
Abstract
This study provides the scientific basis for the inhibitory effect of the aerial parts of Cichorium intybus Linn. (C. intybus) on the activation of the NLRP3 inflammasome in vitro and on high-fat diet (HFD)-induced type-2 diabetes (T2D). Lipopolysaccharide (LPS)-primed bone marrow-derived macrophages were used to study the effects methanolic extract of C. intybus leaf (CI) on inflammasome activation. An insulin resistance model (mice fed a HFD) was used to study the in vivo effect of CI on T2D. CI attenuated interleukin-1β (IL-1β) secretion by inhibiting the activation of the NLRP3 inflammasome in mouse bone marrow macrophages. The CI treatment attenuated the intracellular movement of NLRP3 in Triton X-100 insoluble fraction, without affecting the expression of other NLRP3 inflammasome-related proteins. Attenuated IL-1β secretion may improve glucose metabolism in the HFD-fed insulin resistance mouse model. CI also attenuated the infiltration of M1 macrophages and increased the M2 macrophage population in white adipose tissue. Collectively, our data showed that CI inhibits IL-1β secretion through attenuation of NLRP3 inflammasome activation, leading to an antidiabetic effect by improving glucose metabolism and inhibiting metainflammation.
Introduction
T
Obesity is frequently the basis of insulin resistance in T2D, which is characterized by insulin resistance and islet beta-cell dysfunction. 4 Adipose tissue macrophages consist of at least two different phenotypes: classically activated M1 macrophages and alternatively activated M2 macrophages. 5 The general view of obesity is that an imbalance exists in the ratio of M1/M2 macrophages, with stimulation of M1 macrophages and downregulation of M2 macrophages leading to chronic inflammation and the propagation of metabolic dysfunction. 6
Cichorium intybus Linn. (C. intybus, Chicory) from the family Asteraceae provides a variety of edible products and widely used as an important medicinal herb to treat various ailments, including diabetes. 7 C. intybus also has immune system-promoting properties and is considered one of the best coffee substitutes. 8 Pharmacologically, chicory roots were reported to possess antidiabetic 9 and antihypertriglyceridemia activities. 10 However anti-inflammasome and inflammasome-attenuated antidiabetic activity of C. intybus has not been reported yet. In the present study, we investigated the protective effects of the aerial parts of C. intybus extract against HFD-induced T2D. Furthermore, we performed in vitro experimental studies to elucidate the underlying mechanisms involved in the in vivo efficacy of C. intybus extract in T2D.
Materials and Methods
Reagents and antibodies
IL-1β antibody and IL-1β enzyme-linked immunosorbent assay (ELISA) kit were purchased from R&D Systems (Minneapolis, MN, USA). ASC and NLRP3 antibodies were purchased Enzo Life Science (Farmingdale, NY, USA). Caspase-1 and β-actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Silica crystal, nigericin, poly(dA:dT), flagellin, and zVAD-FMK were purchased from InvivoGen (San Diego, CA, USA). Lipopolysaccharide (LPS; Escherichia coli 026:B6), ATP, and omega-3 fatty acid were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Plant material
Methanolic extract of C. intybus leaf (CI) was purchased from the Korea Research Institute of Bioscience and Biotechnology (KRIBB; Korea). It was authenticated by Dr. Shin-Ho Kang, a taxonomist at KRIBB, based on its microscopic and macroscopic characters. A voucher specimen (KRIB 028–086) was deposited at the herbarium of Plant Extract Bank at KRIBB for future reference. Dried aerial parts of C. intybus (30 g) were powdered and extracted with methanol using Soxhlet's extraction technique for 3 days. The extract was then concentrated under reduced pressure and lyophilized. The final yield of the extract was 5% (w/w), and it was stored at 4°C. The lyophilized powder of CI was dissolved in 10% dimethyl sulfoxide (DMSO) and then filtered through a 0.22-μm syringe filter and stored as stock until used in each experiment. The final concentration of DMSO was 0.1%.
Animals
Male C57BL/6 mice (20–22 g, 6 weeks old) were purchased from Orient Bio Co. (Seongnam, Korea). They were housed in groups of five under standard conditions (temperature 22 ± 2°C, humidity 55 ± 5%, 12-h light/12-h dark cycle), with food and water ad libitum. The animals were allowed to adapt to the laboratory environment for 5–7 days before experimentation. All experiments were performed in accordance with the guidelines of the Konkuk University Animal Care Committee (Korea) and the “Guide for the Care and Use of Laboratory Animals” adopted by the United States National Institutes of Health. The study protocol was reviewed and approved by the Ethics of Animal Experiments Committee of Konkuk University (Republic of Korea) (Permit No. KU14059).
Metabolic studies
The total experimental period was 10 weeks. Mice were randomly divided into three groups as follows: group fed a ND, group fed a HFD, and group treated with 50 mg/kg of CI. In the treatment group, CI (50 mg/kg) was administrated twice a week by oral gavage for 6 weeks after free access to HFD for 4 weeks. Samples for testing fasted blood glucose levels were collected with a glucometer (Roche, Mannheim, Germany). For glucose tolerance testing, mice fasted for 8 h and were injected intraperitoneally with glucose at a dose of 1.5 mg/g body weight. Blood samples were obtained at different time points for glucose measurements using the glucometer.
Real-time polymerase chain reaction
Total RNA was isolated from cells or tissue using QIAzol Lysis Reagent (QIAGEN, Hilden, Germany) according to the manufacturer's instructions. One microgram of RNA was reversely transcribed to make complementary DNA (cDNA) using a Maxima H Minus First Strand cDNA Synthesis Kit (Thermo Scientific, Hudson, NH, USA). Real-time polymerase chain reaction (PCR) was performed using 2X SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) with an iCycler (Bio-Rad), using each of the following primers: iNOS (5′-TCACCTTCGAGGGCAGCCGA-3′ and 5′-TCACCTTCGAGGGCAGCCGA-3′), TNF-α (5′-TCACCTTCGAGGGCAGCCGA-3′ and 5′-AGCAATGACTCCAAAGTAGACC-3′), IL-1β (5′-AGCAATGACTCCAAAGTAGACC-3′ and 5′-ATCTTTTGGGGTCCGTCAACT-3′), NLRP3 (5′-AGAGCCTACAGTTGGGTGAAATG-3′ and 5′-CCACGCCTACCAGGAAATCTC-3′), Arg1 (5′-ATCTTTTGGGGTCCGTCAACT-3′ and 5′-CCACACTGACTCTTCCAT TCTT-3′), IL-10 (5′-CCACACTGACTCTTCCATTCTT-3′ and 5′-CCACACTGACTCTTCCATTCTT), and HPRT (5′-TCAGTCAACGGGGGACATAAA-3′ and 5′-GGGGCTGTACTGCTT AACCAG-3′). The primers were purchased from Bioneer (Seoul, Korea). The PCR condition was 7 min at 95°C, followed by 40 cycles of 95°C, 55°C, and 72°C for 30 s. The resulting mean −ΔCt values from each group were then used to calculate the relative changes in messenger RNA (mRNA) expression as the ratio (R) of mRNA expression.
High-performance liquid chromatography and fingerprint analysis of CI
CI was dissolved in 100% methanol and filtrated through a 0.45-μm PTFE disposable syringe filter (Advantec; Toyo Roshi Kaisha Ltd., Tokyo, Japan) for fingerprint analysis. The components of CI were determined with a Prominance LC-20A (Shimadzu, Kyoto, Japan) high-performance liquid chromatography (HPLC) system, equipped with a CMA-20A communications bus module, LC-20AD liquid chromatograph, SPD-M20A diode array detector, SIL-20AC auto sampler, DGU-20A3 degasser, and CTO-20AC column oven. The constituents of the extract were separated using a Shim-pack VP-ODS column (4.6 × 250 mm, 5 μm) at a flow rate of 0.15 mL/min and an injection volume of 10 μL. The mobile phase was acetonitrile, with the following gradient programs shown in Table. 1. The mass range (m/z) was 100–1500 amu.
HPLC, high-performance liquid chromatography.
To analyze the concentration of known components in peaks, 5 mg of lactucin and lactucopicrin was dissolved in 100 mL methanol. Eleven milligrams of CI was extracted with 500 μL methanol (80%, v/v) and sonicated for 5 min. The extract was centrifuged for 5 min at 900 g, and the supernatant was used for the determination of analytical sample. Chromatographic separation was carried out using an Agilent 1200 series. Separations were performed using Eclipse C18 (250 × 4.6 mm, 5 μm; Agilent, Santa Clara, CA, USA). The eluent was monitored at 280 nm of diode array detector. The mobile phase was composed of 0.5% of acetic acid in water (A) and methanol (B): 0–5 min, 95% A; 5–30 min, 95–72% A; 30–80 min, 72–40% A; 80–90 min, 40–0% A; 90–100 min, 0% A; 100–110 min, 0–95% A; 110–120 min, 95% A. The flow rate used was 1.0 mL/min. Column temperature was maintained at 30°C.
Other experimental methods
Experimental conditions for bone marrow cell culture, cell viability test (MTT), immunoblot analysis, and NLRP3 inflammasome activation were followed, as described in our previous report. 11 The concentration of CI used was 25, 50, and 100 μg/mL.
Statistical analysis
The results are expressed as the mean ± standard error of the mean (SEM) of at least three independent experiments (n = 3). Statistical analysis was performed using the Student's t-test by GraphPad (San Diego, CA, USA) Prism software, and P-values of <.05 were considered statistically significant.
Results
CI inhibited the activation of the NLRP3 inflammasome
Inflammasomes are multiprotein complexes that operate as platforms for the activation of caspase-1 and are composed of one of several NLRPs proteins, including NLRP1, NLRP3, NLRC4, and AIM2. 12,13 We investigated the possible capacity of CI on attenuation of IL-1β secretion through modulation of inflammasome activation. To compare inhibitory function of various extracts from C. intybus on NLRP3 inflammasome activation, LPS-primed bone marrow-derived macrophages (BMDMs) were treated by extracts from water, ethanol, or methanol at a nontoxic condition, followed by stimulation with NLRP3 inflammasome activators, such as ATP, nigericin, or silica crystals (Fig. 1A). As shown in Figure 1B–D, methanolic extract of C. intybus showed the strongest inhibitory effect on NLRP3 inflammasome activation among three kinds of extracts. Therefore, we examined the effects of CI on the activation of various inflammasomes, including NLRP3, AIM2, and NLRC4. CI treatment alone (up to 200 μg/mL) and the medium (0.1% DMSO) did not show any signs of cytotoxicity in the BMDMs, as estimated with an MTT assay (Fig. 1E). To elucidate the effect of CI on inflammasome activation, the LPS-primed BMDMs were treated with various concentrations of CI with or without NLRP3 inflammasome activators, such as ATP, nigericin, and silica crystals. As shown in Figure 1F, G, CI significantly inhibited ATP and nigericin-induced processing of both IL-1β and caspase-1. It also inhibited the cytosolic protein level of NLRP3, but not other NLRP3 inflammasome components. CI also inhibited silica crystal-induced processing of IL-1β (Fig. 1H). To investigate the inhibitory effect of CI on the other type of inflammasomes (i.e., AIM2 and NLRC4), the LPS-primed BMDMs, with or without double stranded DNA (dsDNA) or flagellin of Salmonella typhimurium, were treated with CI. As shown in Figure 1I, J, CI did not inhibit the processing of IL-1β from either the dsDNA-induced AIM2 inflammasome or the flagellin-induced NLRC4 inflammasome. Collectively, these data indicated that CI specifically inhibited the activation of the NLRP3 inflammasome, but not that of the AIM2 and NLRC4 inflammasomes.

Effect of CI on inflammasome activation.
CI attenuated HFD-induced T2D
IL-1β, released from NLRP3 inflammasome activation, is known to be implicated in the pathogenesis of T2D as well as in the development of T1D. 1 Furthermore, some diseases, such as gout, T2D, and atherosclerosis, are considered “particle diseases” because insoluble particles play a role in their pathogenesis. 14 These particles have been shown to be phagocytosed by macrophages followed by NLRP3 inflammasome activation. 1 As CI was shown to a strong inhibitory effect on silica-induced-IL-1β secretion (Fig. 1H), we examined whether CI might improve T2D.
To evaluate the possible effect of CI on T2D, we treated HFD-fed mice with CI by oral administration for 6 weeks (Fig. 2A). As shown in Figure 2B, the body weight gradually increased in the mice fed the HFD compared with those fed a normal diet. The CI and omega-3 treatment did not have any effect on the body weight of the mice fed the HFD. To explore the role of CI in HFD-induced insulin resistance, we conducted a glucose tolerance test. As shown in Figure 2C, the blood glucose content was highest in the mice fed the HFD, and glucose metabolism and insulin sensitivity were significantly improved in the CI-treated mice. The omega-3, used as positive control in our study, also showed a similar effect. 15 To confirm that CI prevents HFD-induced T2D through inhibition of NLRP3 inflammasome activation, we tested whether CI treatment suppresses the IL-1β level in the serum of HFD-fed mice. As shown in Figure 2D, the IL-1β production was significantly increased in HFD-fed mice serum, but CI treatment significantly inhibited HFD-induced IL-1β production in serum.

Effect of CI on the HFD-fed insulin resistance model mice.
During the progression to obesity in HFD-fed mice, a phenotypic switch occurs in adipose tissue macrophages from a M2 to an M1 phenotype, and these M1-type macrophages contribute to the development of insulin resistance. 16 To evaluate the inhibitory effect of CI on M1 macrophage infiltration in the HFD-induced T2D model, we measured the marker gene expression of M1 and M2 in epididymal white adipose tissue. The gene expression level of M1 macrophage markers, such as iNOS, TNF-α, and IL-1β (Fig. 2E–G), was high in the HFD-fed mice. However, the CI treatment attenuated the expression of these M1 macrophage marker genes. It also downregulated elevated NLRP3 gene expression in the HFD-fed mice (Fig. 2H). Furthermore, the CI treatment resulted in elevated expression of M2 macrophage marker genes, such as Arg-1 and IL-10 (Fig. 2I, J). Collectively, CI had a protective effect against HFD-induced T2D through inflammasome inactivation, followed by an improvement in glucose metabolism, attenuation of metainflammation, and M1 macrophage infiltration.
CI attenuated NLRP3 inflammasome activation through regulation of NLRP3
To understand the mechanisms underlying the inhibition of NLRP3 inflammasome activation, we paid attention to the reduced expression level of NLRP3 in the CI-treated cell lysates in Figure 1F, G. Proteasomal degradation of NLRP3 by ubiquitination could be one of the plausible explanations for the reduction of NLRP3 expression in cell lysates. To determine whether CI reduced NLRP3 protein expression through proteasomal degradation, we treated the BMDMs with MG132, a proteasome inhibitor, before the CI treatment of the LPS-primed BMDMs. As shown in Figure 3A, the reduction of NLRP3 expression by CI was not restored by the treatment of MG132, which indicated that CI did not influence the proteasomal degradation of NLRP3. Recent studies suggested that NLRP3 could be degraded by autophagy. 17,18 We, therefore, determined whether CI reduced NLRP3 protein expression through the activation of autophagy. We treated the BMDM with 3-Methyladenine (3-MA), an autophagy inhibitor, before the CI treatment. As shown in Figure 3B, CI inhibited NLRP3 protein expression, but 3-MA did not restore the NLRP3 protein expression level. Shimada et al. suggested that treatment of BMDMs with NLRP3 inflammasome inducers, such as ATP, nigericin, and silica crystals, caused loss of mitochondrial membrane potential and mitochondrial dysfunction. 19 To determine whether CI improves the loss of mitochondrial membrane potential, we treated the LPS-primed BMDMs with nigericin or silica crystals, with or without CI, followed by MitoTracker Deep Red staining. As shown in Figure 3C, the CI treatment alone did not affect the mitochondrial membrane potential, but nigericin or silica treatment reduced the mitochondrial membrane potentials. However, the CI treatment did not restore the nigericin- or silica-induced loss of mitochondrial membrane potential. To further study the direct effect of CI on the expression in NLRP3 inflammasome components, we examined the expression level of NLRP3, Caspase-1, ASC, and IL-1β induced by LPS treatment in both protein and mRNA. Post-treatment with CI of the LPS-primed BMDMs inhibited NLRP3 protein expression in a dose-dependent manner, but it did not influence the expression of other NLRP3 inflammasome-related proteins (Fig. 3D). Furthermore, CI did not affect mRNA transcription of NLRP3 and IL-1β (Fig. 3E, F). Collectively, these data indicated that inhibition of both IL-1β production and activation of the NLRP3 inflammasome by CI treatment was not due to proteasomal degradation, autophagy, loss of mitochondrial potential, or transcriptional regulation.
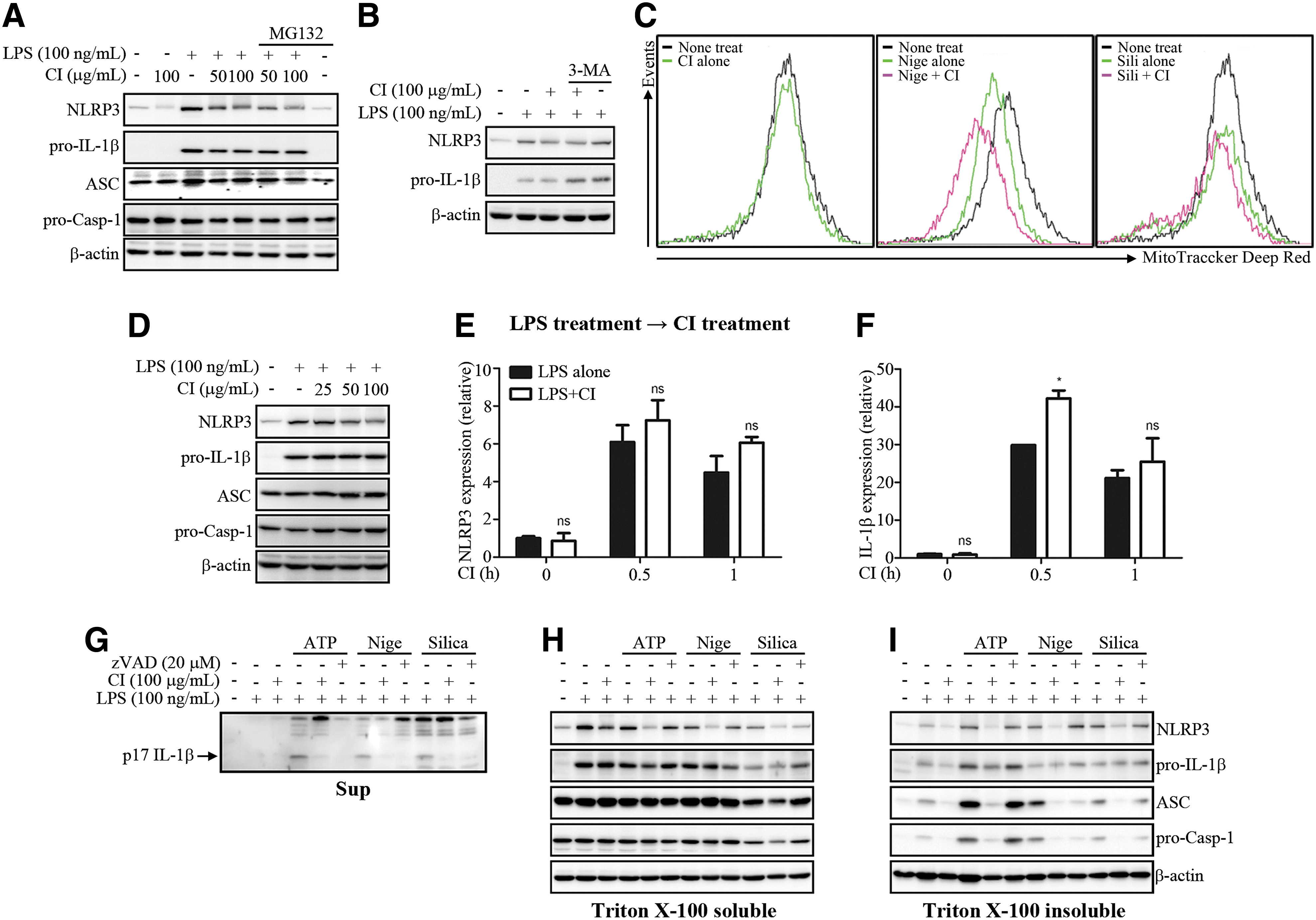
Action mechanism of CI on NLRP3 activation. The LPS-primed BMDMs were pretreated with MG132 (20 μM)
A recent study suggested that NLRP3 inflammasome components, such as NLRP3 and ASC, were detected from mitochondria-associated ER membranes (MAMs) fraction during NLRP3 inflammasome activation process. 20 Hara et al. reported that ASC and its oligomerization were detected in Triton X-100 insoluble fraction in NLRP3 inflammasome-activated BMDMs. 21 Accordingly, we examined NLRP3 inflammasome components in Triton X-100 soluble and insoluble fraction. CI inhibited IL-1β secretion (Fig. 3G) and significantly inhibited NLRP3 protein expression in Triton X-100 soluble fraction (Fig. 3H). Interestingly, CI strongly reduced the translocation of NLRP3 components, such as NLRP3, ASC, and pro-caspase-1, to the Triton X-100 insoluble fraction (Fig. 3I). In NLRP3 inflammasome activation process, ASC translocation from Triton X-100 soluble fraction to Triton X-100 insoluble fraction is essential for NLRP3 inflammasome activation. 21 Both NLRP3 and ASC were detected from MAMs fraction in NLRP3 inflammasome-activated macrophages. 20 Collectively, these data indicated that CI inhibited NLRP3 inflammasome activation through the obstruction of translocation of NLRP3 and ASC to Triton X-100 insoluble fraction. However, the action mechanism of CI for the inhibition of translocation and the downregulation of NLRP3 in the protein level remain to be elucidated.
HPLC fingerprinting analysis of CI and inhibitory effect of lactucin on NLRP3 inflammasome activation
Extensive phytochemical analyses of C. intybus have revealed the presence of vitamins, carotenoids, inulin, and sesquiterpenes, including lactucin and lactucopicrin. 22,23 In the present work, the HPLC fingerprint analysis of CI extract showed peaks of major components of lactucopicrin (71.9 mg/kg) and lactucin (42.2 mg/kg), with different UV absorption conditions (Fig. 4A). The results are in agreement with previously reported works, which found that the aerial parts of C. intybus contain sesquiterpene lactones, such as lactucopicrin and lactucin. 24 Lactucopicrin and lactucin from C. intybus are well known to improve inflammatory and metabolic functions. 25 To examine the inhibitory function of lactucin and lactucopicrin on various inflammasomes' activation, LPS-primed BMDMs were treated by lactucin and lactucopicrin at the nontoxic condition, followed by stimulation with inflammasome activators. Lactucin significantly inhibited ATP-induced IL-1β secretion and also slightly inhibited both nigericin- and silica-induced IL-1β secretion, but not AIM2 and NLRC4 inflammasome activation (Fig. 4B). Lactucopicrin did not inhibit NLRP3, AIM2, and NLRC4 inflammasome activation (Fig. 4C). Lactucin and other active constituents might act synergistically in regulating inflammasome activation, thereby improving T2D.

HPLC fingerprint analysis of CI.
Discussion
In the present investigation, we found that extract of aerial part of CI inhibited the NLRP3 inflammasome activation, but not NLRC4 and AIM2 inflammasome activation. IL-1β, released from NLRP3 inflammasome activation, was known to be implicated in the pathogenesis of T2D, and NLRP3 inflammasome component deficient mouse significantly improved both insulin sensitivity and glucose metabolism. 1 Recent study showed that omega-3 fatty acid prevented HFD-induced T2D through inhibition of NLRP3 inflammasome activation. 15 In the present study, CI treatment inhibited HFD-induced IL-1β production through inhibition of HFD-induced NLRP3 inflammasome activation. HFD-induced IL-1β has been reported to impair the insulin receptor signaling. 26 CI treatment might improve the HFD-induced insulin resistance through reduced HFD-induced IL-1β production.
During the progression to obesity in HFD-fed mice, a phenotypic switch occurs in adipose tissue macrophages from a M2 to an M1 phenotype, and these M1-type macrophages contribute to the development of insulin resistance. 16 NLRP3 has been reported to be important to HFD-induced phenotypic switch of macrophage from an M2 to an M1 phenotype, and in NLRP3-deficient mice, phenotypic switch of adipose tissue macrophages was attenuated in adipose tissue of HFD-fed mice compared to wild-type mice. 2 In the present study, CI treatment might inhibit HFD-induced phenotypic switch of macrophages through inhibited NLRP3 inflammasome activation. CI might improve glucose metabolism through inhibited metainflammation, such as phenotypic switch of macrophages.
Recent studies suggested that NLRP3 protein expression was regulated by autophagy-mediated degradation, and autophagy-mediated NLRP3 degradation could negatively regulate NLRP3 inflammasome activation. 17,18 Yan et al. reported that dopamine, a neurotransmitter, specifically regulated NLRP3 inflammasome activation through the autophagy-dependent NLRP3 degradation. 17 In the present study, CI slightly inhibited NLRP3 protein expression; this NLRP3 protein reduction by CI treatment was due to neither proteasomal degradation nor autophagy-dependent degradation. Although the exact mechanism for the downregulation of NLRP3 protein level by CI treatment is unknown, reduced NLRP3 protein level by CI treatment might contribute to the NLRP3 inflammasome inhibition.
In the process of NLRP3 inflammasome activation, NLRP3 inflammasome components are detected from MAM fraction. 20 ASC and its oligmerized forms are detected in Triton X-100 insoluble fraction in NLRP3 inflammasome-activated BMDMs. 21 NLRP3 inflammasome components are reported to be translocated from Triton X-100 soluble fraction to Triton X-100 insoluble fraction during NLRP3 inflammmasome activation, indicating that MAMs might be included in Triton X-100 insoluble fraction. CI treatment significantly inhibited the level of NLPR3 inflammasome components in Triton X-100 insoluble fraction, indicating that CI might inhibit NLRP3 inflammasome activation through translocation inhibition of NLRP3 inflammasome components to MAMs in NLRP3 inflammasome-activated BMDMs.
Several studies suggested that C. intybus contains a variety of phytochemicals, including sesquiterpene lactones, such as lactucin and lactucopicrin. 27 Lactucin and lactucopicrin have been reported to improve the inflammatory and metabolic functions. 25 In the present study, lactucin specifically inhibited NLRP3 inflammasome activation, but not AIM2 and NLRC4 inflammasome activation. Therefore, the inhibitory function of CI in NLRP3 inflammasome activation might be attributed to its active constituents, such as lactucin.
Collectively, our results demonstrate that CI ameliorated HFD-induced insulin resistance and metainflammation through the regulation of NLRP3 inflammasome activation. CI specifically inhibited NLRP3 inflammasome activation through both the inhibition of NLRP3 protein expression and inhibition of translocation of NLRP3 inflammasome components to Triton X-100 insoluble fraction. This is the first study to report that the aerial part of C. intybus exerts a modulatory effect on IL-1β secretion by attenuating NLRP3 inflammasome activation, leading to the amelioration of T2D.
Footnotes
Acknowledgments
This research was supported by the “Food Functionality Evaluation Program” under the Ministry of Agriculture, Food and Rural Affairs, and partly Korea Food Research Institute, and, in part, by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2014-A419-0015).
Author Disclosure Statement
No competing financial interests exist.