
Abstract
One of the critical features of Alzheimer's disease is cognitive dysfunction, which is, in part, due to decreases in acetylcholine (ACh). The ethanol extract of Perilla frutescens was selected for isolating the acetylcholinesterase (AChE) inhibitor based on preliminary screening. In vivo behavioral tests were performed to examine the effects of the P. frutescens extract on trimethyltin chloride-induced impairment of learning and memory in mice. A diet containing P. frutescens extract effectively reversed learning and memory impairment on the Y-maze and passive avoidance tests. To isolate the active compound from the P. frutescens extract, solvent partitioning, silica gel open column chromatography, thin-layer chromatography, and high-performance liquid chromatography were used. The AChE inhibitor was identified as rosmarinic acid.
Introduction
A
AChE catalyzes the hydrolysis of the neurotransmitter, Ach, to choline and acetate in both the peripheral and central nervous systems (CNS). Clinical trials for AD patients have focused on drugs that increase ACh levels in the CNS. This approach to treatment has resulted in modest improvements in memory and cognition in some patients. Some trials that aim to delay the onset or to slow the progression of AD have focused on other aspects of the pathogenic cascade. Currently, AChE inhibitors that inhibit the hydrolysis of ACh to increase cholinergic neurotransmitters are widely used in patients. Synthetic AChE inhibitors, including tacrine, donepezil, and rivastigmine, have been tested in clinical settings. However, the side effects of these drugs, including hepatotoxicity and gastrointestinal disturbances, have led to the development of new AChE inhibitors from natural resources that are nontoxic and have outstanding activity. 2,4
Various phytochemicals have disease preventative effects due to their antioxidant, anti-inflammatory, antiviral, and anticarcinogenic properties. Some phytochemicals are potential preventive agents against cognitive disorders, including neurodegenerative disorders and AD. 6 In this study, Perilla frutescens extract was used to investigate the effect of AChE inhibition because it exhibited the highest inhibitory effect (34.8%) in preliminary screening.
P. frutescens (L.) Britton var. acuta Kudo is a member of the Lamiaceae family, primarily found in China, India, Thailand, Japan, and Korea. The beefsteak plant, also called perilla mint, is a traditional Asian crop used in cooking, as an herb, and as an ornamental plant. Perilla leaves are used as food and in traditional medicine. The leaves have antiallergic, anti-inflammatory, and antitumor-promoting effects. 7 The constituents of P. frutescens are perillaldehyde, limonene, pinene, shisonin, apigenin, luteolin, caffeic acid, and rosmarinic acid.
TMT decreases in ACh release in brain tissue and increases AChE activity in rat hippocampus. 8 We investigated the inhibitory effect of P. frutescens extract in vitro. To confirm the attenuating effect of the P. frutescens extract against trimethyltin (TMT)-induced learning and memory impairment, in vivo behavioral tests were performed. To separate and identify the active compound from the P. frutescens extract, solvent partition, open column chromatography, thin-layer chromatography (TLC), and high-performance liquid chromatography (HPLC) were used. After each separation step, all fractions were tested for an AChE inhibitory effect using the colorimetric method of Ellman. 9 The AChE inhibitor was identified as rosmarinic acid.
Materials and Methods
Chemicals
Acetylthiocholine iodide, 5,5′-dithiobis-(2-nitro) benzoic acid (DTNB), 9-amino-1,2,3,4-tetrahydroacridine (tacrine), dimethyl sulfoxide (DMSO), rosmarinic acid, trimethyltin chloride (TMT), 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS2−), 2,4-di-tert-butylphenol, ascorbic acid, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were purchased from Sigma (St. Louis, MO, USA). 2,2′-Azobis-(2-amidinopropane) dihydrochloride (AAPH) was purchased from Wako Chemicals USA, Inc., (Richmond, VA, USA).
Sample preparation
Dried P. frutescens was purchased from an herb shop in the Gyeong-dong medicinal herb market (Seoul, Korea). The dried P. frutescens (0.5 kg) was ground into a powder and dissolved in ethanol (2.5 L) by shaking for 24 h at 1.57 g. The ethanol extract was filtered through No. 42 filter paper (Whatman International Ltd., Middlesex, England). This extraction procedure was repeated five times. The ethanol extract was concentrated in a rotary evaporator (Eyela, Tokyo, Japan) under reduced pressure at 37°C.
Cell culture
The rat pheochromocytoma (PC12) cell line (CRL-1721) and human neuroblastoma (MC-IXC) cell line (CRL-2270) were obtained from the American Type Culture Collection (Manassas, VA, USA). RPMI 1640 medium, minimum essential medium (MEM), horse serum from a donor herd, fetal bovine serum (FBS), and antibiotic–antimycotic were purchased from Gibco-BRL (Grand Island, NY, USA). Sodium bicarbonate was purchased from Sigma. The cells were cultured on 100-mm diameter tissue culture dishes (Falcon, BD Biosciences, Franklin Lakes, NJ, USA) and maintained in a 37°C incubator in a humidified 5% CO2 atmosphere. The medium was refreshed approximately thrice per week. The PC12 cells were cultured in RPMI 1640 medium supplemented with 10% (v/v) horse serum from a donor herd, 5% (v/v) FBS, and 1% (v/v) antibiotic–antimycotic. When the cultured cells were 80–90% confluent (split ratio 1:4), they were subcultured. The MC-IXC cell line was cultured in MEM supplemented with 15% FBS (v/v) and 2% penicillin–streptomycin (v/v). Cells were subcultured (split ratio 1:4) using trypsin-EDTA once a week.
Measurement of AChE inhibition activity
AChE activity was determined using the modified spectrophotometric method of Ellman.
9
ACh iodide was used as the reaction substrate, and DTNB was used to measure AChE activity. For the enzyme preparation, PC12 cells were homogenized (Glass-Col homogenizer, Terre Haute, IN, USA) with Tris-HCl buffer [20 mM Tris-HCl (pH 7.5) containing 150 mM NaCl, 10 mM MgCl, and 0.5% Triton X-100]; this solution was then centrifuged at 10,000 g for 15 min. The supernatant was used as the enzyme source. The AChE protein concentration was determined by a protein assay kit using bovine serum albumin as the protein standard (Bio-Rad, Hercules, CA, USA). Briefly, 10 μL of each sample was mixed with 10 μL of enzyme solution, added to 70 μL of reaction mixture [50 mM sodium phosphate buffer (pH 8.0) containing 0.5 mM ACh iodide and 1 mM DTNB], and incubated at 37°C for 15 min. The ACh iodide and enzyme reaction was monitored at a wavelength of 405 nm using a 96-well microplate reader (GENios, Tecan Ltd., Männedorf, Switzerland).
Animals
Institute of Cancer Research (ICR) mice (5-week-old males) were purchased from Samtako Bio Korea (Gyeonggi, Korea) and were provided access to food and water ad libitum. The P. frutescens extract was mixed into the feed. Subsequently, TMT, which was dissolved in a sodium chloride solution, was administered through intraperitoneal (IP) injection. The control group was injected with the sodium chloride solution without TMT. Sample groups (Pf400, Pf800, Pf1200) were all injected with TMT (100 μL per mouse; 2.5 mg/kg body weight), but the animals of each group were maintained on diets differentially supplemented with P. frutescens extract (400, 800, and 1200 mg/kg per day, respectively) for 4 weeks. All experimental procedures abided by the guidelines established by the Animal Care and Use Committee of Korea University.
Y-maze test
The Y-maze test was performed 2 days after the TMT injection. The maze consisted of black plastic, with three maze arms that were each 33 cm long, 15 cm high, 10 cm wide, and positioned at an angle of 120° relative to the others. Each mouse was placed at the end of one arm and allowed to move freely through the maze during an 8-min period. The sequence of arm entries was recorded manually. Spontaneous alternation behavior was defined as entry into all three arms by consecutive choice in overlapping triplet sets. The percentage of spontaneous alternation behavior was calculated as the ratio of actual to possible alternations.
Passive avoidance test
The passive avoidance test was performed 2 days after the TMT injection. The apparatus consisted of an illuminated chamber connected to a dark chamber. An acquisition trial was performed on day 1. Each mouse was placed in the apparatus and left for 1 min with no light or shock, followed by a 2-min period with light and no shock, to habituate the mice to the apparatus. Subsequently, the mice were individually placed in the illuminated chamber. Immediately after entering the dark chamber, an inescapable scrambled electric shock (0.5 mA for 1 sec) was delivered through the floor grid (acquisition trial). The mice were then returned to their cages. Each mouse was again placed in the illuminated chamber 24 h later (retention trial). The interval between placement in the illuminated chamber and entry into the dark chamber was measured as the latency. The maximum testing limit for step-through latency was 300 sec.
Isolation of active compound
The active compound was purified sequentially by solvent partition, silica gel open column chromatography, TLC, and HPLC. The evaporated ethanol extract (145.6 g) was dissolved in distilled water (200 mL) and sequentially partitioned with hexane (600 mL ×3), chloroform (600 mL ×3), and ethyl acetate (600 mL ×3) for 24 h. Subsequently, each layer was separately concentrated in a rotary evaporator. The first ethyl acetate fraction (3.6 g) was isolated by silica gel column chromatography and added to a chromatographic open column packed with silica gel (90 g) suspended in chloroform. The column was washed with chloroform and eluted with a stepwise gradient consisting of chloroform and ethanol (100:0, 90:10, 80:20, 70:30, 60:40, 50:50, 40:60, 30:70, 20:80, 10:90, and 0:100, v/v, sequentially; repeated thrice). The solvent bed volume was 216 mL. The first fraction of the 90:10 gradient was isolated and concentrated in a rotary evaporator. The evaporated fraction (978 mg) was then dissolved in absolute ethanol (978 mg/mL), followed by separation on the silica gel TLC plate (30 × 30 cm; Merck, Darmstadt, Germany).
HPLC was used to identify the active compound from the band (Rf value = 0.15) of the TLC. The HPLC system (YL9100) was equipped with a PDA detector (YL9160), autosampler (YL9150), vacuum degasser (YL9101), binary pump (YL9111), and YL Clarity software version 3.0.4.444 (Young Lin Instruments Co., Inc., Anyang, Korea). Separation was achieved on a C18 column (4.6 × 250 mm, 5 μm Capcell Pak; Shiseido, Chuo-ku, Tokyo, Japan). The eluent flow rate was 1 mL/min. Gradient elution was carried out with 0.1% acetic acid in water (eluent A) and 0.1% acetic acid in acetonitrile (eluent B). The gradient elution program was as follows: 0–3 min, 5% B; 5 min, 10% B; 20 min, 30% B; 30 min, 50% B, and then held for 10 min before returning to the initial conditions. The total running time was 30 min. The column temperature was 25°C and the detection was performed at 330 nm.
VCEAC assay using ABTS radicals
ABTS radical anion was used as described previously. 10 Briefly, 1.0 mM AAPH was mixed with 2.5 mM ABTS2− in phosphate-buffered saline (pH 7.4). The mixed solution was heated in a water bath at 70°C for 30 min. The resulting ABTS solution was diluted to an absorbance of 0.650–0.020 at 734 nm. Twenty microliters of the sample was mixed with 980 μL of the ABTS radical solution. The mixture was incubated in a water bath at 37°C for 10 min. The decrease in absorbance at 734 nm was measured at the end of 10 min.
Measurement of cell viability
Cell viability was evaluated using MTT reduction. MC-IXC cells were preincubated with the sample for 24 h before TMT was added. They were then treated with TMT for 24 h. MTT reduction was initiated by adding 10 μL of MTT stock solution (2.5 mg/mL) per well, and the plates were incubated at 37°C. After a 3-h incubation, the reaction was stopped by adding 150 μL of DMSO. The absorbance was measured at 570 nm. The reference wavelength was then determined at 630 nm using a microplate reader (GENios, Tecan Ltd.). The results are given as a percentage relative to control cells, which were set to 100%.
Statistical analysis
Results are expressed as mean ± SD. Data were analyzed by one-way ANOVA, followed by Duncan's multiple range tests using SAS software (Cary, NC, USA).
Results
P. frutescens inhibition of AChE from PC12 cells
To search for an effective AChE inhibitor, various edible plants were screened using Ellman's method. In preliminary tests, the ethanol extract of P. frutescens exhibited one of the highest inhibitory activities among those tested (35%, data not shown). To confirm the AChE inhibition effect of P. frutescens, AChE inhibition was measured and compared with tacrine. The AChE inhibition standard curve was calculated by using the activity and concentration of tacrine. Tacrine inhibited AChE activity in a dose-dependent manner. As shown in Figure 1, the correlation between AChE inhibition and tacrine concentration was calculated (R 2 = 0.9986). The inhibition capacity of P. frutescens (1 mg/mL solution) was similar to that obtained using 336 nM of tacrine.
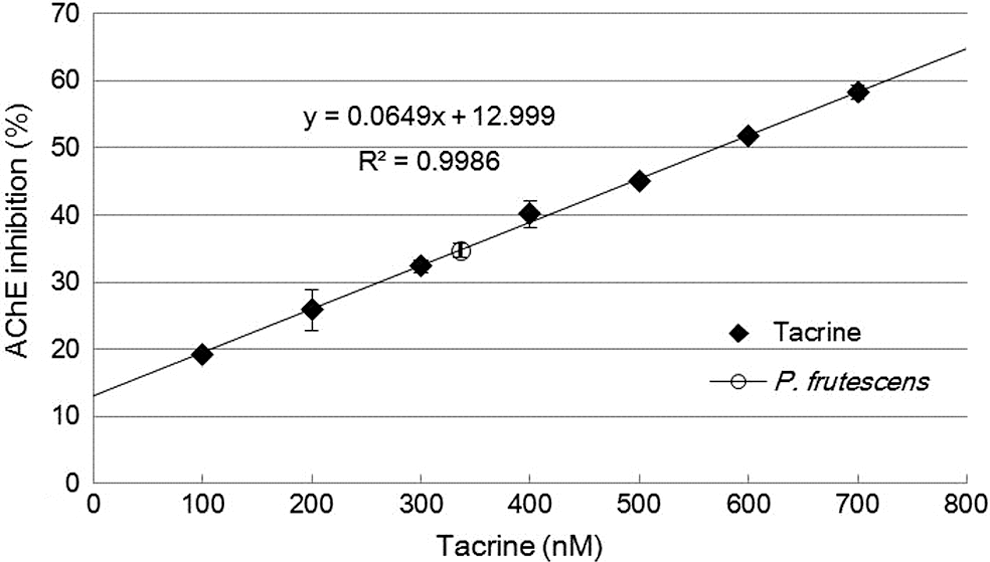
Acetylcholinesterase (AChE) inhibition by Perilla frutescens extract and tacrine.
Y-maze test
The Y-maze test was performed 2 days after the TMT injection. The TMT-injected mice exhibited significantly impaired spatial working memory (24% decrease in alternation behavior) compared with that of the control group (Fig. 2A). Mice fed with the P. frutescens extract diet displayed increased spontaneous alternation behavior after TMT injection. By contrast, the number of arm entries did not change among any of the experimental groups (Fig. 2B).
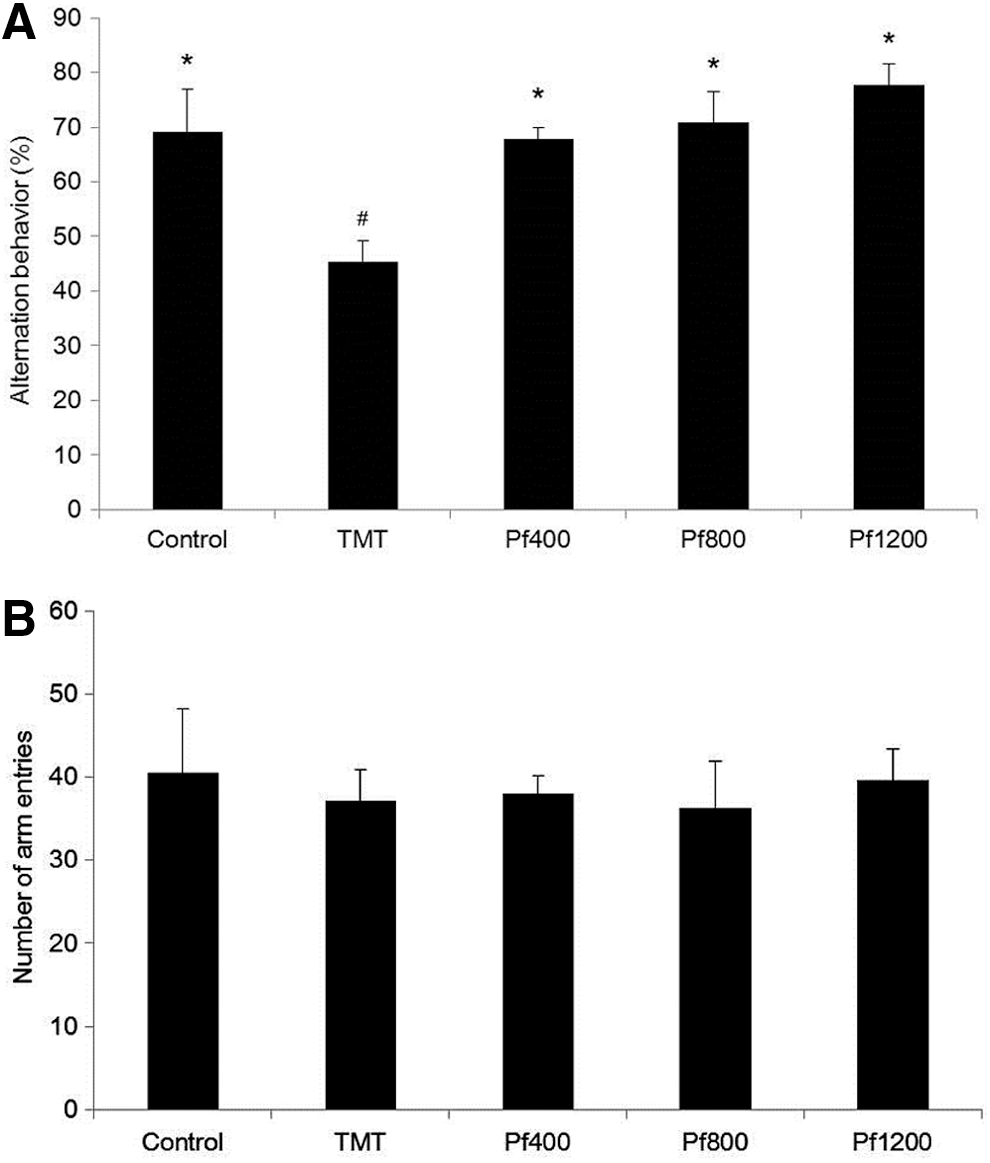
The effect of the P. frutescens extract on spontaneous alternation behavior in the Y-maze test.
Passive avoidance test
The passive avoidance test was performed 2 days after the TMT injection. The TMT-injected mice displayed a significant reduction (57-sec decrease) in step-through latency when compared with the control group. The P. frutescens extract diet attenuated TMT-induced impairment in mice during the passive avoidance test (Fig. 3).
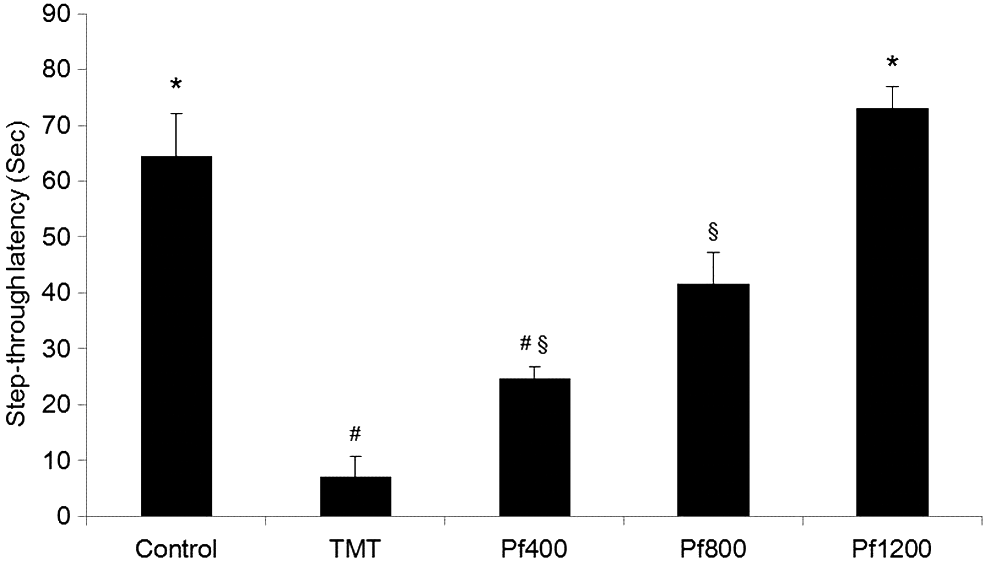
The effect of the P. frutescens extract on step-through latency in the passive avoidance test. Control, injected with sodium chloride solution; TMT, injected with TMT (2.5 mg/kg body weight); Pf400, injected with TMT (2.5 mg/kg body weight) and fed with P. frutescens extract diet (400 mg/kg of body weight per day); Pf800, injected with TMT (2.5 mg/kg body weight) and fed with P. frutescens extract diet (800 mg/kg of body weight per day); Pf1200, injected with TMT (2.5 mg/kg body weight) and fed with P. frutescens extract diet (1200 mg/kg of body weight per day). The step-through latency was determined during 300 sec. Values indicate the mean ± SD (n = 8), P < .01. Different superscript symbols (*, #, and §) represent statistical differences between groups.
Isolation of active compound
To separate the active compound from the P. frutescens extract, solvent partition, open column chromatography, TLC, and HPLC were used. The extract was sequentially partitioned thrice using n-hexane, chloroform, and ethyl acetate. The first fraction of ethyl acetate increased AChE inhibition effectively (32.49%; Fig. 4). On the basis of this result, the first fraction of ethyl acetate was divided into 33 subfractions by silica gel open column chromatography. Among these fractions, the fourth fraction (the first fraction with the chloroform-to-ethanol ratio of 90:10) exhibited the highest activity in the AChE inhibition assay (39.36%; Table 1). The fourth fraction was then subjected to TLC to separate out the active compound. Among 16 fractions, the third band (Rf value of 0.15) showed the highest inhibition of AChE (35.62%; Fig. 5). To identify the active compound in this fraction, the isolated fraction and the ethanol extract were further analyzed. The composition of the P. frutescens ethanol extract was analyzed by HPLC. Major compounds were separated on the column. The significant peak appeared at 23 min in the isolated fraction (Fig. 6A). The purified active compound was identified as rosmarinic acid.
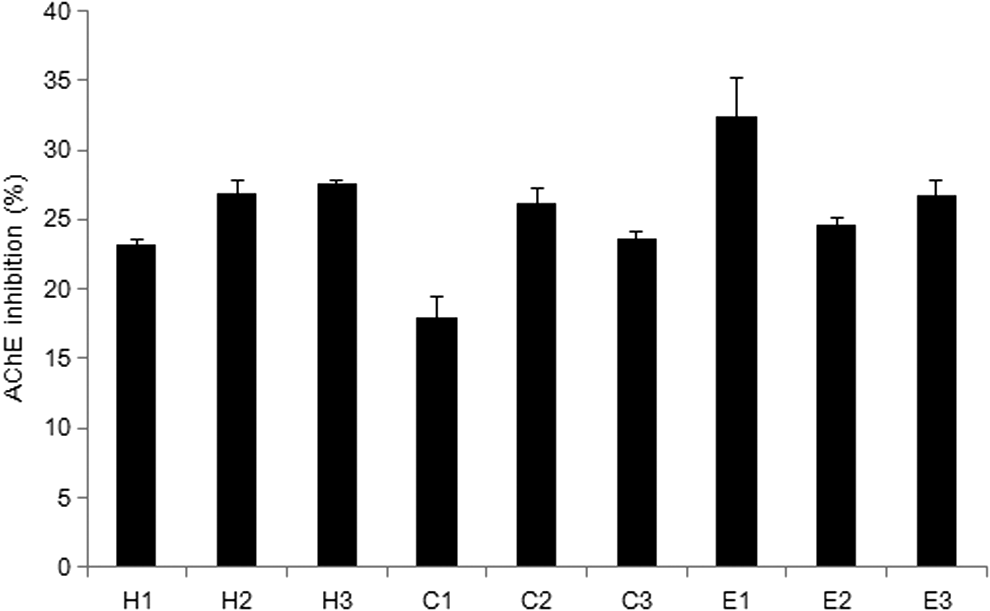
The AChE inhibitory effect of the solvent-partitioned P. frutescens extract. The sample groups were treated with samples that were fractionated with hexane (H1–H3), chloroform (C1–C3), and ethyl acetate (E1–E3). Values indicate the mean ± SD (n = 6).
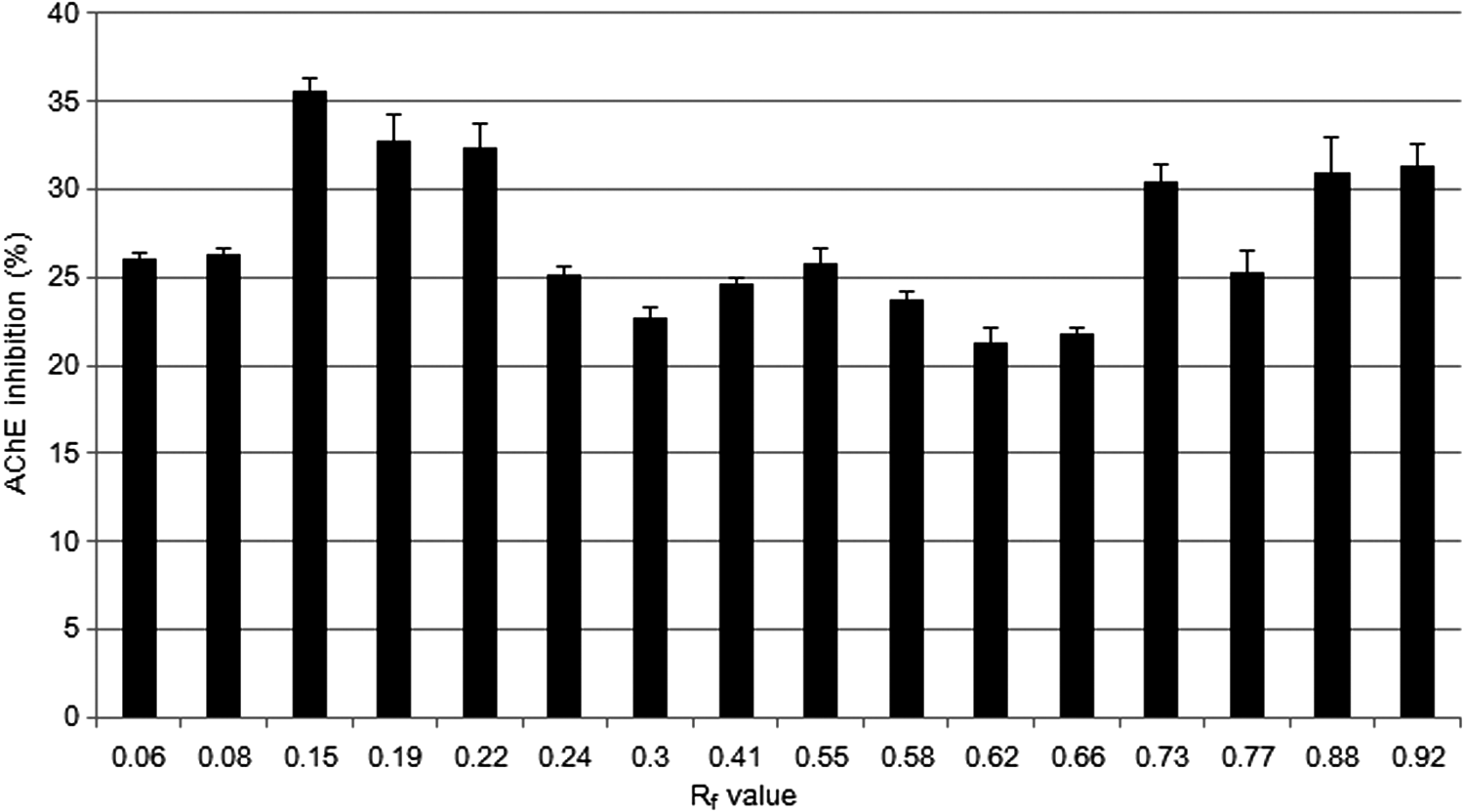
The AChE inhibitory effects of P. frutescens extract separated by thin-layer chromatography (TLC). The sample groups were treated with samples that were separated by TLC. Values indicate the mean ± SD (n = 6).
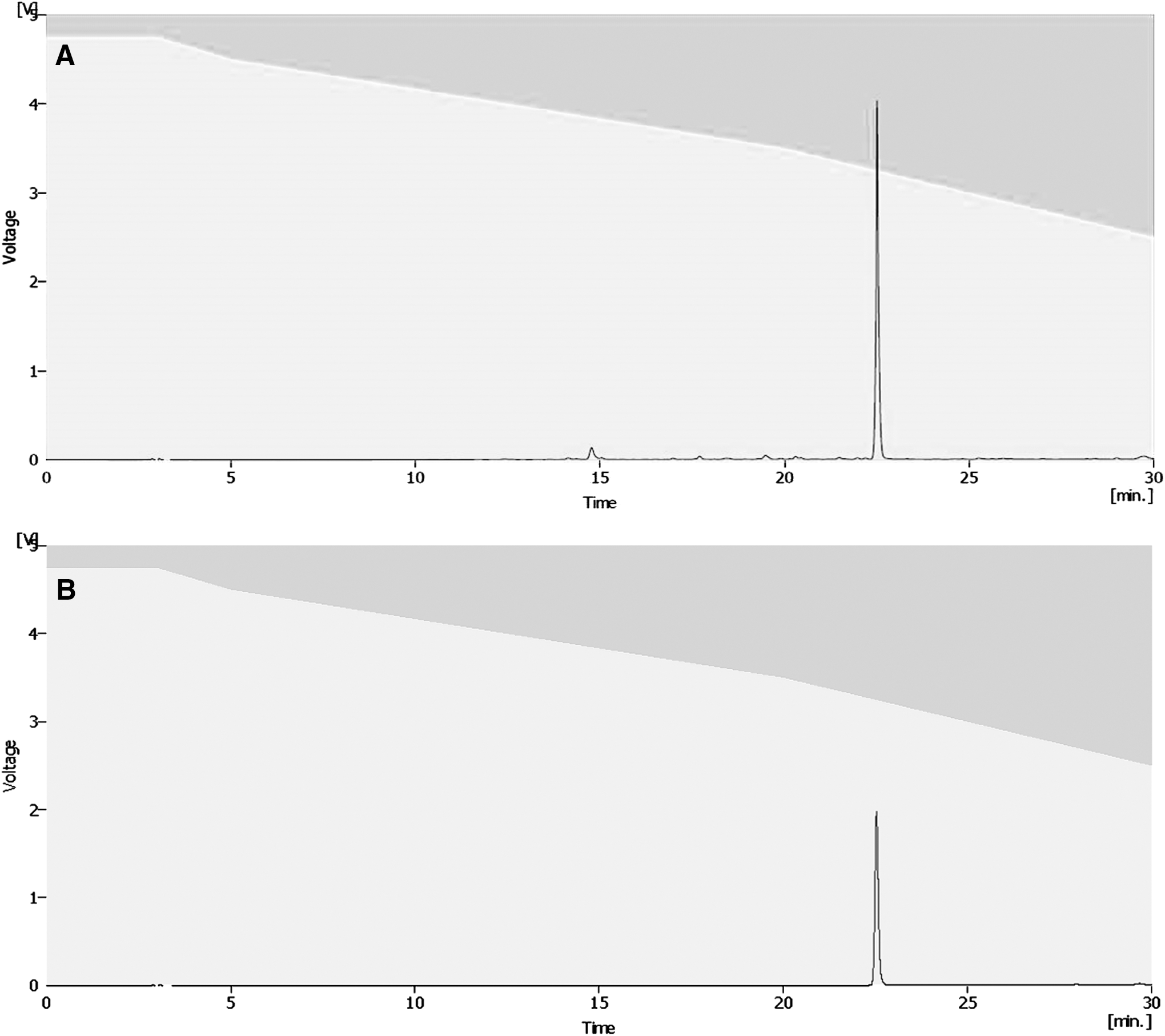
Isolation of rosmarinic acid from P. frutescens extract by using high-performance liquid chromatography (HPLC).
Bold values indicate the mean ± SD (n = 8), p < .01.
AChE, acetylcholinesterase.
Vitamin C equivalent antioxidant capacity
The antioxidant potential of the rosmarinic acid was evaluated by using ABTS radical anions. The vitamin C equivalent antioxidant capacity (VCEAC) standard curve was calculated by plotting the decrease in absorbance against concentration of vitamin C. Antioxidant capacity of rosmarinic acid was converted to VCEAC. As shown in Figure 7A, rosmarinic acid (R 2 = 0.9975) and vitamin C (R 2 = 0.9993) both scavenged ABTS radical anions in a dose-dependent manner. The correlation between rosmarinic acid and VCEAC was calculated (R 2 = 0.9975). The antioxidant capacity of rosmarinic acid was double that of vitamin C (Fig 7B).

Antioxidant capacity of rosmarinic acid.
Effect of rosmarinic acid on MC-IXC cell viability
To evaluate the protective effect of the extract against TMT-induced neurotoxicity, an MTT reduction assay was used to measure the cell viability. As shown in Figure 8, cell viability increased due to rosmarinic acid exposure in a dose-dependent manner (1 mM, 287.46%; 2 mM, 342.45%; 4 mM, 387.23%; 5 mM, 418.65%). These data demonstrate the potent protective effect of rosmarinic acid against TMT.
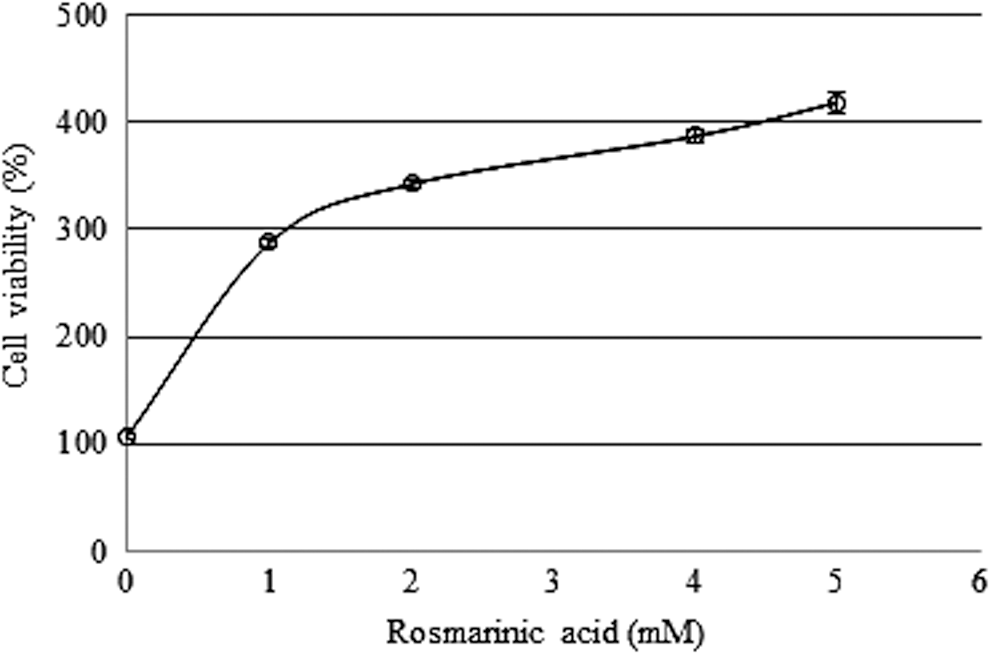
Protective effect of rosmarinic acid against TMT-induced cell death. Sample groups were preincubated with rosmarinic acid for 24 h before 200 μM TMT treatment. Values indicate the mean ± SD (n = 6).
Effect of rosmarinic acid on AChE inhibition
The inhibitory effect of rosmarinic acid was confirmed. As shown in Figure 9, the AChE inhibition was increased by rosmarinic acid exposure (1 mM, 23.84%; 5 mM, 33.98%; 10 mM, 40.33%). Treatment with rosmarinic acid increased AChE inhibition in a dose-dependent manner.
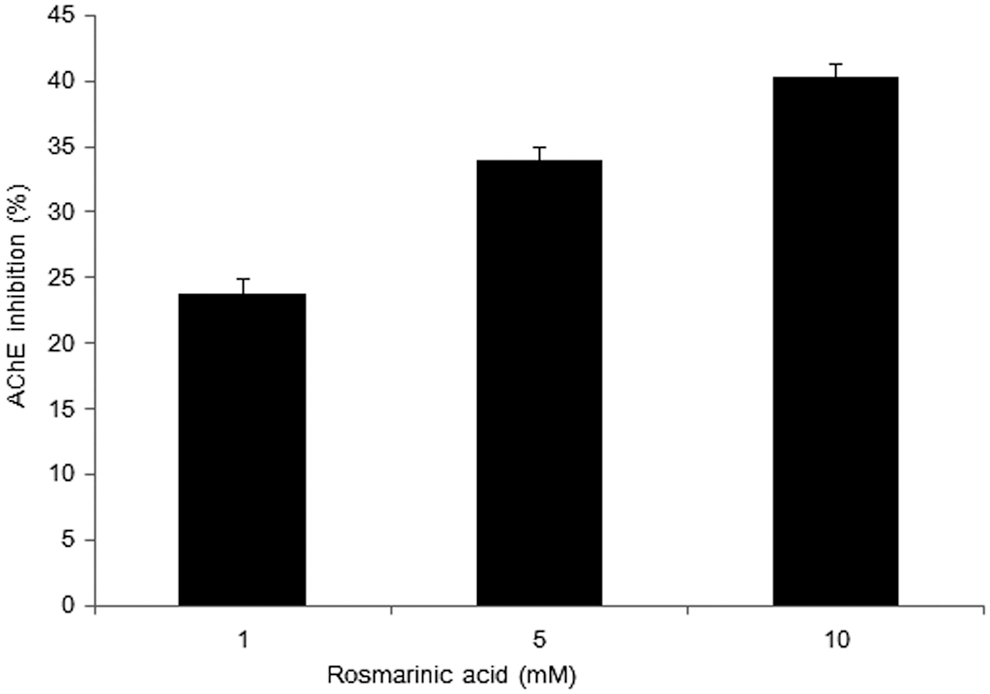
The AChE inhibitory effects of rosmarinic acid. The sample groups were treated with rosmarinic acid. Values indicate the mean ± SD (n = 6).
Discussion
One of the significant factors in AD is the loss of forebrain cholinergic neurons and the consequent loss of ACh. The accompanying decrease in cholinergic activity in turn affects the activity of the Ach-synthesizing enzyme. The cholinergic deficit in AD patients is demonstrated by reduced choline acetyltransferase (ChAT) activity and increased AChE activity. The resulting cholinergic deficit contributes to the cognitive impairment seen in AD patients and has led to the development of AChE inhibitors for use as therapeutic agents. 11 AChE inhibitors decrease the hydrolysis of ACh in the brain, thereby boosting cholinergic neurotransmission. 4 This effect was based on the premise that boosting ACh levels would attenuate the neuronal deficits and cognitive impairments present in AD patients.
In our preliminary test, the ethanol extract of P. frutescens exhibited one of the highest AChE inhibitory activities. Based on this screening, the inhibitory activity of P. frutescens was investigated further. The inhibitory effect of P. frutescens extract on AChE was compared with the synthetic AChE inhibitor, tacrine. Despite the extract being crude, its inhibitory capacity was approximately 35% of that for tacrine. This result demonstrated that P. frutescens extract attenuated hydrolysis of ACh in vitro. It is hypothesized that the extract may boost cholinergic neurotransmission.
In addition, the effect of dietary administration of the extract on behavioral ability was tested using an AD animal model following IP injection of TMT. One of the mechanisms of TMT-induced neurodegeneration is that TMT causes a decrease in ACh release in brain tissue and increases AChE activity ex vivo. 12,13 Memory and learning abilities were evaluated in a Y-maze test and a passive avoidance test. The Y-Maze test is a behavioral test for measuring the willingness of mice to explore new environments. Mice prefer to investigate a new arm of the maze rather than returning to one that was previously visited. Many parts of the brain are involved in this task. This test is used to quantify cognitive deficits in mouse models and to evaluate the effects of active compounds on cognition. The TMT injection induced significant memory and learning impairments evident in both the Y-maze and passive avoidance tests. Spontaneous alternation behavior, a recognized measure of spatial memory, was examined using the Y-maze test. Our studies suggest that a diet containing the extract of P. frutescens had a protective effect against learning and memory deficits. The number of arm entries was not significantly different among the groups. These results indicate that the capacity for general locomotion was not affected by TMT injection or the sample diet, but that the decrease in spatial memory was attenuated by the P. frutescens extract. The passive avoidance test is used to evaluate learning and memory in mouse models of neurodegenerative disorders. In this test, the mice learn to avoid an electric shock in acquisition trials, which occur before testing. The step-through latency for exit from the chamber in which the shock occurs, to avoid the stimulus, is used as an indicator of learning and memory. The passive avoidance test is useful for evaluating the effects of the compound on learning and memory as well as for studying the mechanisms involved in cognition. Mice treated with the P. frutescens extract exhibited a dose-dependent attenuation of TMT-induced impairment of passive avoidance performance. Thus, P. frutescens extract displayed a significant antiamnestic effect in our mouse model. Based on these results, it is possible that the AChE inhibition effects of the P. frutescens extract can explain the antiamnestic effects observed in this study.
Phenolic acids are secondary plant metabolites that are important determinants of the sensory and nutritional quality of fruits, vegetables, and other plants. Many phenolic acids may function as chemopreventive agents. 14,15 In addition, several compounds inhibit neurodegeneration due to their bioactive functions. 15 –18 In this study, P. frutescens extract was used to investigate the effect of AChE inhibition. There are various compounds in P. frutescens extract. To separate the best active compound from the extract, solvent partition, open column chromatography, TLC, and HPLC were used. By using a sequential purification process, the active compound in the extract was identified as rosmarinic acid. Rosmarinic acid is a caffeic acid ester found in a variety of plants. Rosmarinic acid and its derivatives have antioxidant, anticancer, antiviral, antibacterial, and anti-inflammatory activity. 19,20 The antioxidant capacity of rosmarinic acid was confirmed using the ABTS radical assay. Rosmarinic acid exhibits significant antioxidant activity in a dose-dependent manner. TMT-induced neuronal damages generate reactive oxidative stress (ROS). 21 Previous research showed that TMT elevates formation of a major product of lipid peroxidation, 4-hydroxynonenal (4-HNE). 22 In our study, to evaluate protective activity against TMT-induced neurotoxicity, cell viability was measured. The rosmarinic acid dose-dependently protected MC-IXC cells from TMT-induced cell death. This result demonstrates that rosmarinic acid has the ability to ameliorate TMT-induced neuronal damage such as oxidative stress. In addition, the AChE inhibitory effects of rosmarinic acid were confirmed. The AChE inhibition was increased by rosmarinic acid exposure in a dose-dependent manner. By decreasing AChE activity, P. frutescens extract was shown to possess strong antiamnestic properties. There is no direct relationship between antioxidative activity and AChE inhibition. However, one of the mechanisms of TMT-induced neurodegeneration is that TMT causes an increase in AChE activity. Another mechanism is that TMT-induced neuronal damage generates ROS. Based on these mechanisms, antiamnestic properties of rosmarinic acid were due to antioxidative activity and AChE inhibition capacity of rosmarinic acid.
In conclusion, AChE inhibitory efficacy was investigated in P. frutescens extract. By using a sequential purification process, the active compound in the extract was identified. P. frutescens ethanol extract, containing the active compound rosmarinic acid, attenuates memory and learning impairment by inhibiting TMT-induced neuronal death and AChE in a dose-dependent manner. These results suggest that P. frutescens and rosmarinic acid are potentially applicable as protective agents against the development of AD.
Footnotes
Acknowledgment
This research was supported by the High Value-added Food Technology Development Program, Ministry of Agriculture, Food and Rural Affairs.
Author Disclosure Statement
No competing financial interests exist.