
Abstract
Bangle (Zingiber purpureum) is a tropical ginger that is used as a spice in Southeast Asia. Phenylbutenoid dimers isolated from Bangle have exhibited neurotrophic effects in primary cultured rat cortical neurons and PC12 cells. Furthermore, chronic treatment with phenylbutenoid dimers enhances hippocampal neurogenesis in olfactory bulbectomized mice. In this study, we investigated the effects of Bangle extract on behavior and hippocampal neurogenesis in vivo. SAMP8 mice, which are an established model for accelerated aging, with age-related learning and memory impairments, were given a Bangle-containing diet for 1 month, and subsequent behavioral tests and immunohistochemistry for Ki67, a proliferating cell marker, were performed. We found that the Bangle-containing diet improved spatial learning and memory deficits in the Morris water maze and significantly increased the numbers of Ki67-positive cells in the dentate gyrus of the SAMP8 mice. In addition, the Bangle extract exhibited a neurotrophin-like activity as indicated by the induction of neurite sprouting in PC12 cells. Our results suggest that Bangle is beneficial for the prevention of age-related progression of cognitive impairment.
Introduction
A
In contrast, neurotrophins, such as nerve growth factor (NGF), brain-derived neurotrophic factor, and basic fibroblast growth factor, have been recognized to play important roles in the promotion of neurogenesis, neurodifferentiation, neuroprotection, and neuroplasticity 3 –5 throughout the development of the vertebrate nervous systems. Thus, neurotrophins are expected to be potentially beneficial in the treatment of neurodegenerative diseases, including AD. 6 However, these proteins cannot cross the blood–brain barrier due to their high molecular weight and are easily decomposed by peptidases in physiological conditions. To address this issue, considerable efforts have been deployed to identify small molecules that exhibit the neurotrophic activity. 7 –12
Bangle is the rhizome of Zingiber purpureum, a tropical ginger that is widely distributed in Southeast Asia. This plant is used as a spice and is also applied in the traditional Indonesian medicine jamu. Bangle has also been utilized to treat fever, headaches, stomach pain, rheumatism, and obesity and serves as an ingredient in postpartum herbal medicine.
Bangle contains some compounds that have neurotrophin-like activities. For example, cis- and trans-3-(3′,4′-dimethoxyphenyl)-4-[(E)-3",4"-dimethoxystyryl]cyclohex-1-ene, phenylbutenoid dimers isolated from Bangle have been shown to exhibit neurotrophic effects that are characterized by neuritogenesis, neurite outgrowth promotion, and neuronal survival enhancement in primary cultured rat cortical neurons and PC12 cells. Furthermore, chronic treatment with these phenylbutenoid dimers enhances hippocampal neurogenesis in olfactory bulbectomized mice, which are an experimental model of depression and dementia. 13 Moreover, neocassumunarin A and B, which are components of Bangle, have shown NGF-potentiating effects in PC12 cells, and neocassumunarin A accelerates the prevention of Aβ42 aggregation in vitro. 14
These previous results suggest that Bangle may have the potential for development into a functional food or dietary supplement for preventing or delaying the progress of AD. However, the effects of Bangle extract on learning and memory-related behaviors in vivo have not been studied. Therefore, we investigated the effects of Bangle extract as a food additive on learning and memory-related behavior and hippocampal neurogenesis in the senescence-accelerated mouse prone-8 (SAMP8), which is a model for the early stage of AD. 15,16
Materials and Methods
Bangle extract
The sliced and dried rhizomes of Z. purpureum (500 g) were soaked in 70% ethanol (2 L) at room temperature for 3 days, and the mixture was filtered. The filtrate was concentrated and lyophilized under reduced pressure to obtain the resinous brown extract (46.1 g, 9.2% yield), which was dissolved in hot 90% ethanol (200 mL). After the addition of corn starch (220 g), this solution was evaporated under reduced pressure to yield a yellow powder (242.8 g), which was used as the Bangle extract for the present study. The HPLC chromatogram of the Bangle extract is shown in Figure 1. The quantitative analysis of the Bangle extract by HPLC revealed that the contents of cis- and trans-3-(3′,4′-dimethoxyphenyl)-4-[(E)-3",4"-dimethoxystyryl]cyclohex-1-ene, phenylbutenoid dimers, active constituents, were 0.63% and 0.30%, respectively.
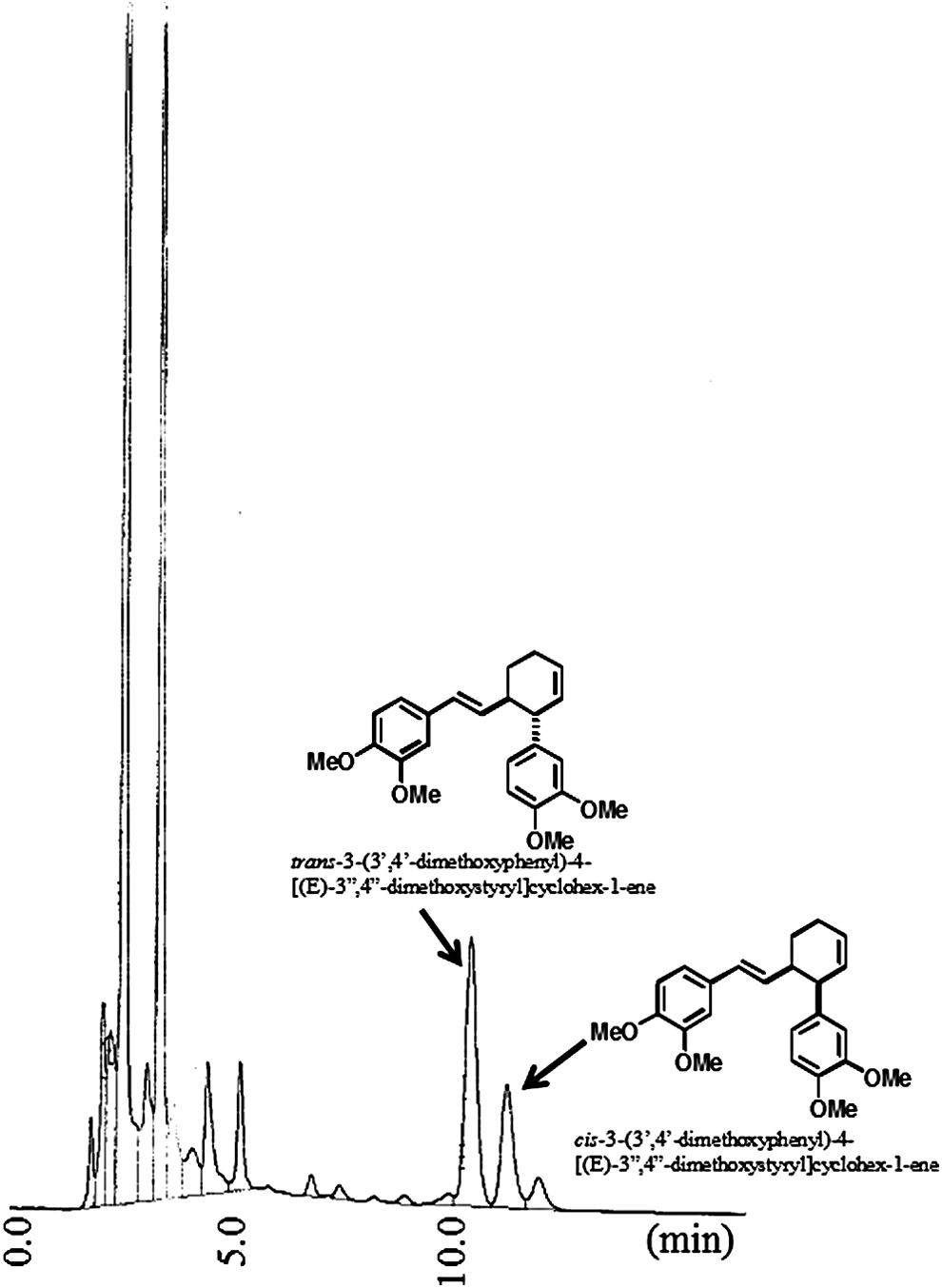
HPLC chromatogram of the Bangle extract.
HPLC conditions:
Nihon Bunko PU-2086 pump; column: TOSOH, ODS-100V, 5 μm; mobile phase: 80% methanol (1 mL/min).
Injection volume:
5 μL (the sample solution consisted of 41.8 mg Bangle extract in methanol 20 mL); detector: UV 254 nm.
Bangle-containing diet
Powder CE-2 (CLEA Japan, Inc., Tokyo, Japan) 60 g and 18.9% Bangle extract powder 238 mg (Bangle extract 45 mg) were passed through a 30-mesh sieve and each powder was mixed. The mixture was made muddy with ∼75 mL of purified water and molded into pellet form. The pellets were dried at 50°C under UV light irradiation overnight. The pellets were made every 2 weeks.
Animals
Male SAMP8 and senescence-resistant mouse prone-1 (SAMR1) were purchased from Japan SLC, Inc. (Hamamatsu, Japan). These mice were individually caged at 23°C ± 2°C and 50% ± 3% humidity. The animals were provided free access to food and water. After a 7-day recovery period, the 2-month-old SAMP8 (n = 12) were randomly divided into two groups: SAMP8 control (n = 6) and Bangle treated (n = 6). Age-matched SAMR1 (n = 6) served as the normal aging control. Bangle extract (Hosoda SHC Co. Ltd., Fukui, Japan) was orally administered in the diet (0.6 mg Bangle extract per gram of diet) for 1 month before the tests began.
All of the animal experiments were performed in accordance with the Guidelines for Animal Experimentation prepared by the Animal Care and Use Committee of Kochi University. All experiments were performed in a manner that minimized animal suffering.
Morris water maze
The Morris water maze was performed as described previously with minor modifications 17 to evaluate spatial learning and memory. A white circular tank (100 cm in diameter and 30 cm in height) was filled with water (25°C ± 1°C), made opaque with nontoxic white paint to a depth of 22 cm. Inside the tank, an escape platform (10 cm in diameter) was placed 1 cm below the water's surface.
Each mouse underwent daily training sessions of four trials for 6 consecutive days with the platform always placed in the same quadrant. In each trial, the mouse was placed into the water facing the wall at one of the three randomly selected standard start locations and then released. The mice that reached the platform within 60 sec were allowed to stay on the platform for 15 sec. If the mouse failed to find the platform within 60 sec, it was assisted and allowed to stay there for the same time.
All the trials were recorded with a camera located 106 cm above the water surface. The latencies were determined by measuring the time and distance that the mice required to reach the platform (Time MWM for Morris water maze; O'HARA & Co. Ltd., Tokyo, Japan). The time between trials was greater than 15 min.
Twenty-four hours after the final trial, the platform was removed, and the mice were given probe trials to test their memories of the hidden platform. The mice were allowed to swim freely for 60 sec. The time in the quadrant in which the platform was formerly located was measured.
Histology
The day after completing the behavioral tests, the mice were transcardially perfused with 2% paraformaldehyde in 0.1 M phosphate buffer (PB) (pH 7.4) under anesthesia. The brains were collected, postfixed at 4°C overnight, and cryoprotected with 25% sucrose containing 0.1 M PB at 4°C overnight. The 16-μm-thick coronal sections of the hippocampal regions were obtained using a cryostat (CM3050S; Leica, Bensheim, Germany) and mounted on MAS-coated glass slides (Matsunami Glass, Osaka, Japan). The slides were air-dried and washed with 0.1 M PB.
Immunohistochemistry was primarily performed as previously described. 18 The slide-mounted cryosections were air-dried, washed, and then blocked with 20% Block Ace (DS Pharma Biomedical, Osaka, Japan) in 0.1 M PB containing 0.01% saponin (Nacalai, Kyoto, Japan) for 30 min at room temperature in a humidified chamber. The sections were then incubated in 0.1 M PB containing 5% Block Ace and 0.01% saponin with a primary antibody against Ki67 (1:800 dilution) (ab15580; Abcam, Cambridge, United Kingdom) for 1 day at 4°C. The sections were washed five times with a wash buffer (0.1 M PB containing 0.005% saponin) for 10 min each and then incubated with Alexa Fluor 488-conjugated anti-rabbit IgG (1:800 dilution) (Life Technologies, Carlsbad, CA, USA) in a dilution buffer for 1 day at 4°C.
The sections were washed and covered with the VECTASHIELD mounting medium with DAPI (H-1200; Vector Laboratories, Burlingame, CA, USA). The sections were observed and photographed with a fluorescent microscope (BZ-9000; Keyence, Osaka, Japan).
Two sections per mouse were used, and one fluorescent image of the dentate gyrus (DG) region of the hippocampus was obtained from each section. The Ki67-positive cells were counted in each image. The mean value from two images was used for each mouse, and there were six mice in each group.
PC12 cells
The assays for the neuritogenic effects in the PC12 cells were performed according to previously described methods. 13
Statistical analysis
The results are expressed as mean ± SEM. Comparisons of the means were performed with analyses of variance (ANOVAs) or repeated-measure ANOVAs followed by Newman–Keuls post hoc tests. The criterion of P < .05 was considered significant.
Results
Bangle intake and body weights
The daily food consumptions per mouse did not differ between the SAMR1-control diet (4.26 ± 0.05 g/day), SAMP8-control diet (4.46 ± 0.05 g/day), and SAMP8-Bangle (4.13 ± 0.08 g/day) groups (day 7: F 2,15 = 0.13, P < .05; day 14: F 2,15 = 2.24, P < .05; day 21: F 2,15 = 3.14, P < .05; day 28: F 2,15 = 0.32, P < .05; day 35: F 2,15 = 0.39, P < .05) (Fig. 2A). The average daily dose of Bangle extract was calculated to be ∼110 mg/kg/day. There were differences in body weights between the SAMR1 and SAMP8 groups, with the exception of day 21, but there were no significant differences between the SAMP8-treated and SAMP8-Bangle-treated groups (day 7: F 2,15 = 5.56, P < .05, day 14: F 2,15 = 5.68, P < .05, day 21: F 2,15 = 3.54, P = .055, day 28: F 2,15 = 6.33, P < .05, day 35: F 2,15 = 6.55, P < .01) (Fig. 2B).
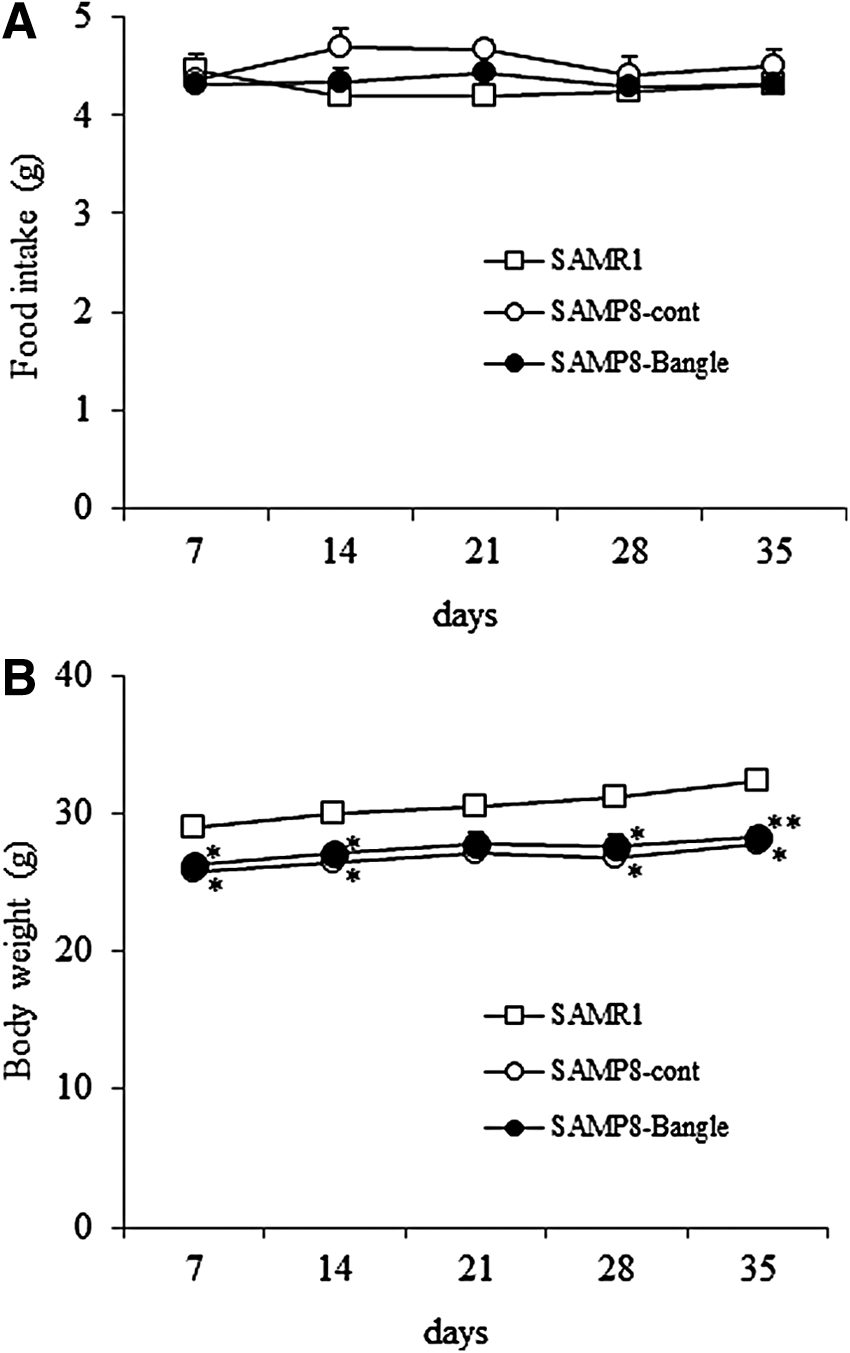
Food intake and changes in body weight. The results are given as mean ± SEM. □, SAMR1 control group; ○, SAMP8 control group; ●, SAMP8 Bangle-treated group.
The effects of Bangle on the spatial learning and memory of mice in the Morris water maze
To determine the effects of Bangle on learning and memory abilities, the mice were subjected to the Morris water maze. The swim speeds of the SAMP8 group were significantly slower than those of the SAMR1 group, and the Bangle treatment did not affect the swim speed in the SAMP8 group (Fig. 3A).
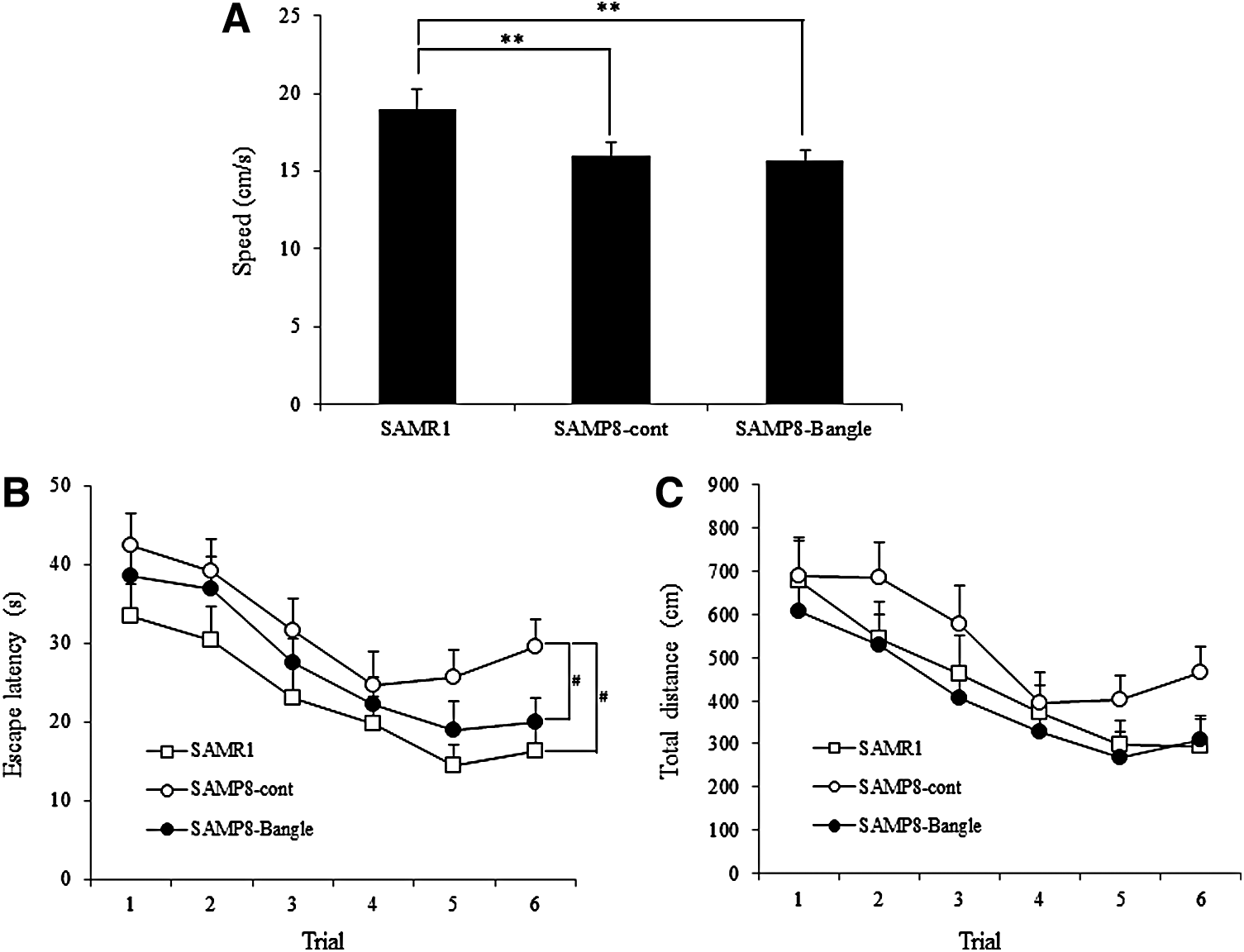
Behavioral changes in the hidden platform Morris water maze. The results are given as mean ± SEM. □, SAMR1 control group; ○, SAMP8 control group; ●, SAMP8 Bangle-treated group.
In the training session, two-way repeated ANOVAs for the escape latency (Fig. 3B) and the total distance (Fig. 3C) revealed effects of day (latency: F 5,345 = 12.70, P < .001, distance: F 5,345 = 11.08, P < .001) and group (latency: F 2,69 = 8.86, P < .001, distance: F 2,69 = 4.42, P < .05). However, there was no day × group interaction. Post hoc tests revealed that the escape latencies of the SAMR1 and SAMP8-Bangle treatment groups were significantly shorter than those of the SAMP8-control group on day 6.
In the probe trial in which the platform was removed (Fig. 4), the times spent in the trained quadrant were significantly greater than those spent in other quadrant in the SAMR1, but there were no significant differences in the time spent in each quadrant in the SAMP8-control group (Fig. 4B). Furthermore, the SAMP8 spent significantly less time in the trained quadrant than the SAMR1 mice (Fig. 4A). Similar to the SAMR1 group, the SAMP8-Bangle-treated groups spent significantly more time in the trained quadrant than in any other quadrant (Fig. 4B). Moreover, the Bangle treatment significantly increased the time spent in the trained quadrant (Fig. 4A).

Behavioral changes in the probe trial of the Morris water maze. The results are given as mean ± SEM.
The effect of Bangle on cellular proliferation in the DG of the hippocampus
To determine the effect of Bangle on hippocampal neurogenesis, immunohistochemistry for Ki67, a cell proliferation marker, was performed (Fig. 5A). Figure 5B shows the result of quantifying the Ki67-positive cells. There were no significant differences in the numbers of Ki67-positive cells in the subgranular zone (SGZ) of the DG of the hippocampus between the SAMP8-control and the SAMR1 groups at 3 months of age. However, the Bangle treatment significantly increased the number of Ki67-positive cells in the SAMP8 group compared with the SAMP8-control mice.
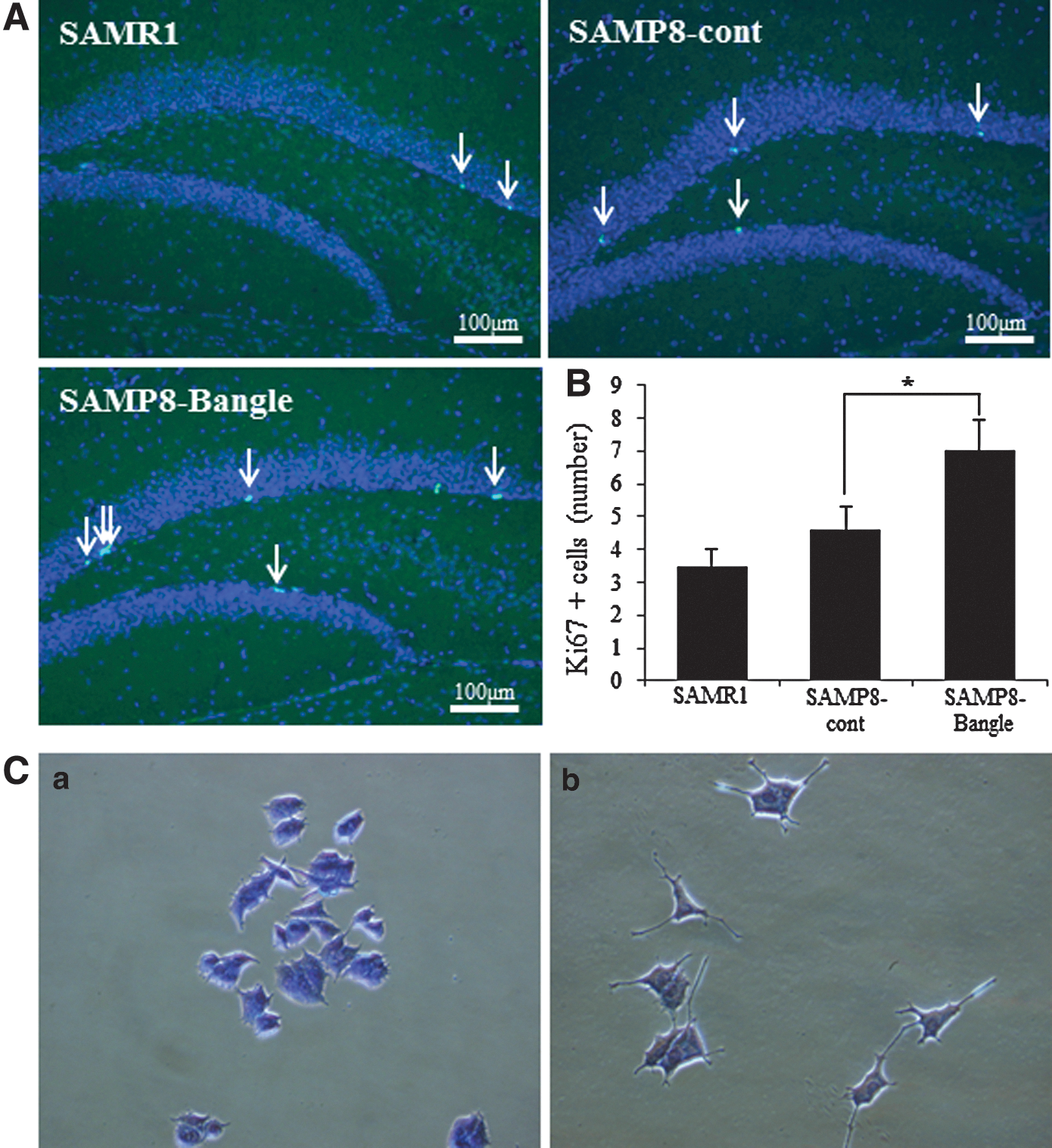
Neuritogenic effects on PC12 cells
In the control group (0.5% EtOH), few cells had neurites. Bangle (25 μg/mL) significantly induced neurite sprouting in the PC12 cells (Fig. 5C).
Discussion
The purpose of the present study was to evaluate the effects of Bangle extract on behavior and neurogenesis in vivo. The results revealed that the Bangle-containing diet improved the spatial learning and memory deficits and enhanced hippocampal cell proliferation in the SAMP8 mice.
Senescence-accelerated mice (SAM) are characterized by a rapid accumulation of senile features and a shortened life span. SAMP8 is recognized as a model of age-related AD because it exhibits early-onset deficits in learning and memory with characteristics that are similar to those of AD, such as cholinergic deficits, amyloid β (Aβ) deposition, and tau hyperphosphorylation. 16 Many reports have shown that SAMP8 exhibits learning and memory deficits from the age of 2 months 16 and exhibits increases in oxidative stressors, such as lipid hydroperoxide, in the brain beginning at 3 months of age. 19
However, the deposition of Aβ and histologic changes are not detectable in the brains of SAMP8 until at least 6 months of age. Therefore, 3-month-old SAMP8 mice are a model for early-stage AD with mild cognitive impairment and are suitable for evaluations of the preventive effects of supplements and drugs for the early stage of AD. Therefore, we determined the administration period from 2 months old as the onset of mild cognitive impairment to 3 months old as the evaluation period.
Moreover, the dose 100 mg/kg/day was determined assuming the realistic maximum dose when it is used as a dietary supplement for human. We found that oral supplementation with Bangle extract at this dose prevented the spatial learning and memory deficits of the 3-month-old SAMP8 mice in the Morris water maze. These results suggest that the consumption of Bangle from the early stages of AD is beneficial for preventing the age-related progression of cognitive impairment. However, a dose dependency study will be required in the future.
Meanwhile, we have already assessed the acute and chronic toxicity of the Bangle extract. The results show that the acute and chronic observed adverse effect levels of the Bangle extract are 2000 mg/kg and 1000 mg/kg for 90 days, respectively. In other words, the dose 100 mg/kg/day ensures both safety and efficacy of the Bangle extract.
Hippocampal neurogenesis is often thought to be necessary for hippocampus-dependent learning and memory. In the present study, we used Ki67 immunohistochemistry to evaluate cell proliferation. Ki67 is a general marker for proliferating cells but is not a specific marker for newly generated neurons. However, Ki67-positive cells in the SGZ of the DG at least partially reflect hippocampal neurogenesis.
Although it has been reported that neurogenesis is decreased in the hippocampus of SAMP8, 20 decreased numbers of Ki67-positive cells were not observed in the SGZ of the DG of the hippocampus of the 3-month-old SAMP8 mice relative to the SAMR1. However, we observed significant increases in the numbers of Ki67-positive cells in the SGZ of the DG of the Bangle-treated SAMP8 mice. Although the hippocampal neuronal loss in 8-month-old SAMP8 mice has been reported, 16 there were no significant neuronal losses in CA1, CA3, and DG of the hippocampus in 3-month-old SAMP8 mice compared with same aged SAMR1 in this study (data not shown) and other reports. 16
Hippocampal neurogenesis contributes to neural plasticity associated with learning and memory. Therefore, the data suggest that Bangle-induced hippocampal neurogenesis could improve learning and memory by enhancing neuronal plasticity, not by replenishing neuronal loss in young SAMP8. In addition, Bangle extract exhibits the neurotrophin-like activity as indicated by the induction of neurite sprouting in PC12 cells. These results strongly suggest that Bangle extract enhanced hippocampal neurogenesis through its neurotrophic effects in the SAMP8 mice.
The role of hippocampal neurogenesis in AD remains controversial. In many animal models of AD with familial-type mutations and in patients with severe AD, hippocampal neurogenesis is decreased 21 ; conversely, there are reports that neurogenesis is increased in some transgenic models of AD and humans with AD. However, these authors have hypothesized that neurogenesis is increased in the AD hippocampus due to a self-compensating mechanism for the replacement of neurons lost due to the disease. Therefore, the stimulation of hippocampal neurogenesis might provide a new treatment strategy for AD.
We have already identified all the compounds isolated from the Bangle extract, and have found that only the cis- and trans-3-(3′,4′-dimethoxyphenyl)-4-[(E)-3",4"-dimethoxystyryl]cyclohex-1-ene, phenylbutenoid dimers have exhibited neurogenesis in PC12 cells and neurite outgrowth promoting activity in the primary cultured rat cortical neurons. Thus, these phenylbutenoid dimers are most likely to be active constituents in the Bangle extract. In addition, the results of the present study are supported by ensuring that chronic treatment with the purified phenylbutenoid dimers also significantly increased the generation of new neurons as indicated by BrdU/NeuN double-labeled cells in the SGZ of the DG in olfactory bulbectomized mice. 13
Conclusion
This study indicated that chronic Bangle treatment improved the spatial learning and memory deficits by enhancing hippocampal neurogenesis in SAMP8 mice, which is a model for the early stage of AD. These results indicate that the regular consumption of Bangle may prevent age-related cognitive deficits, including those of the early stages of AD.
Footnotes
Acknowledgment
This work was supported by the Creation of New Business and Industry through Kochi Prefectural Industry-Academia-Government Collaboration Research Promotion Operations.
Author Disclosure Statement
No competing financial interests exist.