
Abstract
Sterol carrier protein-2 (SCP-2), which is not found in tissues of people with Zellweger syndrome, facilitates the movement of cholesterol within cells, resulting in abnormal accumulation of cholesterol in SCP-2-deficient cells. This study investigated whether synthetic high-density lipoprotein-like nanocarrier (sHDL-NC) improves the cellular transport of lysosomal cholesterol to plasma membrane in SCP-2-deficient fibroblasts. Human SCP-2-deficient fibroblasts were incubated with [3H-cholesterol]LDL as a source of cholesterol and sHDL-NC. The cells were fractionated by centrifugation permit tracking of [3H]-cholesterol from lysosome into plasma membrane. Furthermore, cellular content of cholesteryl ester as a storage form and mRNA expression of low-density lipoprotein (LDL) receptor were measured to support the cholesterol transport to plasma membrane. Incubation with sHDL-NC for 8 h significantly increased uptake of [3H]-cholesterol to lysosome by 53% and further enhanced the transport of [3H]-cholesterol to plasma membrane by 32%. Treatment with sHDL-NC significantly reduced cellular content of cholesteryl ester and increased mRNA expression of LDL receptor (LDL-R). In conclusion, sHDL-NC enables increased transport of lysosomal cholesterol to plasma membrane. In addition, these data were indirectly supported by decreased cellular content of cholesteryl ester and increased gene expression of LDL-R. Therefore, sHDL-NC may be a useful vehicle for transporting cholesterol, which may help to prevent accumulation of cholesterol in SCP-2-deficient fibroblasts.
Introduction
C
The known elements of cholesterol trafficking machinery include clathrin-coated pits, caveolae, Niemann–Pick disease protein, sterol carrier protein 2 (SCP-2), and microsomal triglyceride transfer protein. 7 Among them, SCP-2, known as a nonspecific lipid transfer protein, is a 13.3-kDa protein present in peroxisomes and facilitates the movement of cholesterol, phospholipids, fatty acids, and fatty acyl-CoA within cells. 5,8,9 In addition, SCP-2-deficient mice accumulate methyl-branched fatty acids, which suggest a physiological role for SCP-2 in α-fatty acid oxidation. 10 For reasons that are not yet clear, SCP-2 is not found in tissues from patients with Zellweger syndrome, a lethal autosomal recessive disorder of infancy. 9,11 Therefore, it has been suggested that certain carriers such as liposomes or lipoproteins could reduce the accumulation of cellular cholesterol to prevent the related diseases.
Nanotechnology has enabled the manipulation of the biological and physiological characteristics of materials to facilitate drug targeting and delivery. 12 Among them, bionanocarriers, including LDL and HDL, have drawn attention as therapeutic agents for delivering drugs or natural compounds. However, since it is difficult to isolate lipoproteins in large quantities and they are variable in size and composition, 12 gold nanocarriers coated with biochemical components may be useful as substitute of bionanocarriers. According to several studies, administration of synthetic HDL (sHDL) is associated with increased cholesterol efflux ability and is able to promote reverse cholesterol transport. 12 –14 Therefore, in the present study, we investigated whether the synthetic HDL-like nanocarrier (sHDL-NC) is able to accelerate the transport of LDL-derived lysosomal cholesterol to the plasma membrane in human SCP-2-deficient fibroblasts.
Materials and Methods
Cell culture
Human SCP-2-deficient fibroblasts were obtained from the Mutant Cell Repository (Camden, NJ, USA) and maintained in 5% CO2 at 37°C, in Dulbecco's modified Eagle's medium (DMEM) plus 10% (v/v) fetal calf serum. For analysis, the medium containing 10% fetal bovine serum deficient in lipoprotein (Biomedical Technologies, Inc., Stoughton, MA, USA) was supplemented to deliver [3H-cholesterol]LDL and sHDL-NC on the day of the experiment.
Preparation of [3H-cholesterol]LDL
LDL containing [3H]-cholesterol with phospholipids and apolipoprotein A1 was prepared by the cholate dialysis method after cumulative flotation ultracentrifugation according to James et al. 15 Briefly, LDL was incubated with radiolabeled [3H]-cholesterol (1 μCi/mL) for 16 h, and then the solution of [3H-cholesterol]LDL was adjusted to a density of 1.06 g/mL by the addition of solid KBr in a final volume of 5 mL. The solution was then transferred to a 13-mL centrifuge tube, overlayered with saline, and centrifuged at 50,000 g for 28 h in a Ti50 rotor (Thermo Scientific, Rockford, IL, USA). After centrifugation, the tube contents (∼0.4 mL) were fractionated, pooled, and dialyzed. Particle diameters were determined by electrophoresis on 3–40% nondenaturing gradient gels. The radioactivity was measured by liquid scintillation counting (Beckman Coulter, Brea, CA, USA). Aliquots of [3H-cholesterol]LDL were frozen at −80°C until use.
Synthesis of sHDL-NC
Unconjugated nanocarrier (UNC) with median size of 5 nm was purchased from Ted-Pella, Inc., (Redding, CA, USA). Two phospholipids were used to construct sHDL-NC. The phospholipid, 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC), was purchased from Sigma-Aldrich (Saint Louis, MO, USA) and 1,2-dipalmitoyl-sn-glycero-phosphoethanolamine-N-[3(2-pyridyldithio)propionate] (DiS) was purchased from Avanti Polar Lipids, Inc., (Alabaster, AI, USA). Human apolipoprotein A1 (ApoA1) was obtained from Meridian Life Sciences (Memphis, TN, USA). For preparation of sHDL-NC according to Tripathy et al., 14 unconjugated gold nanocarrier was incubated with diethylpyrocarbonate (DEPC) for 16 h, autoclaved at 121°C for 30 min, and passed through a 0.22-μm filter. The concentration of nanocarrier was determined using a UV-Vis spectrophotometer (Agilent Technologies, Inc., Santa Clara, CA, USA). Nanocarrier was incubated with a fivefold molar excess of ApoA1 for 4 h at room temperature with gentle shaking. Next, two phospholipids dissolved in ethanol were added in 250-fold molar excess to the solution of ApoA1 nanocarrier to complete sHDL-NC (Fig. 1). To purify sHDL-NC from unbound nanocarrier, the solution was centrifuged at 16,500 g for 40 min, and the pellet was resuspended in nuclease-free water. For control vehicle, UNC was used after measuring concentration by spectrophotometric readings.
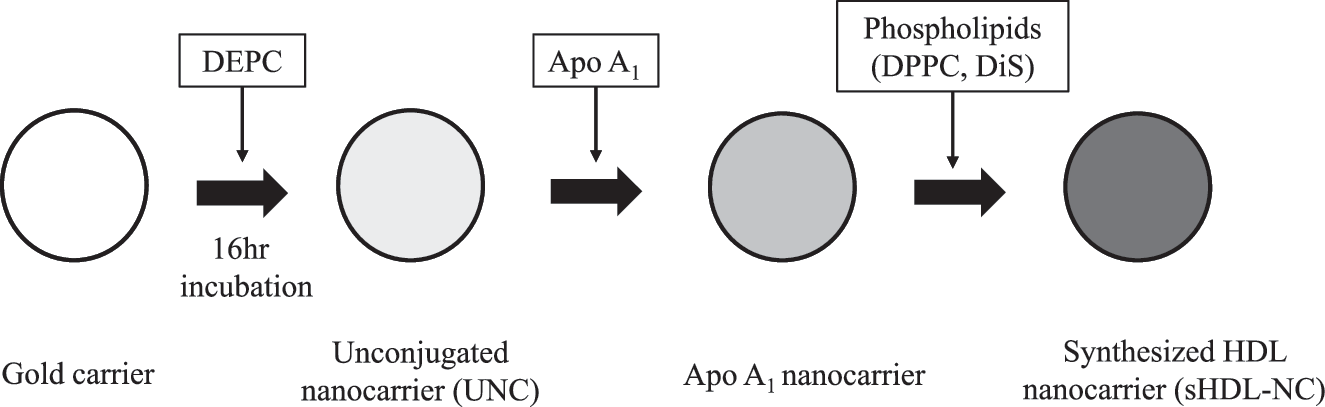
Postulated scheme for synthesis of synthetic high-density lipoprotein-like nanocarrier (sHDL-NC). Gold nanocarriers were surface coated with apolipoprotein A1 (ApoA1) and two phospholipids (DPPC and DiS).
Cell homogenization
At 80% confluency, the cells were washed thrice with phosphate-buffered saline without Ca2+ and Mg2+ and suspended in homogenate buffer containing 0.25 M sucrose, 1 M EDTA, and 5 mM Tris–HCl, pH 7.4, by scraping with a rubber policeman. A homogenate was prepared by up and down strokes with a Dounce homogenizer after adding 1 mL buffer in a T75 flask.
Fractionation of cellular components
At first, a total cellular homogenate supernatant (TCS) was obtained by centrifuging the cell homogenate for 5 min at 1000 g. Total membrane fraction (MF) was prepared from TCS by centrifugation for 1 h at 10,000 g, 4°C, and resuspending the pellet in 4 mL 0.25 M sucrose/EGTA. For density centrifugation on metrizamide gradients, the supernatant was layered onto a discontinuous gradient of 2 mL of 50%, 2 mL of 42%, 1 mL of 33%, 1 mL of 30%, 1 mL of 27%, 1 mL of 23%, 1 mL of 20%, and 1 mL of 17% (v/v) metrizamide in buffer containing 0.25 M sucrose, 3 mM imidazole, 1 mM EGTA, and 0.1% ethanol. The amount of plasma MF was determined by measuring alkaline phosphodiesterase activity adapted from Beaufay et al. 16 Lysosomal fraction (LF) was extracted from TCS using a commercial lysosome isolation kit (Sigma-Aldrich). For compensation, the amount of lysosome was determined by measuring β-N-glucosaminidase activity. The residue from TCS was considered as cytosolic fraction (CF).
Determination of cytosolic cholesteryl ester content
To determine the cytosolic cholesteryl ester content, the cells were incubated for 8 h after sHDL-NC treatment, and then the CF was obtained by the above method. Next, thin-layer chromatography was performed using an ITLC-SA plate. The development solvent was petroleum ether:toluene (v/v, 65:35) that provided good separation of cholesteryl ester (Rf values of 0.84, respectively). The plate was dried for at least 20 min in air and visualized by staining with iodine vapor. The amount of cholesteryl ester was enzymatically measured by the cholesteryl ester quantification kit (Abcam, San Francisco, CA, USA).
Real-Time PCR of LDL receptor
Total RNA from cells was isolated using the RNeasy mini kit (Qiagen, Valencia, CA, USA). Briefly, 1 μg of total RNA obtained from SCP-2-deficient fibroblast was incubated with reverse transcriptase (Invitrogen, Carlsbad, CA, USA) to produce cDNA. Real-time PCR was performed using selective primer sets with universal SYBR Green PCR Master Mix according to the manufacturer's instructions (Qiagen). The sequences of the sense and antisense primers used for amplification were as follows: GAPDH, 5′-CCATGAGAA GTATGACAACAGCC-3′ and 5′-TGGCAGGTTTTTCTA GACGG-3′; LDL receptor (LDL-R), 5′-GTGTGAAGATA TTGACGAGTG-3′ and 5′-AGTAGATTCTATTGTTGG TCA-3′. Data analyses were carried out using the 7500 System SDS software version 1.3.1 (Applied Biosystems, Inc., Foster City, CA, USA).
Statistical analysis
All values are expressed as mean ± SD and repeated a minimum of three times. Data were analyzed by the difference between means, and statistical significance was assessed by Student's t-test using SPSS PASW Statistic 20.0 (SPSS, Chicago, IL, USA).
Results and Discussion
Cellular uptake of [3H-cholesterol]LDL with no treatment of sHDL-NC
At first, to identify the transport of exogenous cholesterol to the plasma membrane, SCP-2-deficient fibroblasts were preincubated with [3H-cholesterol]LDL for 8 h, and then cellular uptake of [3H]-cholesterol in each fraction was measured using a liquid scintillation counter. As shown in Figure 2, [3H]-cholesterol was detected in all fractions of plasma membrane (MF), cytosol (CF), and lysosome (LF). In detail, the level of [3H]-cholesterol in CF and LF increased 2 h after incubation and reached a plateau at 6 h and continued to slowly increase to a maximal level at 8 h. However, MF [3H]-cholesterol increased quite slowly, indicating that most of [3H]-cholesterol was not efficiently transported into the plasma membrane due to lack of SCP-2. According to several studies, ∼80–90% of cellular cholesterol is located in the plasma membrane with the remainder distributed in other cellular MF, including endoplasmic reticulum, mitochondria, and Golgi apparatus. 17,18 Because cholesterol accumulation inside mammalian cells is detrimental to cholesterol deposition in atherosclerosis and cell survival, cellular cholesterol homeostasis is tightly regulated by balancing of the influx and efflux of cholesterol. 5,19 Thus, genetic mutations, including Niemann–Pick disease type C and Zellweger syndrome, affect the transport of cellular cholesterol. Several proteins have been proposed to contribute to intracellular sterol trafficking and distribution. SCP-2, also known as nonspecific lipid transfer protein, is conserved among bacteria, archaea, and eukaryotes. 20 Johnson et al. reported that SCP-2-deficient fibroblasts showed a modest decrease by ∼25% in cholesterol efflux compared with normal control cells. 21 In addition, fibroblasts transfected with antisense DNA to SCP-2 showed delayed transfer of newly synthesized cholesterol to the plasma membrane. 22 This fact may suggest that SCP-2 is not the only carrier of cellular cholesterol to regulate the transport of exogenous cholesterol to the plasma membrane. Consistent with our data, the level of cholesterol in the plasma membrane reaches a maximum through a slower cytoskeleton- and Golgi-dependent route in SCP-2-deficient human fibroblasts. 22 Our results also demonstrated that cholesterol transport into the plasma membrane with no treatment of sHDL-NC was slowly increased in SCP-2-deficient cells, supporting that SCP-2 deficiency partially influenced the cholesterol transport, but not enough to induce the related diseases through cholesterol accumulation. Therefore, an experimental approach for a substitute for SCP-2 could be useful for preventing cellular cholesterol accumulation.
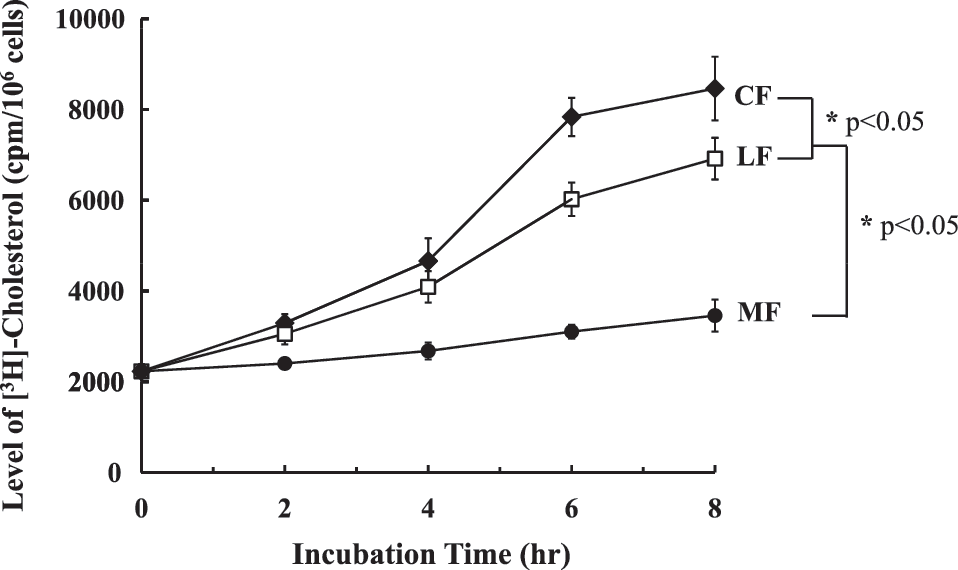
The cellular uptake and distribution of [3H]-cholesterol in each fraction of SCP-2-deficient fibroblasts with no treatment of sHDL-NC. The cells were incubated with 3 μg/mL [3H-cholesterol]LDL for 8 h after exposure. A significant difference was determined at P < .05 (n = 3). CF, cytoplasmic fraction; LF, lysosomal fraction; MF, plasma membrane fraction.
Enhancement of [3H]-cholesterol release from [3H-cholesterol]LDL in lysosomes treated with sHDL-NC
To examine whether treatment with sHDL-NC accelerates the hydrolysis of [3H]-cholesterol from [3H-cholesterol]LDL in lysosomes, we monitored the lysosomal [3H]-cholesterol by liquid scintillation counting. After 8 h of incubation with sHDL-NC, the level of lysosomal [3H]-cholesterol released from [3H-cholesterol]LDL was significantly increased by 53% compared with UNC control vehicle (Fig. 3). These results indicated that sHDL-NC treatment facilitates the uptake of [3H-cholesterol]LDL into lysosome and releases FC by hydrolysis of LDL particles.
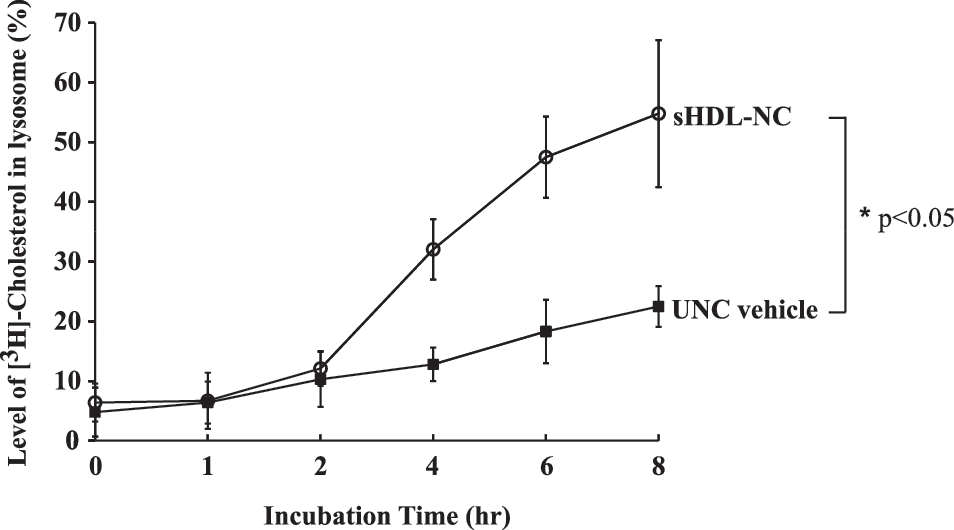
Level of [3H]-cholesterol in LF after treatment with sHDL-NC or unconjugated nanocarrier (UNC) in SCP-2-deficient fibroblasts. The cells were incubated with 1 μg/mL sHDL-NC or UNC for 8 h after exposure. [3H]-cholesterol was measured by liquid scintillation counter. A significant difference was determined at P < .05 (n = 3).
Transport of [3H]-cholesterol to plasma membrane with treatment of sHDL-NC
The movement of endogenously synthesized cholesterol from the endoplasmic reticulum to the plasma membrane appears to be distinct from movement of LDL-derived cholesterol, in which exogenous cholesterol is transported from lysosomes to the plasma membrane. 6 The majority of exogenous cholesterol and cholesteryl ester enters the cells through LDL-R endocytosis. In mammalian cells, LDL binds to its receptors at the cell surface and is recruited into clathrin-coated endocytotic vesicles. After endocytosis, LDL moves toward the lysosome where cholesteryl ester is hydrolyzed to unesterified cholesterol, and then FC rapidly transfers to the plasma membrane. 23 –26 SCP-2 expression in mammalian cells results in enhanced cholesterol transfer to the plasma membrane. 5,9,24 Baum et al. reported that transfection with SCP-2 cDNA in intact cells increased the proportion of cellular FC in the plasma membrane pool. 27 Because of the difficulty in applying SCP-2 cDNA transfection for human tissues, sHDL-NC in this study was added to SCP-2-deficient fibroblasts to stimulate cholesterol movement into the plasma membrane. To determine whether treatment with sHDL-NC could stimulate cholesterol transport from the lysosome to plasma membrane, SCP-2-deficient fibroblast was incubated with sHDL-NC for 8 h and the plasma membranes were fractionated. As shown in Figure 4, the transport of [3H]-cholesterol with UNC control vehicle was slightly increased by 11.4%, whereas sHDL-NC treatment significantly increased the level of [3H]-cholesterol in MF fraction, especially at 6h and 8 h by 28% and 32%, respectively, compared with UNC control vehicle.

Level of [3H]-cholesterol in plasma MF after treatment with sHDL-NC or UNC in SCP-2-deficient fibroblasts. The cells were incubated with 1 μg/mL sHDL-NC or UNC for 8 h after exposure. [3H]-cholesterol was measured by liquid scintillation counter. A significant difference was determined at P < .05 (n = 3).
Cellular cholesteryl ester content and LDL-R mRNA expression
The excess cholesterol in a cell is converted to cholesteryl ester by ACAT as a storage form or is removed from a cell by cellular cholesterol transport to the plasma membrane. 6,28 Therefore, it was necessary to determine whether increased transport of cholesterol to the plasma membrane by sHDL-NC affects the level of cytosolic cholesteryl ester. As shown in Figure 5, the level of cytosolic cholesteryl ester was increased by UNC vehicle in a time-dependent manner. However, treatment with sHDL-NC reduced the content of cholesteryl ester by 30.4%, indicating that sHDL-NC inhibited the accumulation of cholesteryl ester by increasing transport of cholesterol into the plasma membrane.
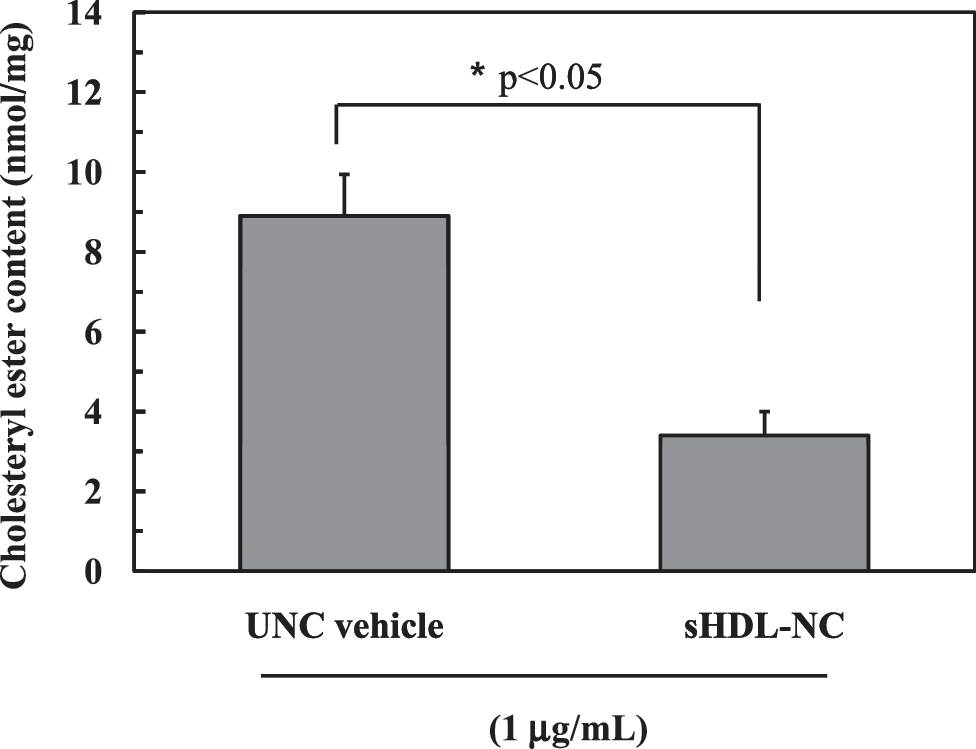
Level of cellular cholesteryl ester content after treatment with sHDL-NC or UNC. For chromatographic separations for cholesteryl ester, an ITLC-SA plate was used and further colorimetrically determined. A significant difference was determined at P < .05 (n = 3).
Gene expression of the LDL-R can be upregulated when the cellular cholesterol level is reduced due to transport into plasma membrane. As shown in Figure 6, treatment with sHDL-NC significantly increased the mRNA level of LDL-R by 42.3% at 8 h of incubation compared with UNC vehicle.
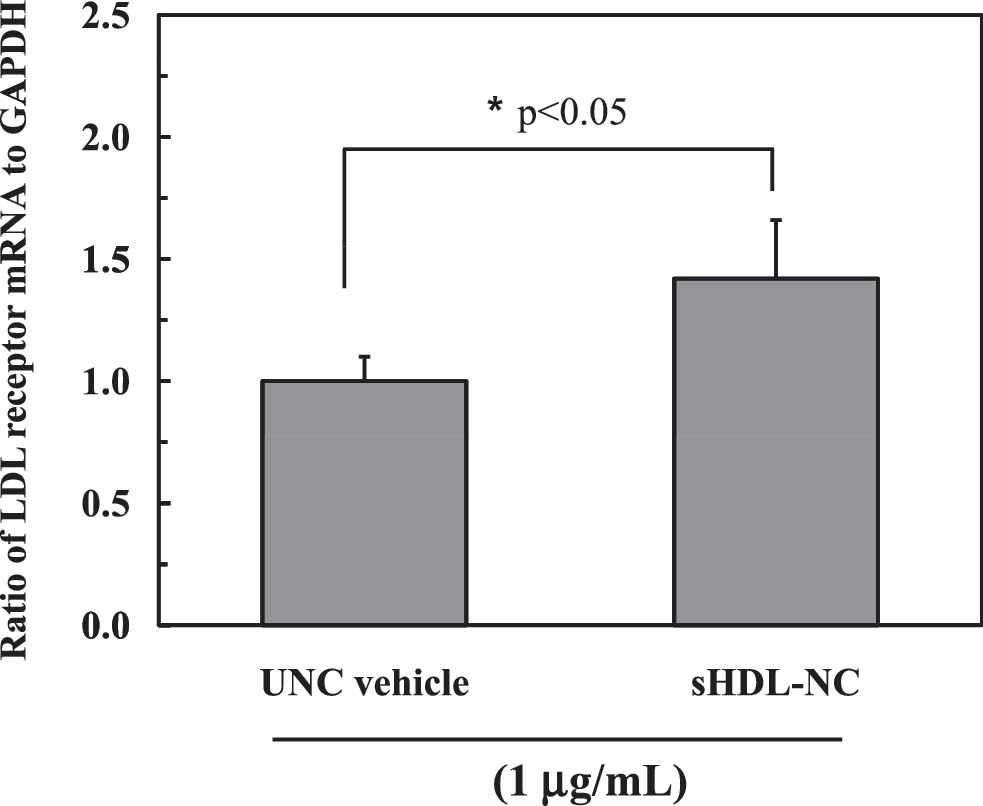
mRNA expression of low-density lipoprotein receptor (LDL-R) in SCP-deficient fibroblast after treatment with sHDL-NC or UNC. The data are shown as a relative ratio of LDLR mRNA expression to GAPDH. A significant difference was determined at P < .05 (n = 3).
Nanocarrier has attracted a great deal of interest in biological and physicochemical applications as a therapeutic or diagnostic agent. Lipoproteins, LDL and HDL, are spherical nanocarriers characterized by an insoluble core consisting of cholesterol and triacylglycerol surrounded by a shell of phospholipids and proteins. 29 They have a strong potential to act as a drug delivery carrier due to their small size and long residence time in circulation. 12 However, due to its difficulty in manipulation and storage, gold nanocarriers in encapsulation with phospholipids and apoprotein were recently used to mimic lipoprotein nanocarriers. In this study, sHDL-NC was constructed based on gold nanocarriers encapsulated with phospholipids and ApoA1 that is a component of HDL and promotes the transport of cholesterol from inside the cells to the membrane. To our knowledge, although nanocarriers have been applied to several research areas, this is the first trial for gold nanocarrier with ApoA1 to be treated on SCP-2-deficient fibroblast for facilitating cholesterol transport. Our data suggested that sHDL-NC enhanced the intracellular cholesterol transport from the lysosome to plasma membrane. In addition, treatment with sHDL-NC reduced the cellular content of cholesteryl ester, as a storage form of cholesterol, suggesting that sHDL-NC acted to inhibit the accumulation of cholesterol by transporting cholesterol to the plasma membrane, resulting in reduced conversion of cholesterol to cholesteryl ester. It is also supported by increased gene expression of LDL-R, which is upregulated when the cellular level of cholesterol and cholesteryl ester becomes lower than the normal level.
In summary, our results demonstrated that incubation with sHDL-NC enables increased transport of cholesterol from the lysosome to plasma membrane in SCP-2-deficient fibroblasts. It was demonstrated by tracing the radiolabeled cholesterol in each fraction of cytosol, lysosome, and plasma membrane. Furthermore, it was also indirectly evidenced by reduced cholesteryl ester content and increased gene expression of LDL-R. Taken together, sHDL-NC may be a useful vehicle for transporting cellular cholesterol to the plasma membrane, which may help to prevent accumulation of cholesterol in SCP-2-deficient tissues (Fig. 7). In future studies, it would be useful for sHDL-NC to be tested in an SCP-2-deficient animal model to identify further function in regulating the intracellular cholesterol level.

Postulated scheme for action mode of sHDL-NC to increase cholesterol transport to the plasma membrane and to decrease accumulation of cholesteryl ester in SCP-2-deficient fibroblast.
Footnotes
Acknowledgment
This work was supported by a grant from the Kyung Hee University, 20110255.
Author Disclosure Statement
No competing financial interests exist.