxidative, antibacteria, anticancer, and antiadipogenic effects. 7 –11 Also, an herbal formulation containing G. mangostana was reported to help weight management. 12 However, the antiobesity effect of mangosteen per se on mice fed a HF diet remains unknown. Thus, in this study we investigated the effects of mangosteen ethanol extract (M) on metabolic disorder states in HF diet-fed mice and the underlying mechanisms.
Materials and Methods
Extraction and isolation of test material
Peels of G. mangostana were obtained from a local store in Phuket, Thailand, in June 2012. These plant samples were identified by one of the authors (Dr. J.H. Paik). Raw material used in this study (CYWDU-CP002) has been deposited at the College of Pharmacy at Dongguk University, Seoul, Republic of Korea, and it was extracted with ethanol followed by evaporation and lyophilization. The quantitative analysis of α- and γ-mangostins was conducted, in accordance with a reported method; 25.7% of α-mangostin and 3.8% of γ-mangostin were present in M, as shown in the Supplementary Table S1 (Supplementary Data are available at
Reagents
Polyethylene glycol 400 (PEG 400) was procured from Showa Chemical Company (Tokyo, Japan). Phosphorylated AMPK (p-AMPK), AMPK, SirT1, PPARγ, β-actin, and CPT1a antibodies were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Animals
The protocols for the animal studies were approved by the Institute of Laboratory Animal Resources of Dongguk University, Seoul, South Korea (# 2012-0673; July 25, 2012). Male C57BL/6 mice were obtained from Charles River Orient (Seoul, South Korea) and acclimated for 1 week before starting the study. Upon arrival, the animals were randomized and housed at three per cage under strictly controlled environmental conditions (20–25°C and 48–52% relative humidity). A 12-h light/dark cycle was used at an intensity of 150–300 lux. The C57BL/6 mice were started on either a normal diet (ND, 10% of kilocalories as fat; Product N12450B), or HF (60% of kilocalories as fat; Product D12592, Research Diets, New Brunswick, NJ, USA) for 80 days. After 5 weeks (at day 35) of ND or HF, the mice were distributed into six treatment groups orally administered with either 50 or 200 mg/kg/day of M (HFM50 and HFM200 groups, respectively) or orlistat (50 mg/kg/day; HFO group) dissolved in PEG 400 and distilled water (6:4, v/v) to the mice with HF every day during the last 45 days of feeding. The number of mice in each group was eight.
Hepatic triglyceride measurement
The samples of mouse liver (0.3 g) were homogenized in 0.1 M Tris-acetate buffer (pH 7.4). And then, six volumes of chloroform: methanol (2:1) were added. After vigorous stirring, the mixtures were incubated on ice for 1 h and then centrifuged at 800 g for 3 min. The resulting lower phase was aspirated. The serum triglyceride (TG) content was ascertained using a Sigma-Aldrich Kit. 14
Blood chemistry
A serum sample was collected to measure total protein, albumin, urea nitrogen, glutamate oxaloacetate transaminase (GOT), glutamate pyruvate transaminase (GPT), creatinine, total cholesterol (TC), high-density lipoprotein (HDL) cholesterol, low-density lipoprotein (LDL) cholesterol, TG, free fatty acid (FFA), and glucose; the analysis was carried out at the Green Cross Reference Laboratory (Seoul, South Korea).
Immunoblot analysis
Protein expression was assessed by western blot analysis, using a kit in accordance with the manufacturer's instructions. Protein concentration in liver was determined using the BIO-RAPID protein assay reagent. There was a 1:1 mixing of 20 μg of protein, from liver, with 2× sample buffer (20% glycerol, 4% sodium dodecyl sulfate (SDS), 10% 2-mercaptoethanol, 0.05% bromophenol blue, and 1.25 M Tris-buffer of pH 6.8), which was loaded on to an 8% or 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel, and run at 150 V for 90 min. Tissue proteins were transferred to an immunoblot polyvinylidene difluoride membrane with a Bio-Rad semidry transfer system. The polyvinylidene difluoride membrane was then incubated with primary antibody (dilutions 1:500–1:1000) with 0.1% Tween 20 in Tris-buffered saline overnight. The blots were washed thrice with Tris-buffer saline containing 0.1% Tween 20 and incubated for 1 h with HRP-conjugated secondary anti-IgG antibody (dilution 1:2000–1:20 000). The blots were washed again thrice with 0.1% Tween 20 in Tris-buffered saline, and immunoreactive bands were developed, using the chemiluminescent substrate (Amersham Biosciences, Piscataway, NJ, USA).
Statistical analysis
A P value < .05 was deemed to be statistically significant using Duncan's multiple range test of the Social Package of Statistical Sciences (SPSS) a posteriori analysis of variance. All data are expressed as mean ± standard deviation.
Results
Effects of M on obesity in in vivo model
The body weight gain of the HF group was significantly higher than that of the ND group from day 14 to the end of this experiment; the body weight gains compared with the initial body weight: 10.3% vs. 18.5% at day 14, and 36.6% and 89.2% at day 80, respectively (Fig. 1). Also the glucose, TG, TC, LDL, and FFA were distinctly higher (60.1%, 160%, 38.8%, 103%, and 58.6%, respectively), but there were no significant difference in HDL levels between the ND and HF groups (Fig. 2). The GPT and GOT levels, which are markers of hepatic function, were elevated at 133% and 168%, respectively, in the HF group versus the ND group (Fig. 2); however, kidney function, based on the serum creatinine level and kidney weight, was identical to that of the ND group.
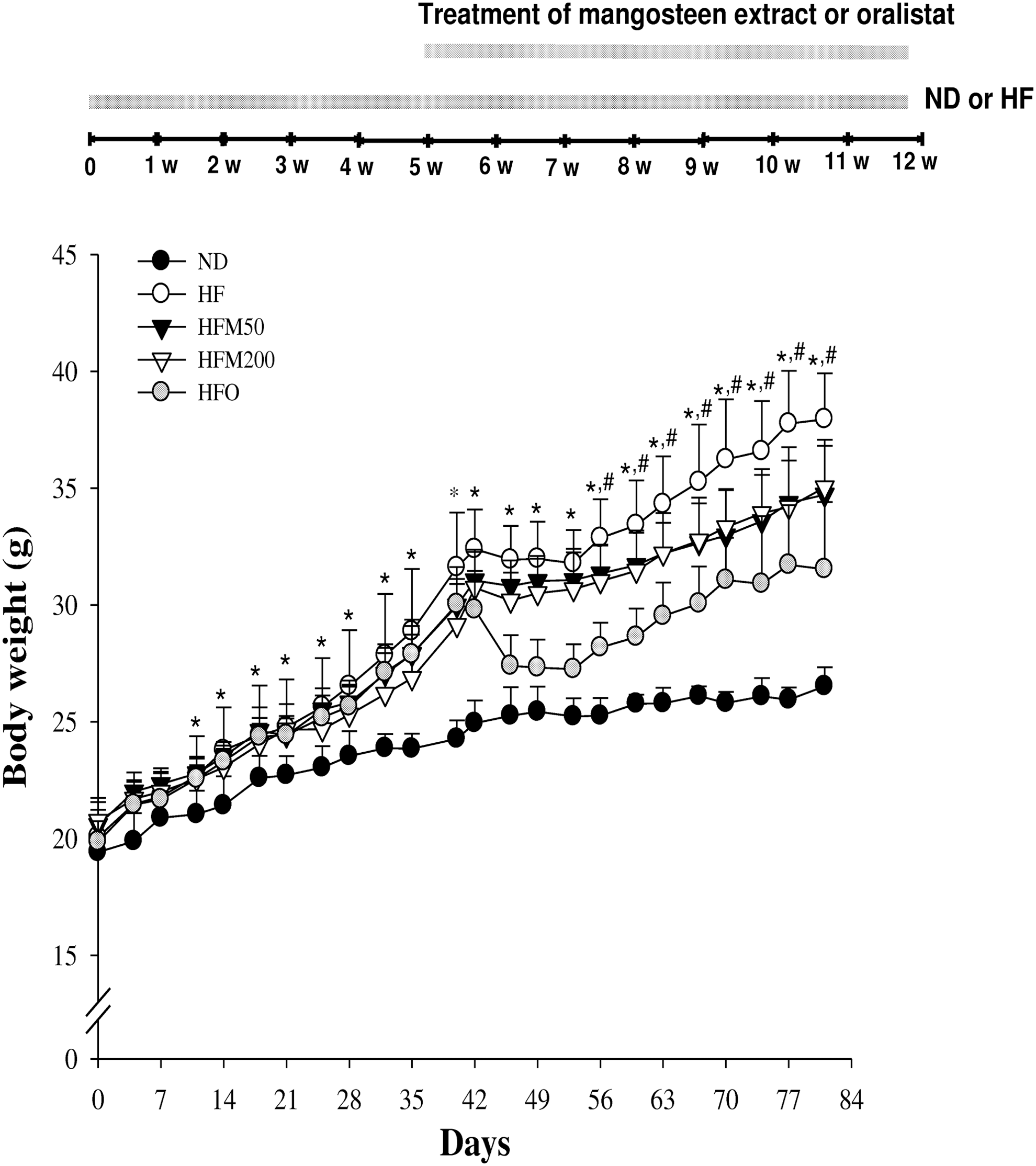
Effects of mangosteen ethanol extract (M) on body weight gain in ND, HF, HFM50, HFM200, and HFO groups. Statistical significance of differences between each treatment group and ND (*P < .05) or HF groups (# P < .05) were determined. Between HFM50 and HFM200, the statistical significance of difference was not significant. HF, high-fat; ND, normal diet.
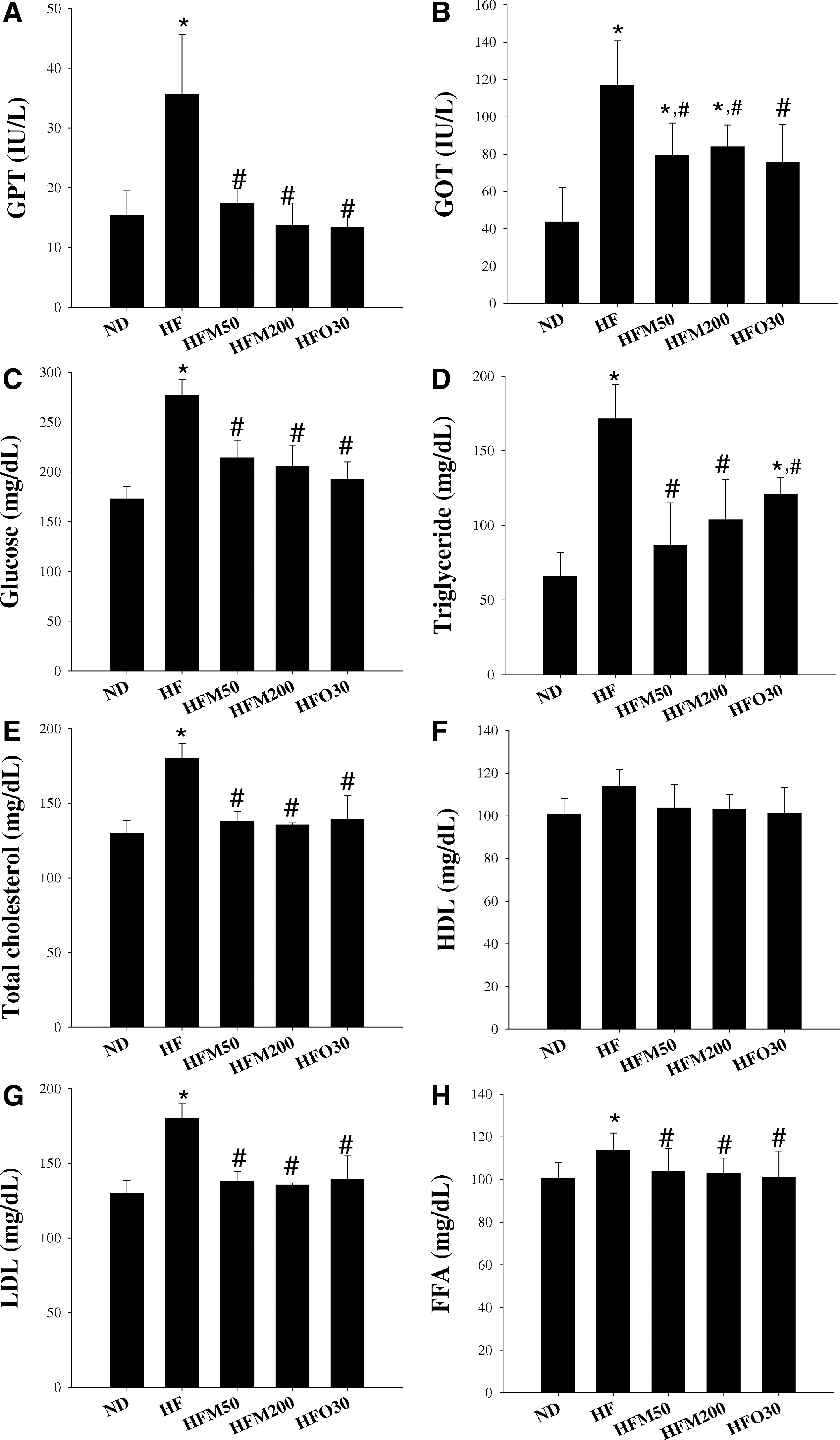
Effects of M on serum profiles in ND, HF, HFM50, HFM200, and HFO groups. At the end of the experimental period, GPT
Mice, given a HF diet for 80 days, exhibited marked increase in liver weight (by 34.8%) and increase of epididymal, inguinal, mesenteric, and retroperitoneal fats (by 407%, 423%, 232%, and 456%, respectively; Fig. 3) while HF-fed mice supplemented with 50 or 200 mg/kg/day of M for 5 weeks (from day 35 to day 80) (HFM50 and HFM200 groups) had significantly lower body weight and liver weight (Figs. 1 –3). With regard to the body fat, epididymal was significantly lower (by 22.4% and 20.0% for HFM50 and HFM200 groups, respectively, vs. HF group), inguinal (by 31.2% and 39.0%, respectively), and mesenteric (by 22.5% and 43.9%, respectively) fats were distinct at day 80, but the retroperitoneal fat was still unchanged (Fig. 3). To determine whether or not M improves lipid metabolism, the metabolic parameters representing serum concentrations of TG, TC, HDL, LDL, and FFA were measured (Fig. 3). Compared with the HF group, TG (by 49.6% and 39.5%, respectively), LDL (by 46.8% and 63.9%, respectively), and TC (by 23.3% and 24.8%, respectively) were significantly lower in the HFM50 and HFM200 groups. The HDL levels in all five groups were still unchanged, regardless of ND or HF diet and treatment by M or orlistat (O). There was no difference in the amount of food intake among these groups (Fig. 2).

Effects of M on food intake
Effects of monocot grass extract supplementation on hepatic fatty acid metabolism in HF-fed mice
To investigate how M controls in vivo lipid metabolism, western blot analysis was performed. The M and O effectively activated the AMPK and SirT1, both of which facilitated the reduction of body weight in obese mice. Moreover, the PPARγ levels were reduced in HFM 200 groups, which suggested the decrease of lipogenesis in adipocyte differentiation. Subsequently, the effects of M treatment on the levels of lipogenic gene transcripts were determined in the liver of HF-fed mice. In parallel with the inhibition of visceral fat accumulation by M, the increased PPARγ was suppressed by M treatment in HFM200 group (Fig. 4). Overall, M had a bona fide antiobesity effect in the HF-fed mice, which was accompanied by the inhibition of PPARγ and induction of AMPK and SirT1. In this study, the CPT1a decreased in the HF group (vs. ND group), but its level was significantly increased in the HFM200 group, compared with the HF group.
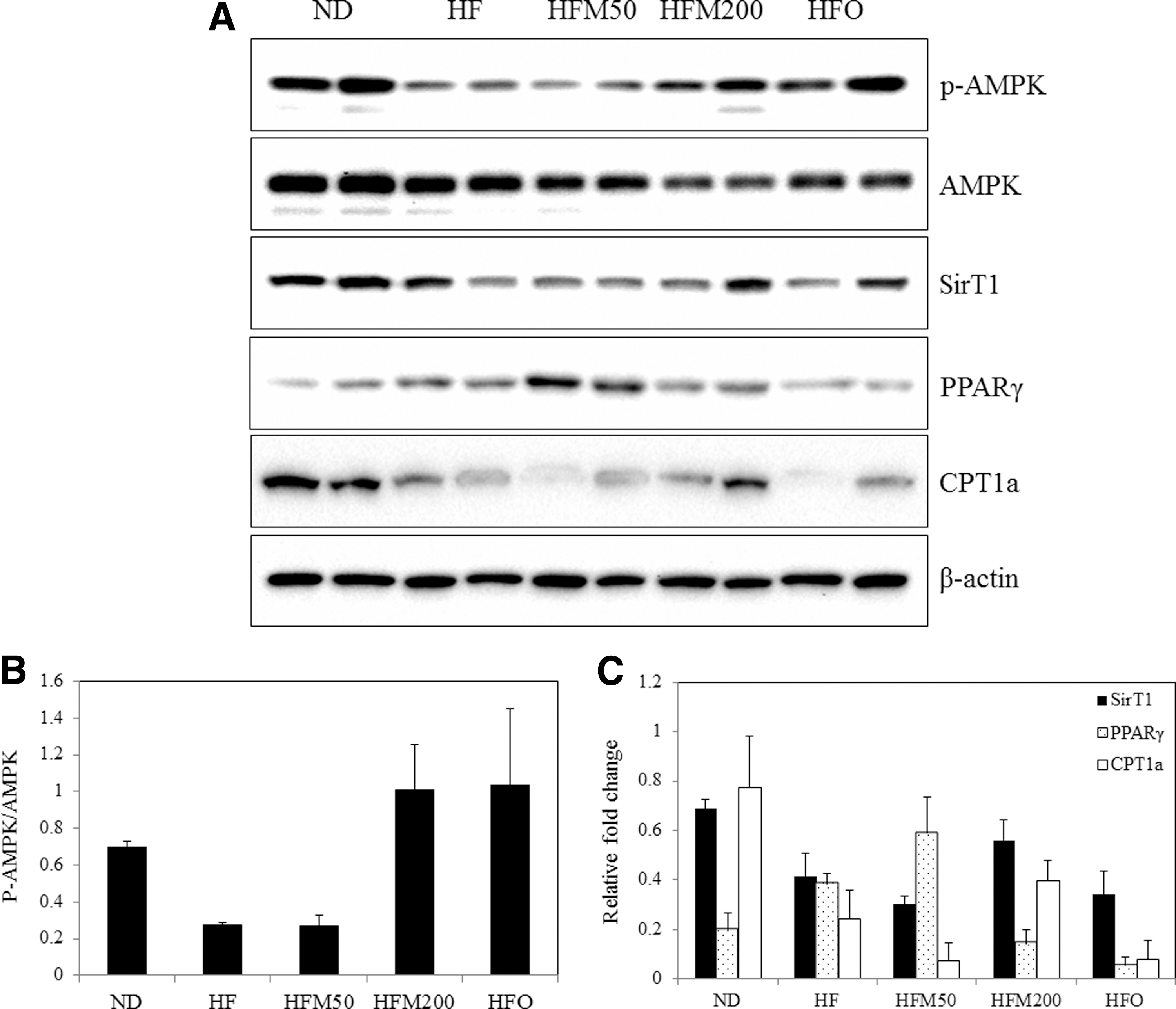
The AMPK, SirT1, PPARγ, and CTP1a pathways regulate energy metabolism and their relationship with hepatic lipid homeostasis.
Discussion
A metabolic syndrome is defined as one with symptoms of abdominal obesity, dyslipidemia, impaired glucose metabolism, hypertension, and inflammatory responses. 15,16 In particular, dyslipidemia is caused by an unhealthy diet or lifestyle, or both, such as diets rich in carbohydrate, fat, and cholesterol, or being overweight. 17,18 This study provides the first direct evidence of the antiobesity effects of M. These results suggest that M exerts beneficial antiobesity effects by modulating the lipid metabolism pathways. Moreover, the elevated levels of TC and LDL are associated with an increased risk of coronary heart disease. 19 Despite the reduced LDL/HDL ratio as a risk factor for coronary heart disease, the HF mice showed increasing levels of serum TC, HDL, LDL, and TG, which increased less throughout the treatment period in M-treated HF mice (Fig. 2). Also, it was found that M suppressed visceral fat accumulation in HF-induced obese mice (Fig. 2).
AMPK is the downstream component of a protein kinase cascade that acts as an intracellular energy sensor, thereby maintaining the energy balance within a cell. This pivotal role of AMPK makes it an ideal candidate for regulating whole-body energy metabolism, and additionally AMPK might play a part in protecting the body from metabolic diseases, such as type 2 diabetes and obesity 20 AMPK is a pivotal control of oxidation for regulating glucose and lipid metabolism. 21 That enzyme functions as a kind of fuel gauge: it is activated under conditions of high-energy depletion, such as hypoxia and ischemia. 22 PPARγ forms heterodimers with the retinoid X receptor that regulate genes involved in lipid metabolism and insulin sensitization. 23,24 These transcription factors interact with coactivators from the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α). 25 PGC-1α is a substrate for AMPK phosphorylation, leading to its activation, even though the manner by which this phosphorylation activates PGC-1α remains elusive. 26 SirT1, the mammalian orthologue of Sir2, is a nuclear protein implicated in the regulation of many cellular processes, including cellular senescence, glucose homeostasis, aging, and longevity. 27 The activation of SirT1 mimics several metabolic aspects of calorie restriction that enhance selective nutrient utilization and mitochondrial oxidative function. 28 Furthermore, the AMPK/SirT1 signaling pathway is the mechanism by which hormones enhance mitochondrial metabolism, as in the case of adiponectin. 29 The regulation of AMPK and SirT1 are potential future therapeutic targets for the attenuation of complications of obesity.
SirT1 is a longevity-associated protein that regulates energy metabolism. The activation of SirT1 repressed PPARγ and, as a result, inhibited adipogenesis. 30 SirT1 decrease adipose tissue weight and stimulates brown remodeling of the white fat in white adipose tissue. 31 In this study, lower body fat accumulation was observed with M treatment, indicating probable involvement of this regulator. SirT1 and PPARγ might coordinate the regulation of adipogenesis and insulin sensitivity. 32,33 The impairment of CPT-1a mediated lipid oxidation is observed in obese patients, and the inhibition of fat oxidation by the CPT-1 inhibitor contributes to increased adiposity risk. 34 Although M has also been found to be effective in fatty liver by inhibiting PPARγ activation, it remains unclear as to the interplay between the AMPK signaling pathway and regulation of lipid disorders.
In conclusion, mangosteen exerts antiobesity effects in HF-induced obese mice by activating AMPK and SirT1 and by suppressing PPARγ expression in the liver. In addition, the administration of mangosteen to HF-induced obese mice reduced body weight gain, adipose mass, and serum TG, TC, and LDL levels. The antiobesity effect of mangosteen in HF-induced mice may support the use of mangosteen to reduce the risks of metabolic diseases.
Footnotes
Acknowledgments
This study was supported by a grant from the National Research Foundation of Korea (NRF) as funded by the government of South Korea (NRF-2012R1A1A2004424, Y.H. Choi) and the GRRC program of Gyeonggi Province, South Korea (GRRC DONGGK2015-B01, Development of new health supplements/therapeutics for neurodegenerative diseases and GRRC DONGGUK2015–B03, Development of functional food to alleviating metabolic syndromes and circulatory disorders).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.