
Abstract
This study was performed to investigate the antiobesity effect of germinated waxy black rice (GWBR) in high-fat diet (HFD)-induced obese mice. The mice were divided into a normal diet (ND) group, HFD group, and 2 test groups for 8 weeks: 2.5% GWBR-supplemented (GWBR-2.5) group and 5% GWBR-supplemented (GWBR-5) group. Supplementing with GWBR significantly reduced body weight gain and lipid accumulation in the liver and adipose tissue compared to the HFD control group. Triglyceride (TG), total cholesterol, and low-density lipoprotein-cholesterol levels in serum were decreased by GWBR supplementation, whereas high-density lipoprotein-cholesterol level significantly increased. In addition, mRNA levels of transcriptional factors, such as peroxisome proliferator-activated receptor-γ, CCAAT enhancer-binding protein (C/EBP)-α, C/EBP-β, sterol regulatory element-binding protein-1c, and related genes, including adipocyte fatty acid-binding protein, fatty acid synthase, and lipoprotein lipase, were significantly lower in the GWBR groups. However, lipolytic enzymes, such as hormone-sensitive lipase, adipose TG lipase, and carnitine palmitoyltransferase-1, and uncoupling protein 2 mRNA levels were significantly higher in GWBR-supplemented mice. These results suggest that GWBR exerts antiobesity effects by decreasing lipid accumulation and promoting lipolysis in HFD-induced obese mice.
Introduction
A
To treat and regulate obesity, with methods other than diet therapy and exercise, many different approaches, such as drugs and dietary supplements for weight loss or appetite suppression, have been suggested to date. However, some of the treatments have undesirable side effects, including valvular heart disease, vomiting, and headache. 5 Thus, numerous studies have participated in the search for antiobesity phytonutrients. 6,7
Black rice has been regarded as a beneficial functional food for its abundant content of fiber, tocopherols, oryzanols, vitamin B complex, and anthocyanins. 8,9 Especially, anthocyanins are a class of water-soluble flavonoids that are abundant in various fruits and vegetables, such as grapes, berries, and red cabbage. 10 Recent studies demonstrate that black rice and anthocyanin extract from black rice can attenuate hyperlipidemia and hepatic steatosis by fatty acid oxidation in high-fat diet (HFD) animal models. 9,10
In our previous study, germinated waxy black rice (GWBR) inhibited lipid accumulation using Oil Red O dye in 3T3-L1 adipocytes. 11 To the best of the author's knowledge, the effects of GWBR in an animal model on obesity have not yet been demonstrated. Therefore, the present study was performed to examine the antiobesity effect of GWBR supplementation for 7 weeks on body weight gain, serum lipid profiles, histological changes of adipose tissue and liver, lipase expression, adipogenic gene expression, and fatty acid oxidation-related gene expression in HFD-induced obese mice.
Materials and Methods
Preparation of GWBR
GWBR and waxy black rice (WBR) were harvested in Hamyang, Korea, in October 2010. To induce germination, WBR was soaked for 2–3 days in water and dried at 60°C. The dried GWBR was cut into small pieces and ground. The GWBR powder was stored at −20°C until used.
Animals and diets
Four-week-old male C57BL/6J mice (Orient Bio, Inc., Seongnam, Korea) were individually housed in standard cages in a controlled environment at 22°C ± 0.5°C and relative humidity of 50% ± 5% with a 12-h light–12-h dark cycle. Following the 1-week adaptation period, the mice were divided into 4 dietary groups of 10 mice each: a normal diet (ND) group, an HFD group, HFD +2.5% GWBR-supplemented (GWBR-2.5) group, and HFD +5% GWBR-supplemented (GWBR-5) group. GWBR was mixed homogeneously into the powdered food as appropriate for each treatment, and all food was provided in solid form. The components of the diet are described in Table 1. Animals were maintained on the feeding program for 8 weeks, and body weight and food intake were recorded every week throughout the experimental period. The care and treatment of the animals conformed to the Korea University Guidelines for the Ethical Treatment of Laboratory Animals (KUIACUC-2015–188).
GWBR, germinated waxy black rice; GWBR-2.5, high-fat diet + GWBR-2.5%; GWBR-5, high-fat diet + GWBR-5%; HFD, high-fat diet; ND, normal diet.
Preparation of adipose tissue, serum, and liver
At the end of the study, animals were fasted for 12 h and then sacrificed. The epididymal adipose tissue and liver were removed, weighed, and stored at −70°C until analysis. The serum was separated from the blood by centrifuging the sample at 700 g and 4°C for 15 min.
Biochemical analysis
Serum TG and total cholesterol (TC) concentrations were measured using commercial kits (Cayman Chemical, Ann Arbor, MI, USA). Low-density lipoprotein (LDL)-cholesterol and high-density lipoprotein (HDL)-cholesterol concentrations were also measured using commercial kits (Roche, Indianapolis, IN, USA). Alanine transaminase (ALT) and aspartate transaminase (AST) were determined enzymatically using commercial kits (Roche). Hepatic TG and TC concentrations were analyzed with the same commercial kits used for serum analysis.
Epididymal adipose tissue and liver histology
Epididymal adipose tissue and liver were carefully removed and immediately fixed with 10% (v/v) buffered formalin and embedded in paraffin. Sections of 3 μm were cut, and each section was stained with hematoxylin and eosin (H&E). All sections were examined by optical microscopy (Olympus BX50; Olympus Optical Ltd., Tokyo, Japan) and were printed at a final magnification of 100×. The average size of adipocytes was measured using the image analysis software (Image-Pro® Plus version 3.01; Media Cybernetics, Inc., Atlanta, GA, USA). The area of lipid accumulation in the liver was measured in randomly selected liver cells (50 cells) by the image analysis software.
Reverse transcription–polymerase chain reaction analysis
Total RNA was isolated from epididymal adipose tissue and liver of each mouse using an easy-BLUE™ Total RNA Extraction Kit (iNtRON Biotechnology, Seongnam, Korea) according to the manufacturer's protocol. Subsequently, 4 μg RNA was used for synthesis of cDNA. All primer sequences for polymerase chain reaction (PCR) are shown in Table 2. The amplification cycles were carried out at 95°C for 5 min, 95°C for 30 sec, 52–63°C for 30 sec, and 72°C for 30 sec (40 cycles). The PCR products were separated on a 1.5% agarose gel containing ethidium bromide (0.5 μg/mL) and visualized by UV light (Gel Doc 2000 and the Quantity One program; Bio-Rad, Sydney, Australia).
RT-PCR, reverse transcription–polymerase chain reaction.
Western blot analysis
Epididymal adipose tissue and liver were homogenized in modified RIPA buffer (50 mM Tris–HCl [pH 7.4], 1% Triton X-100, 0.2% sodium dodecyl sulfate [SDS], 1 mM PMSF, and 1 mM protease inhibitors). Insoluble material was removed by centrifugation for 20 min at 13,000 g at 4°C. The protein concentration of the supernatants was determined by a Bradford protein assay using bovine serum albumin as a standard. Aliquots (50 μg) of protein were separated by SDS-polyacrylamide gel electrophoresis, transferred to nitrocellulose membrane, and blotted with antiperoxisome proliferator-activated receptor-γ (PPAR-γ), CCAAT enhancer-binding protein-α (C/EBP)-α, sterol regulatory element-binding protein-1c (SREBP-1c) (Cell Signaling Technology, Beverley, MA, USA), and β-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The nitrocellulose membranes were developed using commercial chemiluminescence kits. Band intensities were quantified by optical densitometry.
Statistical analysis
Data are presented as mean ± SD (n = 10). The statistical analysis was performed using the SAS statistical software package (SAS Institute, Cary, NC, USA). Differences between means of individual groups were assessed by Tukey's multiple range test. Differences with P < .05 were considered significant.
Results
Body weights and food intake
The body weight gain (g/d) in the HFD control group was 1.5-fold greater than that in the ND group (Table 3). GWBR supplementation significantly reduced the body weight gain by 20% (GWBR-2.5) and 28% (GWBR-5) compared to the HFD control group. Food intake did not significantly differ among the HFD groups. However, food efficiency ratio (FER) in the GWBR-5 group was significantly lower than that in the HFD control group.
FER was estimated as the body weight gain for experimental period per food intake. Values are expressed as mean ± SEM (n = 8). Values in a column with different superscripts are significantly different (P < .05).
FER, food efficiency ratio.
Weights of adipose tissue and liver
The weights of adipose tissue (epididymal, subcutaneous, visceral, and retroperitoneal) in the HFD control group were significantly higher by 1.21- to 1.62-fold compared to the ND group (Table 3). However, the weights of adipose tissue in the GWBR-5 group were significantly lower by 22.8–41.5% compared to the HFD control group. In addition, liver weight (g/100 g body weight) was higher in the HFD control group than in the ND group, and liver weights were not significantly different among the groups.
Lipid profiles in serum and liver
The effects of GWBR on serum and liver lipid levels are shown in Table 4. Serum TG and TC levels in the HFD group were significantly elevated compared to the ND group. However, supplementing with GWBR for 8 weeks dose dependently decreased serum TG and TC levels in the HFD mice. Final serum TG levels in the GWBR-2.5 and GWBR-5 groups were 267.34 and 182.77 mg/dL, respectively, and both these were significantly lower than that in the HFD control group (343.11 mg/dL). Furthermore, final serum TC levels in the GWBR-5 group were 40% lower than those in the HFD control group. LDL-cholesterol was also significantly increased in the HFD control group compared to the ND group, but GWBR dose dependently lowers it by 22% and 44% in the GWBR-2.5 and GWBR-5 groups, respectively, compared with those in the HFD control group. In contrast, HDL-cholesterol levels in the GWBR-5 group significantly increased by 18% compared to the HFD control group. Hepatic TG and TC levels in the HFD control group were significantly higher in the ND group by 2.1- and 1.76-fold, respectively. Hepatic TG and TC levels were significantly lower by 56% and 52% in the GWBR-5 group, respectively, compared to the HFD control group.
Values are expressed as mean ± SEM (n = 8). Values in a column with different superscripts are significantly different (P < .05).
ALT, alanine transaminase; AST, aspartate transaminase; HDL, high-density lipoprotein; LDL, low-density lipoprotein.
Adipose tissue and liver histopathology
We examined the effects of GWBR extract on adipocyte diameter in HFD fed mice for 8 weeks. The adipocytes in epididymal adipose tissues in the HFD control group were dramatically larger than those in the ND group. Supplementing with GWBR resulted in dose dependently smaller adipocytes compared to the HFD control group (Fig. 1A). As shown in Figure 1B, the diameters of adipocytes in the GWBR-2.5 and GWBR-5 groups were 93.2 and 82.3 μm, respectively, and both these were significantly smaller than those in the HFD control group (95.3 μm). Histological examinations of the livers revealed that the HFD control group had notably large hepatic droplets, whereas lipid droplets in the GWBR-supplemented group were clearly fewer than that in the HFD control group (Fig. 2A). As shown in Figure 2B, hepatic lipid accumulation was significantly less in the GWBR-supplemented group compared to the HFD control group.
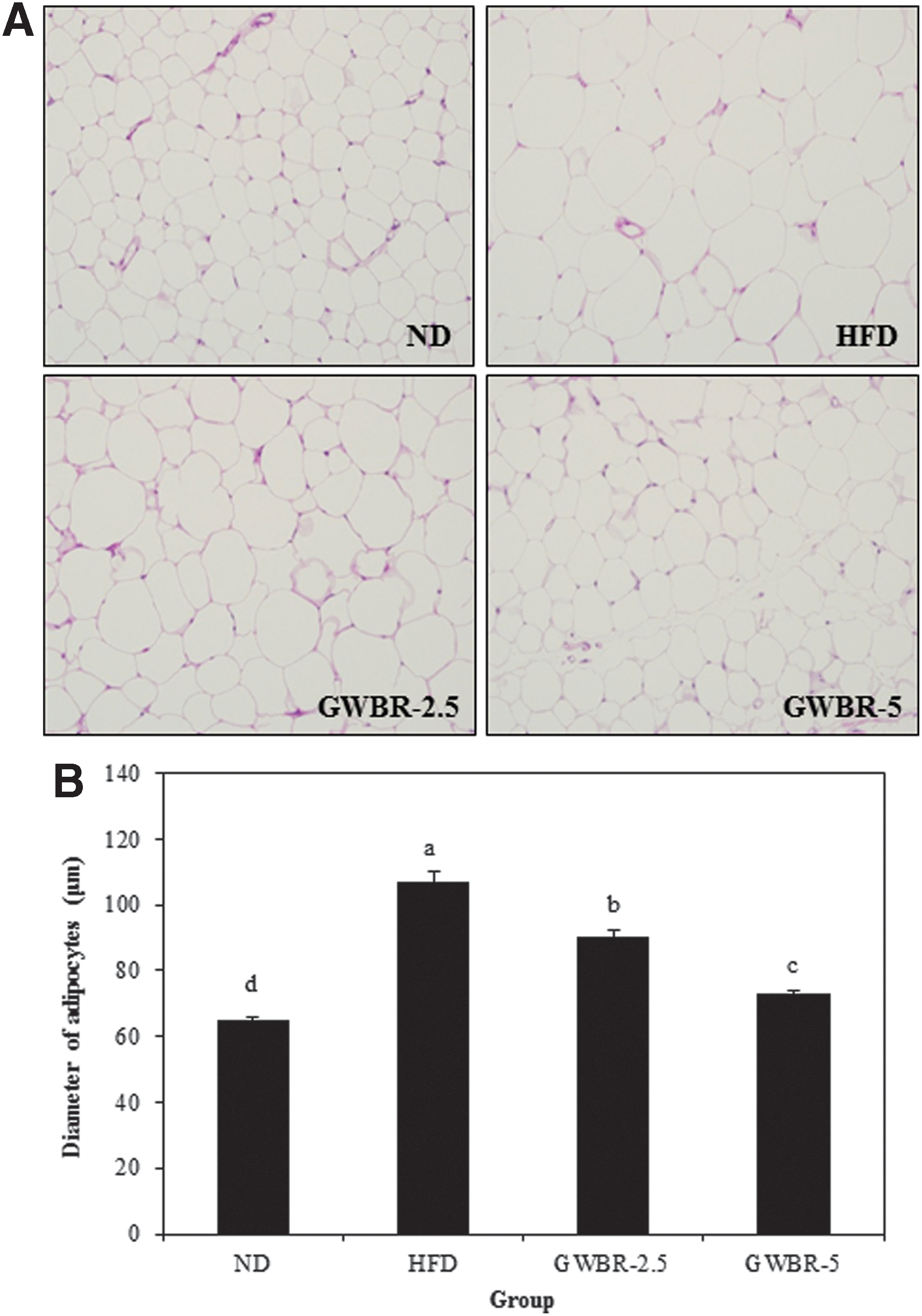
Histological changes in adipose tissue in high-fat diet-induced obese mice. Histology of adipose tissue
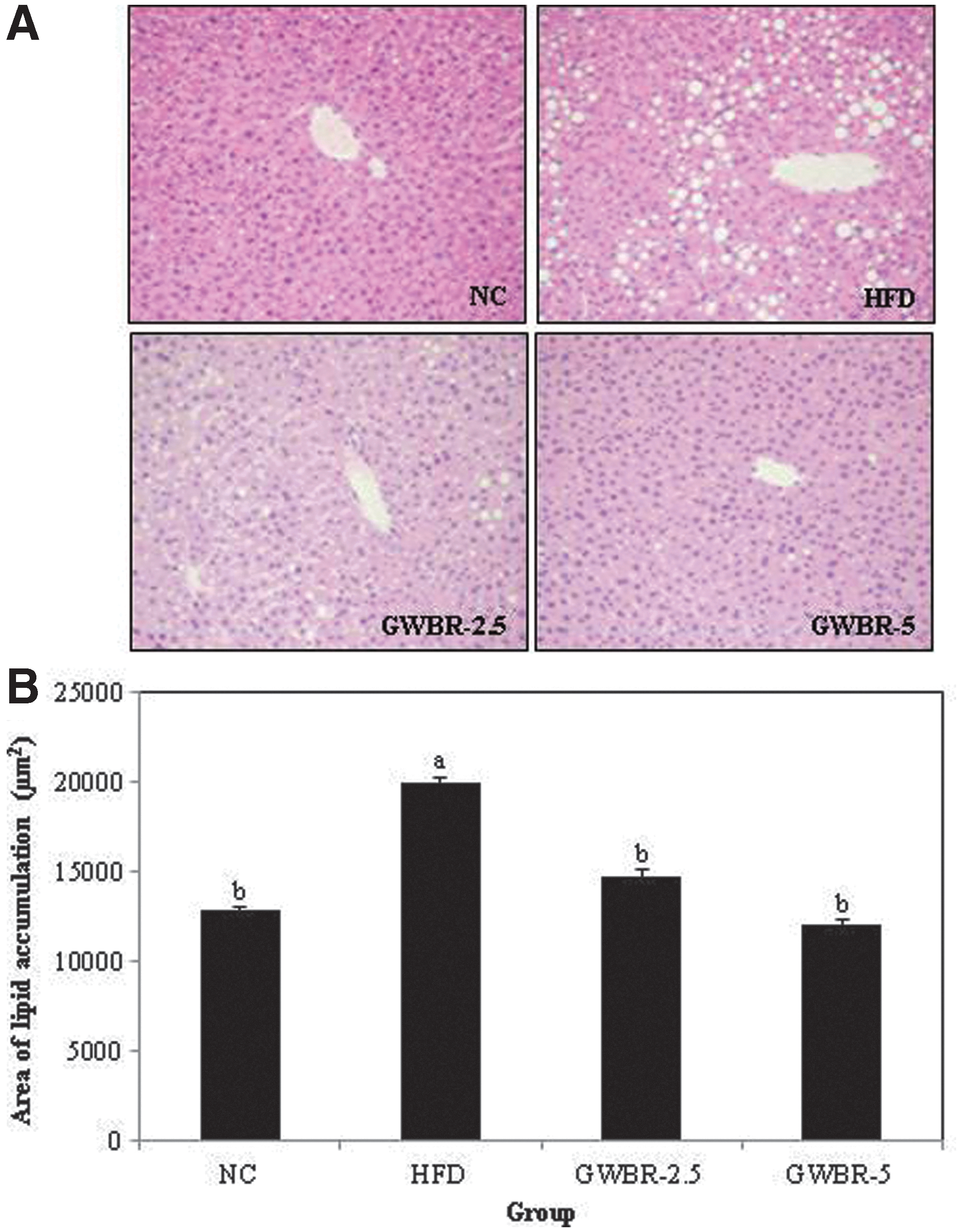
Histological changes in liver in high-fat diet-induced obese mice. Histology of liver
mRNA expression of lipogenic-related genes
As shown in Figure 3A, the adipogenic transcription factors, C/EBP-α and PPAR-γ, mRNA expressions in the adipose tissue were significantly lower in the groups supplemented with GWBR compared to the HFD control group. We also examined the PPAR-γ target gene, aP2. GWBR supplementation resulted in dose dependently lower aP2 mRNA expression by 7% and 46% in the GWBR-2.5 and GWBR-5 groups, respectively, compared to the HFD control group. In addition, PPAR-γ and SREBP-1c mRNA expressions in the liver were significantly lower after 5% GWBR supplementation by 34% and 66%, respectively, compared to the HFD control group (Fig. 3B). The mRNA expression of fatty acid synthase (FAS), a key enzyme of fatty acid synthesis, was significantly higher in the HFD control group than in the ND group. However, GWBR supplementation resulted in significantly lower FAS mRNA expression by 30% compared to the HFD control group. These data indicate that GWBR supplementation inhibited lipogenesis through FAS expression by suppressing the expression of adipogenic transcription factor in the liver. In addition, the protein levels of adipogenic transcription factors (C/EBP-α, PPAR-γ, and SREBP-1c) were significantly higher in adipose tissue and liver from the HFD control group compared with the ND group and significantly lower with GWBR supplementation (Fig. 3C).
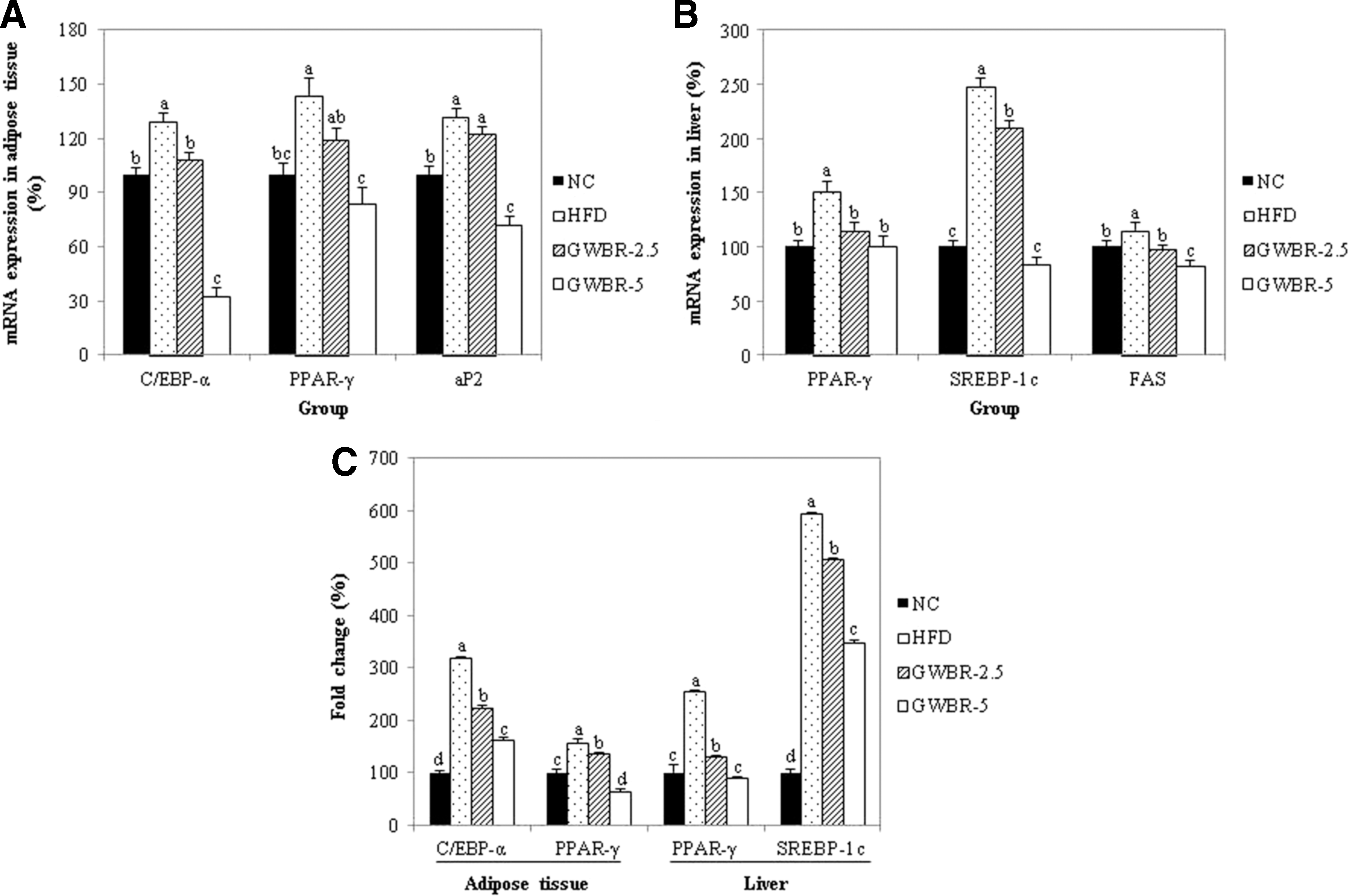
Effects of GWBR supplementation on expression of adipogenic genes in high-fat diet-induced obese mice. mRNA expression in adipose tissue
mRNA expression of lipases
To examine whether GWBR supplementation affects the mRNA expression of lipases involved in lipolysis, reverse transcription–polymerase chain reaction (RT-PCR) was performed. The mRNA expression of LPL increased in the HFD control group compared to the ND group (Fig. 4). Compared to the HFD control group, LPL levels in the GWBR-5 group were significantly lower by 40%. However, mRNA expression of HSL and ATGL was significantly lower in the HFD control group compared to the ND group. Supplementing with GWBR resulted in significantly higher HSL and ATGL compared to the HFD control group.
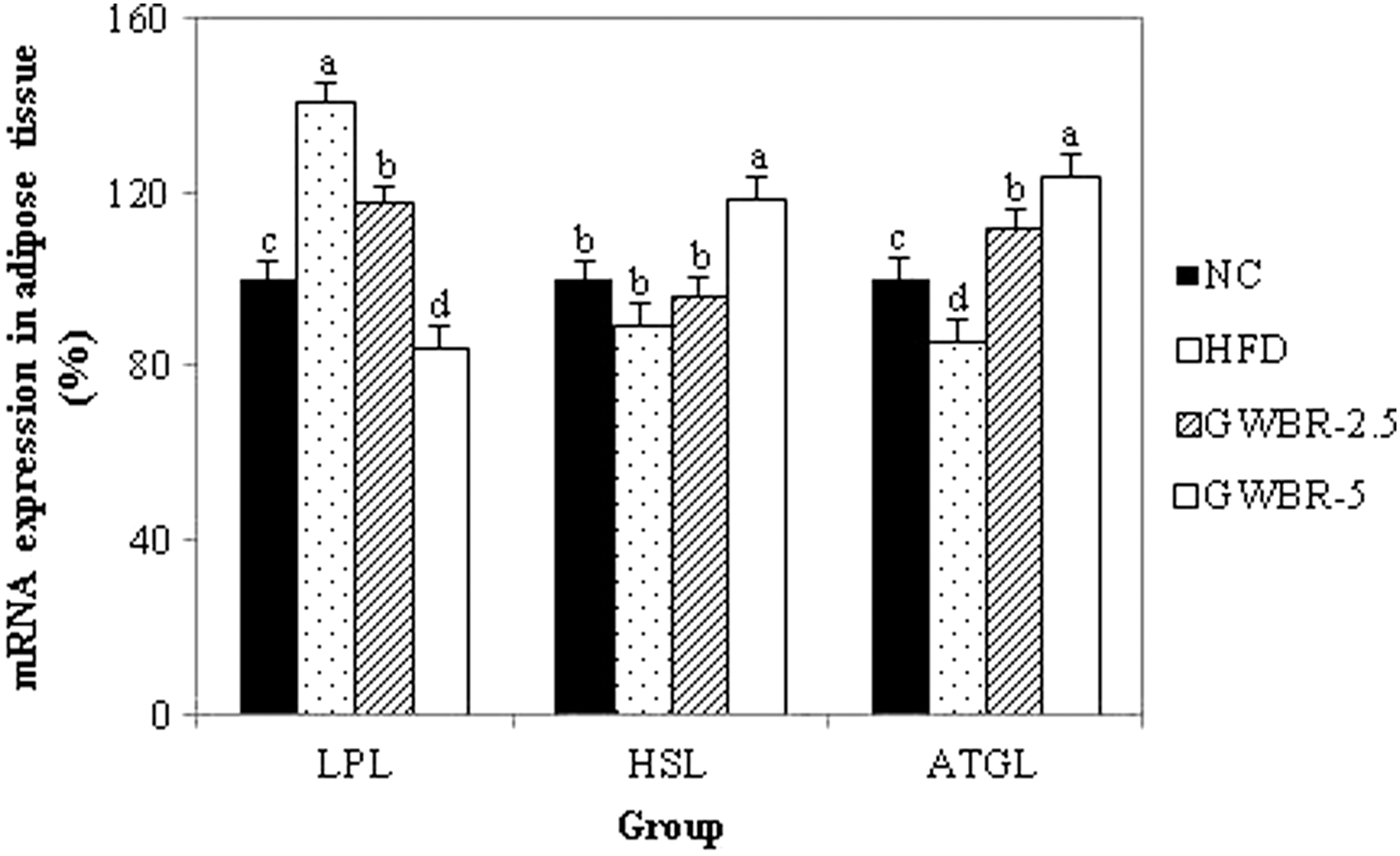
Effects of GWBR supplementation on mRNA expressions of lipases in high-fat diet-induced obese mice. Adipose tissue was prepared and used in RT-PCR, as described in the “Materials and Methods” section. Values were calculated as a percentage of lipase mRNA expression versus that of β-actin as an internal control. Results are presented as mean ± SD with at least 3 independent experiments, each performed in triplicate (n = 3). Different letters indicate a significant difference (P < .05), as determined by Tukey's multiple range test.
mRNA expression of genes involved in β-oxidation
Fatty acid β-oxidation is regulated in part by several molecular markers, such as CPT-1 and UCP. 12 Supplementation of GWBR resulted in higher CPT-1 and UCP2 mRNA expression in a dose-dependent manner compared to the HFD control group (Fig. 5).
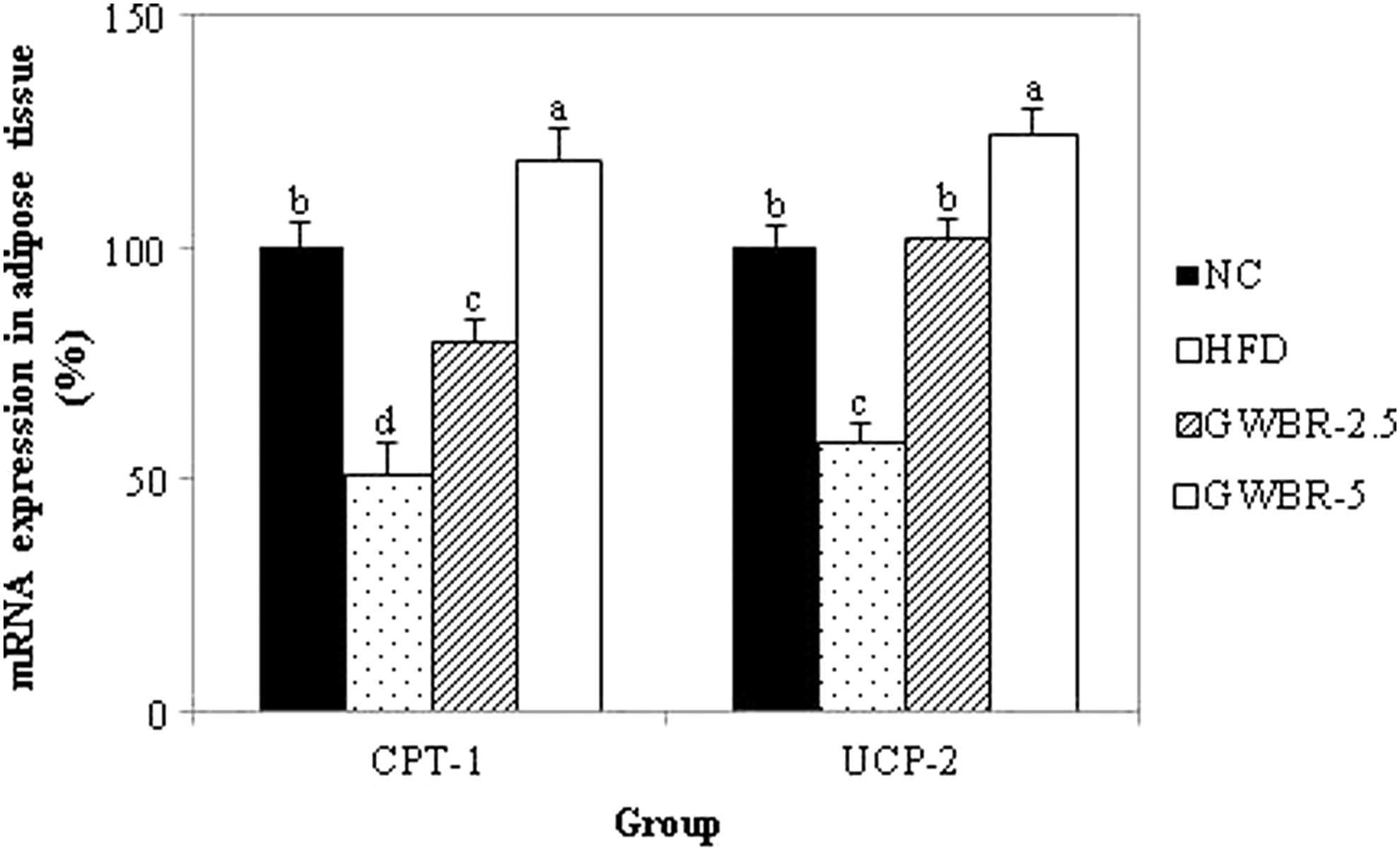
Effects of GWBR on the expression of β-oxidation-related genes in high-fat diet-induced obese mice. Adipose tissue was prepared and used in RT-PCR, as described in the “Materials and Methods” section. Values were calculated as a percentage of β-oxidation-related gene mRNA expression versus that of β-actin as an internal control. Results are presented as mean ± SD with at least 3 independent experiments, each performed in triplicate (n = 3). Different letters indicate a significant difference (P < .05), as determined by Tukey's multiple range test.
Discussion
Germination of grains is used to soften the texture, enhance the flavor, and increase the content of nutrients, such as γ-aminobutyric acid (GABA) and minerals, in brown rice. 13 Several studies have reported that rice extracts, such as red yeast rice, black rice, and germinated brown rice, exhibit antiobesity effects in HFD-induced obese animal models. 9,11,14 Therefore, in this study, we examined the antiobesity effect of GWBR on body weight gain, liver and adipose tissue weight, lipid profiles in serum and the liver, and mRNA expression of lipases, CPT-1, and UCP2 in HFD-induced obese mice. In addition to exploring the mechanism of GWBR effects on adipogenesis, we measured mRNA expression and protein level of adipogenic transcription factors and related genes.
In the present study, body weight gain and adipose tissue weights were significantly lower in the GWBR-supplemented group compared to the HFD control group. GWBR supplementation did not affect food intake; however, FER was significantly lower in the GWBR-supplemented group than in the HFD control group. In addition, lipid droplet formation in adipose tissue and liver was significantly less in the GWBR-5 group. Thus, the data demonstrate that the antiobesity effect of GWBR is associated with less fat accumulation in adipose tissue and liver as well as increased energy expenditure.
Serum TG and TC levels were significantly higher in the HFD control group compared to the ND group, whereas those levels were significantly lower in the GWBR-supplemented groups. Moreover, HDL-cholesterol in the GWBR-supplemented group was significantly higher than that in the HFD control group. These results indicate that supplementation of GWBR dose dependently suppressed lipid accumulation, resulting in improved lipid profiles in serum and liver.
In contrast, stored TGs in adipose tissue are hydrolyzed to FAs and glycerol by lipases, such as HSL, ATGL, and monoacylglycerol lipase, in a process known as lipolysis. 15,16 Over the years, a well-characterized model of lipolysis has been developed, in which activation of PKA leading to phosphorylation of a cellular lipase is known as HSL. 16 HSL and ATGL are expressed predominantly in adipose tissue. According to Zimmerman et al., ATGL is responsible for 75% of the cytosolic acylhydrolase activity in adipose tissue of HSL-deficient mice. Therefore, ATGL could participate together with HSL in adipose tissue lipolysis. 17 In this study, supplementation with GWBR significantly increased HSL and ATGL mRNA expressions compared to the HFD control group. Additionally, LPL is a key enzyme involved in TG accumulation. 18 Human and rodent studies reported increased adipose tissue LPL activity with obesity. Weight loss also selectively increases adipose tissue LPL activity as an attempt to maintain lipid stores. 19 LPL mRNA expression in the HFD control group was higher than that in the ND group, whereas GWBR supplementation significantly decreased LPL mRNA expression. These data suggest that the GWBR supplementation reduced adipose tissue weight and TG and TC concentrations, at least in part, by upregulating HSL and ATGL and downregulating LPL in adipose tissue.
Adipose tissue is a dynamic endocrine organ that not only maintains energy balance but also regulates lipid homeostasis mainly by lipogenesis and lipolysis. 20 PPAR-γ, C/EBP-α, and SREBP-1c are the critical transcriptional factors for adipogenesis. 21,22 These adipogenic transcriptional factors promote the expression of glucose-6-phosphate dehydrogenase (GPDH), FAS, acetyl-CoA carboxylase (ACC), and adipocyte-selective fatty acid-binding protein. 22 FAS, which plays a central role in de novo lipogenesis in mammals, is sensitive to nutritional status in lipogenic tissues, such as adipose tissues and livers. The regulation of FAS expression occurs primarily through changes in FAS gene transcription by SREBP-1, a master regulator of lipogenesis. 12 In the present study, adipogenic transcriptional factors and FAS mRNA expression were significantly downregulated by supplementation with GWBR. PPAR-γ and C/EBP-α, which are mainly found in adipose tissue, act as key transcription factors during adipogenesis and lipogenesis. 23 We found that GWBR significantly decreased the mRNA expression of PPAR-γ and C/EBP-α in adipose tissue and that GWBR significantly reduced the expression of these proteins compared to the HFD control group. Furthermore, the mRNA expression of PPAR-γ target gene aP2 was significantly reduced in adipose tissue by supplementing with GWBR. 24
Fatty acid oxidation is controlled in part by several genes, such as CPT-1, UCP, and acetyl-coenzyme A oxidase (ACO). 12 CPT-1 is a rate-limiting enzyme in β-oxidation, and CPT-1 mRNA expression is found to be inversely correlated with hepatic TG accumulation. 25 UCP2 is involved in the regulation of thermogenesis and plays an important role in lipid metabolism, energy balance, and the regulation of body weight. 26,27 GWBR significantly increased CPT-1 and UCP2 mRNA expressions in adipose tissue, suggesting that GWBR promotes energy metabolism. 28 Taken together, GWBR supplementation led to a decreased adipose tissue weight, which might be mediated by increased CPT-1, UCP2, HSL, and ATGL and decreased aP2, FAS, and LPL in adipose tissue.
In the present study, HFD control mice exhibited a significant upregulation of aP2, FAS, and LPL mRNA expressions along with downregulation of CPT-1, UCP2, HSL, and ATGL mRNA expressions, thus shifting the overall balance toward lipid accumulation. 29 However, GWBR supplementation increased HSL, ATGL, CPT-1, and UCP2 mRNA expressions, contributing to the stimulation of lipid catabolism in adipose tissue. 21 Thus, our results suggest that the decreased lipid accumulation in the GWBR-supplemented group was attributable to the inhibition of fatty acid synthesis and the promotion of fat catabolism. 7,21
In conclusion, in the present study, GWBR supplementation reduced body weight gain, liver and adipose tissue weights, serum TG and TC concentrations, expressions of adipogenic genes (aP2, FAS, and LPL), and lipases in HFD-induced obese mice. GWBR ameliorated HFD-induced obesity, probably by decreasing the expressions of regulatory genes associated with adipogenesis and increased lipolysis through upregulation of lipases (HSL and ATGL) and oxidation-related genes (CPT-1 and UCP2). These results suggest that GWBR may be a useful natural ingredient for controlling obesity.
Footnotes
Acknowledgment
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2010–0024307).
Author Disclosure Statement
No competing financial interests exist.