
Abstract
Citrus unshiu peel has been used empirically as a traditional medicine to improve bronchial asthma and blood circulation in northeast Asian nations, including Korea, Japan, and China. In this study, we report the proangiogenic effects of the aqueous extract of Citrus unshiu peel (AECUP). In human umbilical vein endothelial cells, AECUP significantly induced cellular migration and capillary tube formation. We also demonstrated that AECUP markedly increased the phosphorylation of FAK and ERK1/2 through the integrin signaling pathway. Additionally, we identified that narirutin and hesperidin were major constituents of AECUP and both showed proangiogenic effects, but at different levels. Collectively, these results suggest that the AECUP may have potential as a therapeutic agent for improving angiogenic functions with reduced harmful side effects.
Introduction
T
Angiogenesis is the formation of new blood vessels from existing endothelium. Angiogenesis is recognized to play a negative function in various physiological and pathological conditions, that is, tumors, rheumatoid arthritis, and ocular disorders. Therefore, blockade of angiogenesis is a critical goal of some therapeutic strategies. 7 Incessant efforts to develop novel antiangiogenic reagents have resulted in the approval of Avastin, a vascular endothelial growth factor (VEGF) neutralizing antibody, by the U.S. Food and Drug Administration regardless of the effectiveness of Avastin. 8 However, angiogenesis also plays an essential positive function in burns/scalds, wound healings, stroke, cardiac disorders, and various diabetes-related diseases. 9 Mutually incompatible functions of angiogenesis in human health reflect the complexity of the process mechanism in various diseases. It has been reported that basic fibroblast growth factor and VEGF are the most active inducers of angiogenesis both in vitro and in vivo. 10,11 However, their use in clinical applications is limited due to their high cost and short half-lives.
Satsuma mandarin (Citrus unshiu Marc.) peel has been used traditionally as a medicine to improve bronchial asthma and blood circulation. 12 This citrus unshiu peel contains various bioactive compounds such as phenolic acids and flavonoids. Hesperidin, naringin, and nobiletin are representative components of the Citrus unshiu peel. Recently, Citrus unshiu peel has been reported to inhibit allergic reactions, skin inflammation, oxidative stress, and tumor promotion. 13 –16 However, there have been no reports about the proangiogenic effect of the Citrus unshiu peel, despite the actual application of Citrus unshiu peel as a medicinal herb for blood circulation. Therefore, scientific evidence delineating the correct application of Citrus unshiu peel in angiogenesis is urgently needed.
In the present study, we report that aqueous extract of Citrus unshiu peel (AECUP) acts as a proangiogenic factor and we reveal the mechanism responsible for the proangiogenic effects of AECUP for the first time. Our results demonstrate that AECUP is an attractive proangiogenic material with its high safety and low cost, compared with existing angiogenic stimuli.
Materials and Methods
Preparation of AECUP
AECUP was prepared from the waste after citrus juice processing in Jeju Island. In brief, citrus peel waste was lyophilized, ground, and then the powder was suspended in 50 mM sodium acetate buffer (pH 4.8).
Cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from the American Type Culture Collection (ATCC, Manassa, VA, USA). HUVECs were grown in EGM-2 Bulletkit medium (Lonza Biologics, Hopkinton, MA, USA) at 37°C in a humidified atmosphere containing 5% CO2. All experiments were performed using HUVECs within 3–6 passages.
Reagents
Antibodies for phospho-focal adhesion kinase (FAK) (Y397), FAK, phospho-AKT (S473), AKT, phospho-ERK (T202/Y204), and ERK were from Cell Signaling Technology (Beverly, MA, USA). Anti phospho-Tyr (Y99) and GAPDH antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Inhibitors of MEK1/2 (U0126), FAK (SC203950), and negative control of MEK1/2 (U0124) were purchased from Santa Cruz Biotechnology. Recombinant VEGF-A was purchased from R&D Systems (Minneapolis, MN, USA).
Cell proliferation
Cell proliferation was measured using a WST-1 assay (Boehringer Mannheim, Mannheim, Germany) as previously described. 17 To assess cell viability, the absorbance was measured at 450 nm using a microplate reader (Bio-Rad, Richmond, CA, USA).
Cell migration
Cell migration assays were performed as previously described. 18 In brief, after an 18-h starvation in EBM-2 with 0.2% FBS, HUVECs (2 × 104 cells/well) were suspended in 200 μL serum-free EBM containing different dosages of AECUP and loaded into each upper chamber. After six hours, cells on the upper surface of the filter were removed with a cotton swab. Filters were fixed with 4% paraformaldehyde and stained with 1% crystal violet solution. The absorbance of the eluted dye was measured at 560 nm in an ELISA reader (Bio-Rad, Richmond, CA, USA).
Tube formation
Tube formation assays were performed as previously described with some modifications. 19 In brief, 250 μL growth factor-reduced Matrigel (BD Biosciences, San Jose, CA, USA) was used to coat 24-well plates (SPL Life Sciences, Pocheon, Korea) and allowed to polymerize at 37°C for 30 min. HUVECs (3 × 104 cells/well) were suspended in 500 μL serum-free EBM medium containing different dosages of AECUP. After incubation for 16 h at 37°C, photographs of four representative fields per well were taken using phase-contrast microscopy. Endothelial tubes were quantified by counting the number of junctions defined as the origin of two or more branch protrusions.
High-performance liquid chromatography and LC/MS analysis
The Shimadzu high-performance liquid chromatography (HPLC) system (Shimadzu Scientific Instruments, Columbia, MD, USA), equipped with a photodiode array detector and a Luna C18(2) column (5 μm particle size, 4.6 mm ×250 nm, Phenomenex, Torrance, CA, USA), was used for HPLC analysis as follows: 20% acetonitrile at 0 min, 40% acetonitrile at 10 min, 70% acetonitrile at 20 min, 70% acetonitrile at 25 min, 20% acetonitrile at 27 min, and 20% acetonitrile at 30 min at a column flow rate of 1 mL/min. Detection of the peaks was made at 270 min. The LC-MS/MS analysis was performed using an Accela HPLC system coupled to a Thermo LXQ ion trap mass spectrometer with ESI negative interface (Thermo Scientific, Waltham, MA, USA).
Western blot analysis
HUVECs were seeded in six-well plates (Nunc, Roskilde, Denmark). After 48 h, the cells were starved in EBM-2 medium for 15 h before stimulation with the time-dependent AECUP. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used to ensure equal protein loading. The bands were measured by densitometry using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All data are expressed as the means ± standard error of the means (SEM). Levels of significance for comparisons between two independent samples were determined using Student's t-test. P values < .05 were considered statistically significant. Data are presented as mean ± standard deviation (SD). The level of significance for comparison between two independent samples was determined using Student's t-tests. Groups were compared using a one-way analysis of variance (ANOVA) with Tukey's post hoc test for significant main effects (SPSS 12.0K for Windows; SPSS, Inc., Chicago, IL, USA).
Results
Aqueous extracts of citrus unshiu peel induce endothelial cell proliferation, migration, and tube formation
To investigate the functional effects of AECUP in HUVECs, we first examined HUVEC proliferation under AECUP treatment or VEGF-A treatment as a positive control. 20 As shown in Figure 1A, treatments with 2.5 or 5 mg/mL of AECUP resulted in weak, but statistically significant, increases in the proliferation of HUVECs as the treatment dosages increased (2.5 mg/mL, 7.00% ± 0.03%; 5 mg/mL, 11% ± 0.01%). Since endothelial cell migration and morphogenesis are required for angiogenesis, we next examined the effects of AECUP and VEGF-A on HUVEC migration and tube formation using a transwell assay and capillary-like tubular structure forming on the Matrigel, respectively. Treatment with 2.5 mg/mL AECUP induced HUVEC migration, and with 5 mg/mL AECUP, to a higher degree compared with the control and exogenous 10 ng/mL VEGF-A treatment (Fig. 1B). Quantitative evaluation of tube formation by counting the junctions of branches revealed that exposure to AECUP (5 mg/mL) significantly increased the number of junctions of the tubular structure compared with control and exogenous VEGF-A treatment (Fig. 1C, D).
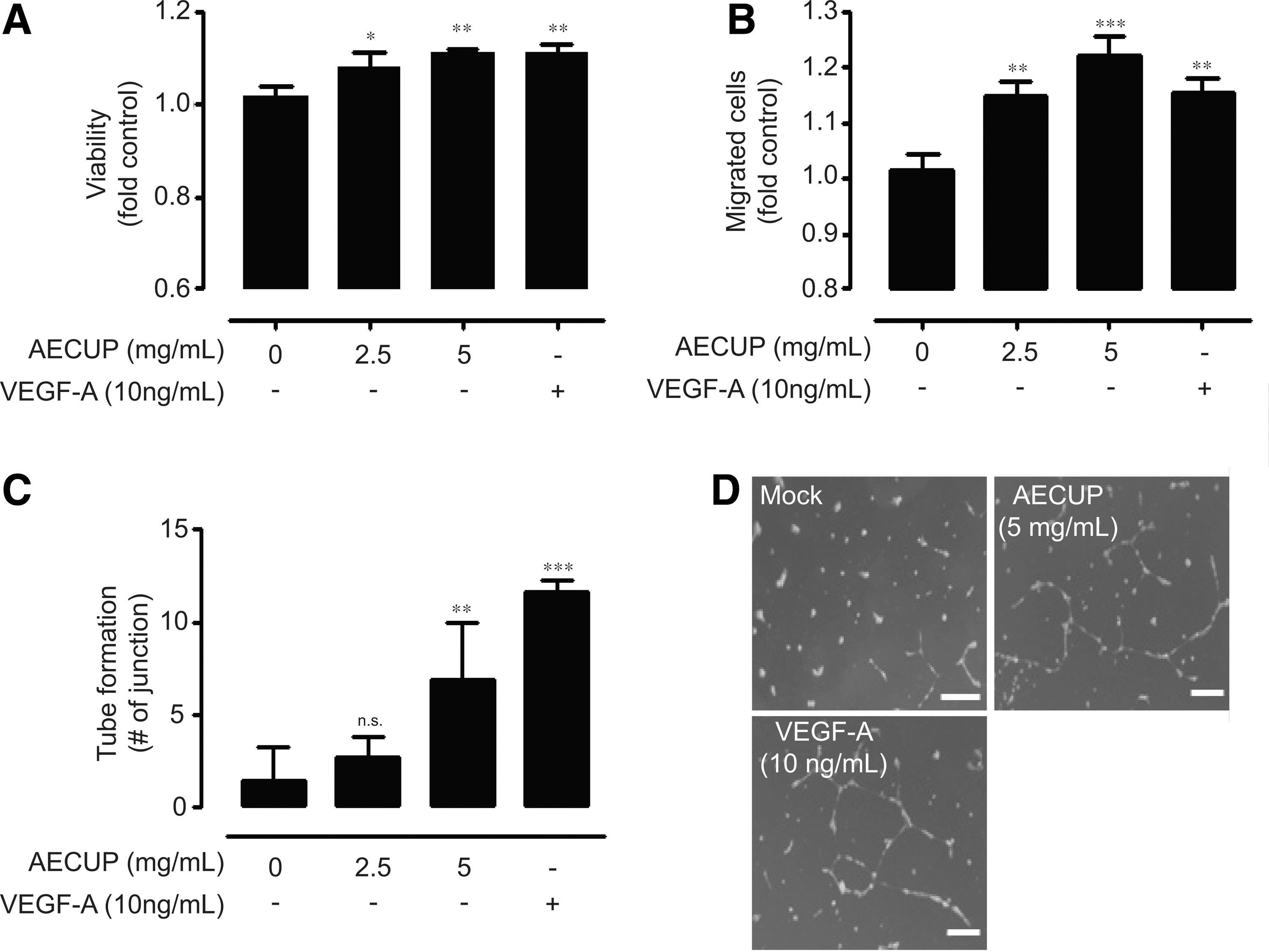
Proangiogenic effects of AECUP.
Aqueous extracts of Citrus unshiu peel activate the FAK and ERK1/2 phosphorylation through integrins
Proangiogenic phenotypes of HUVECs are controlled by various signaling pathways related to protein tyrosine kinases. 21 –23 To understand the proangiogenic mechanism of functional AECUP in HUVECs, we examined the time course of AECUP-induced tyrosine-phosphorylated signal transduction. Treatments with AECUP (5 mg/mL) time dependently increased tyrosine-phosphorylated proteins, particularly of molecular sizes approximately 60 and 120 kDa in HUVECs. Phosphorylation of a 120 kDa protein increased to a peak level at 10 min after stimulation and declined thereafter (Fig. 2A left and right). According to previous reports, FAK, PI3K/AKT, and ERK1/2 are the critical molecules in angiogenesis. 19,24 To further analyze the detailed mechanism of AECUP treatment, phosphorylated levels of FAK, AKT, and ERK1/2 were examined under the same conditions. Exogenous treatment with AECUP increased the phosphorylation of FAK and ERK1/2 (Fig. 2B left and right). In contrast, the same treatment caused no significant increase of AKT in HUVECs (data not shown). Several studies have shown that the activation of VEGF receptors or integrins is involved in endothelial cell proliferation, migration, and morphogenesis. 25,26 Therefore, we further investigated the involvement of integrins in AECUP-induced signal transduction. Exogenous treatment with AECUP increased the phosphorylation of FAK, whereas pretreatment with Arg-Gly-Asp (RGD) peptide (an inhibitor of the integrin–ligand interaction) clearly inhibited the phosphorylation of FAK in a dose-dependent manner (Fig. 2C left and right).
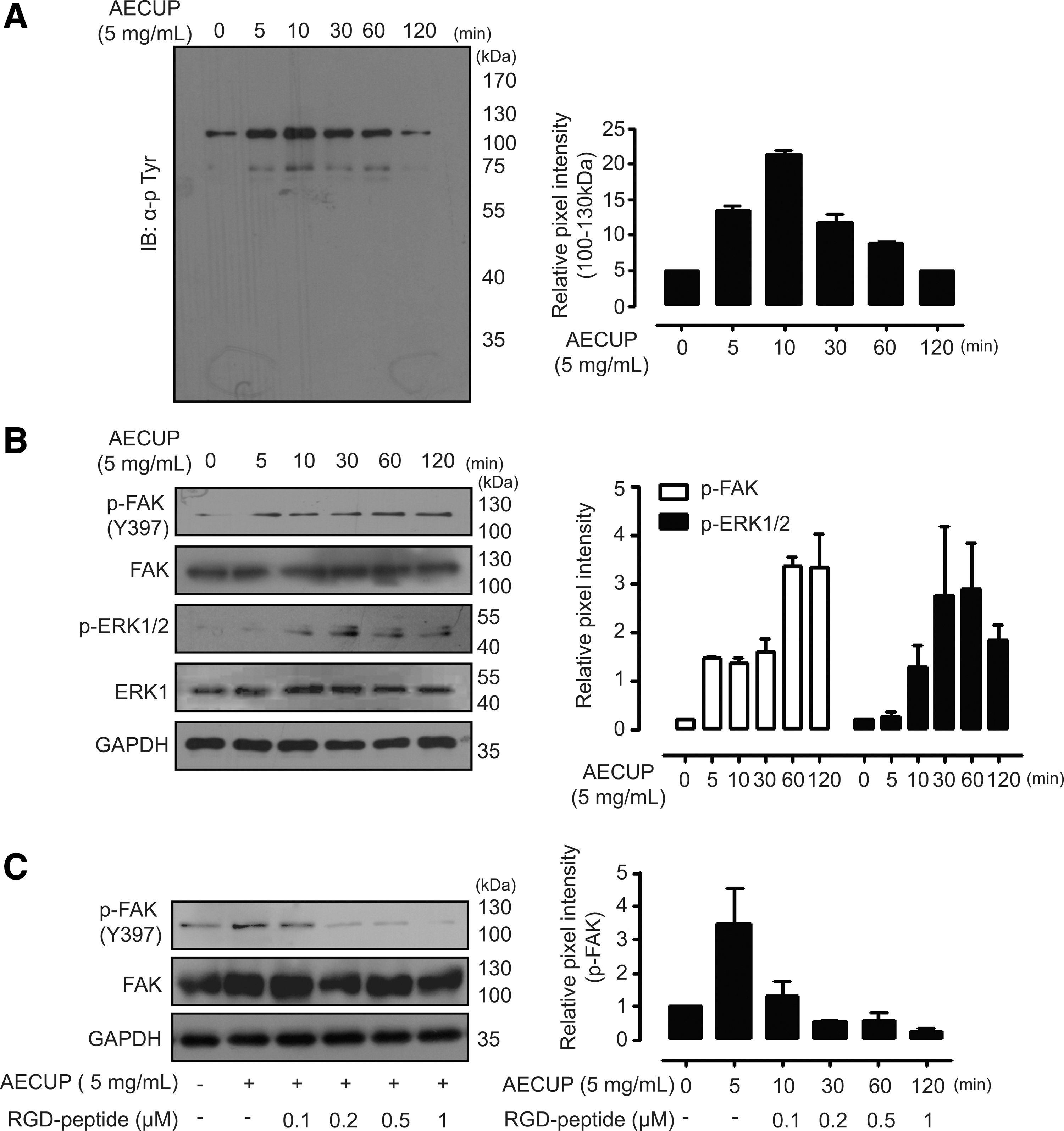
Involvement of FAK and ERK1/2 in AECUP-induced proangiogenic effects
Next, we assessed the involvement of FAK and ERK1/2 signaling in AECUP-induced proangiogenic effects using various pharmacological inhibitors; FAK inhibitor SC203950, MEK1/2 inhibitor U0126, and an inactive structural analog of MEK1/2 U0124. Pretreatment of HUVECs with SC203950 or U0126 dose-dependently inhibited AECUP-induced HUVEC proliferation. However, U0124 treatment had no effect on HUVEC proliferation under the same conditions (Fig. 3A). In particular, pretreatment with SC203950 or U0126 markedly inhibited HUVEC migration compared with proliferation (Fig. 3B). In addition, pretreatment with SC203950 or U0126 significantly inhibited AECUP-induced tube formation. Treatment of U0124 also had no effect on tube formation under the same conditions (Fig. 3c). These results indicate that AECUP mediates the proangiogenic effects of HUVECs through the FAK and ERK signaling pathway involved in integrin–ligand interactions.
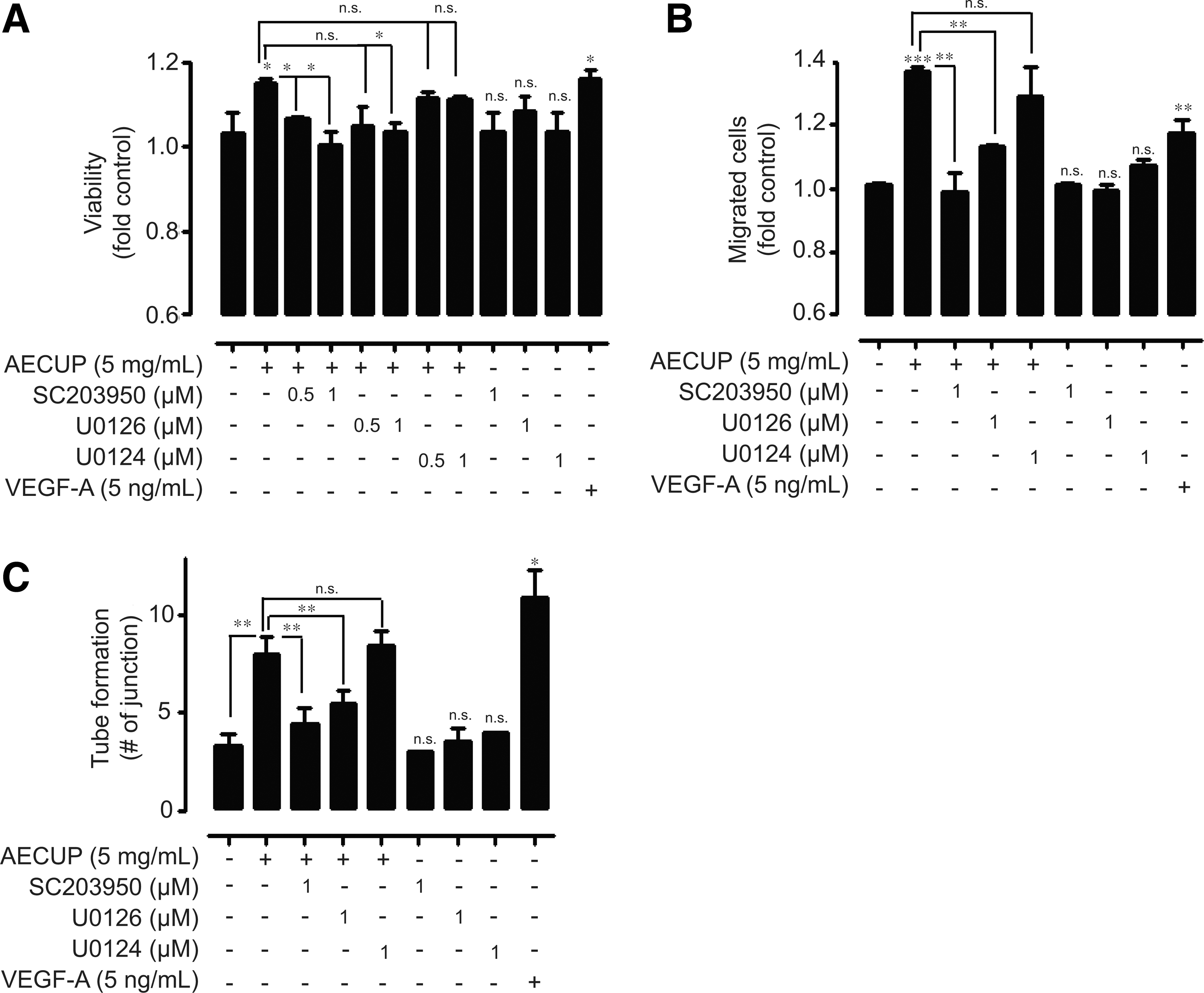
Involvement of FAK and ERK1/2 in aqueous extraction of citrus unshiu peel-induced proangiogenic effects
Characterization of functional components in aqueous extracts of Citrus unshiu peel
The flavonoid components of AECUP were characterized using HPLC and LC/MS analysis. The HPLC chromatograms of two major components (a and b) evidenced good resolution (Fig. 4A). LC/MS analysis determined the molecular weights of these two major components in the ESI negative ion condition (compound a; 579.05, compound b; 609.13), (Fig. 4B, C). Comparison of the molecular weights with established databases suggested that compound a may be narirutin (mw; 580.54) and compound b, hesperidin (mw; 610.56), respectively. These two components, narirutin and hesperidin, in AECUP were identified by the retention time comparisons with a coinjection analysis (AECUP mixed narirutin or AECUP mixed hesperidin), respectively (Fig. 4D left and right).
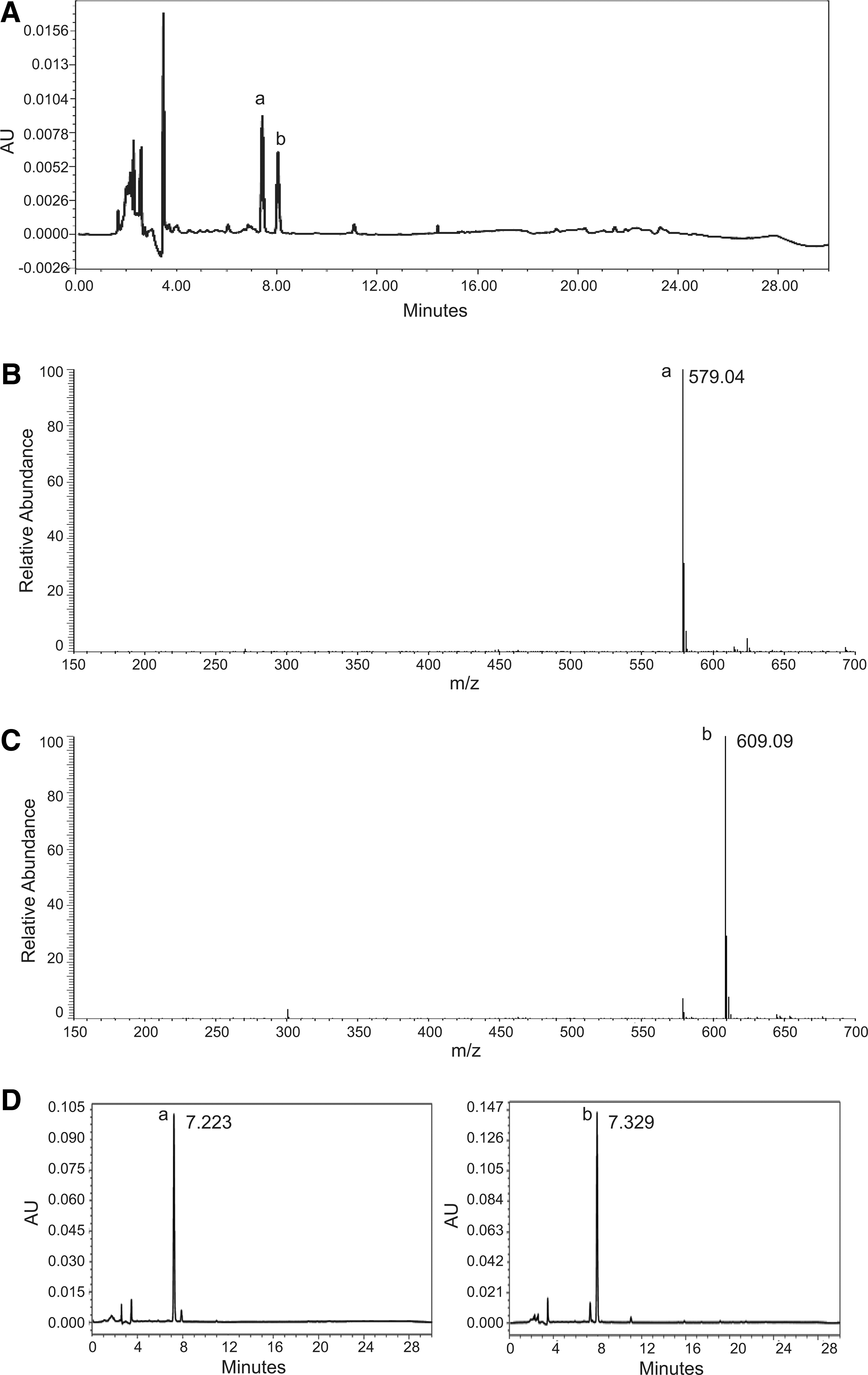
Identification of constituent elements in AECUP.
Involvement of hesperidin and narirutin in proangiogenic effects by aqueous extracts of Citrus unshiu peel
We next tried to identify which component of AECUP exerted the proangiogenic activity. Narirutin and hesperidin had simultaneously weak and similar effects on HUVEC proliferation (data not shown). With respect to migration, treatment of hesperidin had a particularly striking effect on HUVEC migration compared with narirutin treatment. Additionally, pretreatment of HUVECs with SC203950 or U0126 significantly inhibited hesperidin- or narirutin-induced HUVEC migration (Fig. 5A left and right). Similar results were also obtained using the tube formation assay (Fig. 5B left and right). To analyze the involvement of hesperidin as the major component of proangiogenic effects by AECUP, we examined the phosphorylated levels of FAK signaling under the RGD peptide pretreatment. Exogenous treatment with hesperidin increased the phosphorylation of FAK, and pretreatment with RGD peptide significantly inhibited hesperidin-induced FAK phosphorylation at the treatment dosages (Fig. 5C left and right).
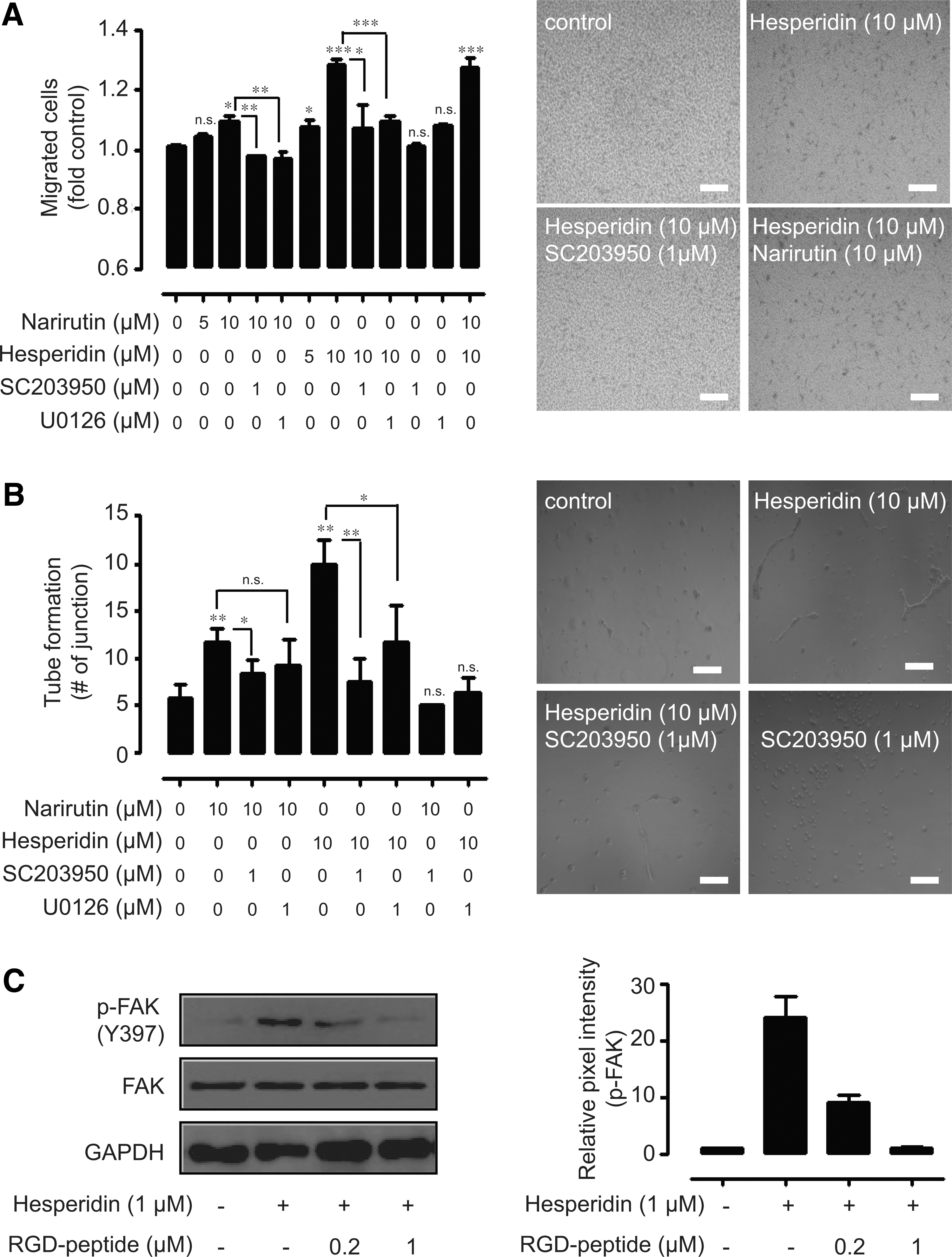
Involvement of hesperidin and narirutin in AECUP-induced proangiogenic effects
Discussion
Various proangiogenic reagents, including VEGF, hepatocyte growth factor, placental growth factor, and fibroblast growth factor, have been suggested as strong biopharmaceuticals for angiogenic therapy. 27,28 However, their use in clinical applications is limited due to the high cost. Following this line of thought, various natural substances seem to be attractive substitutes as they may have high therapeutic efficacy and low price, in addition to few side effects. In the present study, we examined functional roles of AECUP in HUVEC proliferation, migration, and tube formation. Additionally, we demonstrated that AECUP induced the activation of the FAK and ERK1/2 signaling pathway that is involved with integrins and their ligand interactions. Furthermore, we identified narirutin and hesperidin as components of AECUP; especially, hesperidin was found to have more proangiogenic activity than narirutin.
In previous reports, citrus peel has been shown to have an inhibitory effect in allergic reactions, skin inflammation, oxidative stress, and tumor development and to have a contradictory promotional effect in vascular functions, according to effects of various flavonoids present within it. 13 –16,29 This may, in part, be due to the cellular environment and/or to the variety of functions of the flavonoids. 30 In particular, we observed an in vitro proangiogenic effect of AECUP through HUVEC proliferation, migration, and tube formation for the first time. Especially, the AECUP effect on migration and tube formation was significant compared with those on proliferation in HUVECs.
Several studies have demonstrated that FAK and ERK activation are critical for angiogenesis. FAK and ERK have been reported as key molecules for endothelial cell migration and tube formation through various signaling pathways, including the integrin-related signaling pathway. 19,23,24,31 –33 Consistent with previous reports, we demonstrated that treatment with AECUP induced Y397-FAK and ERK1/2 phosphorylation in HUVECs, and AECUP-induced Y397-FAK phosphorylation was abolished by RGD peptide treatment that inhibited the interaction between the integrins and their ligands. It has been known that phosphorylation of the tyrosine 397 residue in FAK is responsible for cellular migration and tube formation. 34 –36 Accordingly, our results revealed that the pharmacological FAK inhibitor, SC203950, effectively inhibited the proangiogenic effects in HUVECs.
In agreement with previous reports, we identified that functional aqueous extracts of citrus unshiu peel consisted of narirutin and hesperidin. 12,37 Significantly, our results revealed that hesperidin exerted its proangiogenic effects predominantly through the FAK, ERK1/2, and integrin signaling pathway. Hesperidin has been reported to have antioxidant and anti-inflammatory effects in parallel with the improvement of vascular functions through its antiangiogenic effects. 38 –41 It is likely that the hesperidin and narirutin dynamically function in various cell types with a variety of outcomes, such as the medicinal herb ginseng, which contain both proangiogenic and antiangiogenic compounds. 42,43
Collectively, our findings provide the first pharmacological rationale for the use of AECUP as an angiogenesis inducer and suggest that AECUP might be considered as a potential therapeutic strategy for vascular-related diseases.
Footnotes
Acknowledgments
This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2009-0094059). We are grateful to Sustainable Agriculture Research Institute (SARI) in Jeju National University for providing the experimental facilities.
Author Disclosure Statement
There are no potential conflicts of interest.