
Abstract
The aim of this present study was to investigate the effect of oridonin on visceral hyperalgesia and colonic serotonin availability in a rat model of trinitrobenzenesulfonic acid-induced postinflammatory irritable bowel syndrome (PI-IBS). Rats were randomly divided into five groups: normal control, PI-IBS model, PI-IBS+low-dose oridonin (5 mg/kg), PI-IBS+median-dose oridonin (10 mg/kg), and PI-IBS+high-dose oridonin (20 mg/kg). Rats in control and model groups were orally administered with water by gavage, whereas rats in oridonin-treated groups were orally administered with different dosages of oridonin, and drugs were given for 14 consecutive days. Compared with the control group, the pain threshold pressure was significantly reduced in PI-IBS rats. The colonic enterochromaffin (EC) cell number, serotonin content, and the protein expression of tryptophan hydroxylase (TPH) were markedly increased and the protein expression of serotonin reuptake transporter was significantly decreased in PI-IBS rats. The spleen index in PI-IBS rats was decreased, and the levels of tumor necrosis factor (TNF)-α, interferon (IFN)-γ, interleukin (IL)-4, and IL-13 in the colon of PI-IBS rats were also markedly decreased. Oridonin treatment dose dependently increased pain threshold pressure, and markedly decreased colon EC cell numbers, TPH expression, and serotonin content in PI-IBS rats. Oridonin treatment also significantly increased the spleen index as well as the levels of TNF-α, IFN-γ, IL-4, and IL-13 in the colon of PI-IBS rats. Results of this study demonstrate that the analgesic effect of oridonin in PI-IBS rats is associated with reduced colonic EC cell hyperplasia and 5-HT availability, the regulatory effect of oridonin on colonic cytokine production may be correlated with its effect on colonic EC cell number.
Introduction
P
It is well known that the enterochromaffin (EC) cell is the main cell type responsible for intestinal serotonin (5-HT) synthesis, storage, and release 5 ; tryptophan hydroxylase (TPH) is the rate-limiting enzyme of 5-HT synthesis. Upon stimulation, 5-HT is quickly released from EC cells and activates serotonergic receptors, and thus mediates gastrointestinal secretion, vasodilation, and perception of pain, 6 and the released 5-HT is finally removed by the serotonin reuptake transporter (SERT). Currently, colonic EC cell hyperplasia and decreased SERT expression were both considered to play vital roles in IBS symptom generation, such as visceral hypersensitivity. 7,8
The pathogenic mechanism of EC cell hyperplasia in IBS is still not clear. It was previously demonstrated that EC cell hyperplasia in IBS is commonly accompanied with intestinal infiltration of T lymphocytes. 9 It is reported that parasite infection-induced EC cell hyperplasia is T lymphocyte dependent, and EC cell number becomes increased in mice treated with helper T-cell type 2 (Th2) cells or deficient mice with impaired Th1 cytokines production, 10,11 suggesting that EC cell number may be influenced by intestinal Th1/Th2 balance. EC cells are the major sources of intestinal 5-HT availability, but up to now, few therapeutic strategies have been found to prevent or treat EC cell hyperplasia 12,13
Oridonin, one of the active components isolated from traditional Chinese herbal medicine Rabdosia rubescens (Donglingcao), has been reported to have multiple biological effects, such as anti-inflammatory, antibacterial, and anticancer effects. 14 A recent study showed that oridonin had multiple modulatory effects on the production of proinflammatory cytokines, such as nitric oxide, tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6, suggesting that oridonin may possess anti-inflammatory and immunoregulatory effects. 15 Our previous study also found that oridonin tablet had therapeutic effects on acetic acid-induced ulcerative colitis in mice, the underlying mechanism may have close correlation with its immunoregulatory effects. 16
Based on the above findings, we hypothesized that oridonin may attenuate visceral hyperalgesia in PI-IBS rats through reducing colonic EC cell hyperplasia and 5-HT bioavailability. In this study, 2, 4, 6-trinitrobenzenesulfonic acid (TNBS) was used to establish a PI-IBS model in rats. After oridonin treatment, pain threshold pressure, colonic EC cell numbers, 5-HT content, and the expression of TPH and SERT were evaluated. In addition, the underlying mechanisms of oridonin in PI-IBS were also explored.
Materials and Methods
Reagents
TNBS was purchased from Sigma-Aldrich Co. (St. Louis, MO). Chloral hydrate was purchased from Bangjing Scientific Supplies (Shanghai, China). ELISA kits for TNF-α, interferon (IFN)-γ, IL-4, and IL-13 assays were purchased from NEOBIOSCIENCE (Shenzhen, China). Oridonin was purchased from Hao-Xuan Pharmaceutical Co. Ltd. (Xian, China).
Animals and development of PI-IBS model
Sprague Dawley male rats (aged 6 weeks with body weight around 220 g) were obtained from the Laboratory Animal Center of Gansu University of TCM. Rats were maintained at 25°C under 12–12 h alternating light–dark cycle with free access to food and water. Rats were allowed to adapt the environment for at least 1 week before beginning the study. All the animal studies were carried out in accordance with the proposals of the Committee on the Ethics of Animal Experiments of Gansu University of TCM (Approval ID: 2014-031). The PI-IBS model was developed according to our previous study. 17 Briefly, rats were fasted for 12 h, and then deeply anesthetized with chloral hydrate (350 mg/kg, i.p.). A plastic catheter was inserted into the descending colon of rats at the depth of 8 cm from the anus, then TNBS (5 mg/rat, 0.8 mL in 50% ethanol) saline solution was slowly instilled into the colon. Rats in the control group were instilled with 0.8 mL saline. After TNBS administration, rats were kept in a head-down position to prevent TNBS leakage. At 4 weeks after TNBS administration, only the rats with acquired visceral hyperalgesia were selected as the PI-IBS rats.
Study design
Five groups were used in this study. Group 1 (n = 6) was set as the normal control, rats in this group were orally administered with drinking water. Group 2 (n = 6) was set as the model group, the rats in this group were orally treated with drinking water. Groups 3–5 (n = 6 per group) were set as oridonin-treated groups, rats were orally administered with oridonin at the dose of 5, 10, and 20 mg/kg, respectively. After 2 weeks of drug treatment, pain threshold pressure was measured by abdominal withdrawal reflex (AWR) test, and the rats were sacrificed for sample collection. A 6 cm long section of the proximal colon (1–2 cm from the cecum) was surgically removed and divided into three parts; the proximal was fixed in 4% paraformaldehyde for immunohistochemistry; the middle part was used for measuring 5-HT content and Western blot assay; and the distal colon was used for ELISA assay. Spleens and thymuses were also removed and weighed to determine thymus and spleen indices, which were calculated as the ratio of thymus or spleen weight (mg) to the body weight (g).
Abdominal withdrawal reflex
AWR test was performed as previously reported. 18 Briefly, rats were fasted for 12 h, lightly anesthetized with ether, then a 6 cm-long flexible latex balloon was inserted into the descending colon and the end of the balloon was fastened to the anal verge of rats. The tube of the balloon was connected through a Y-connector to a sphygmomanometer. After recovery for about 1 h, colorectal distension was applied to rats until a visible contraction of the abdominal wall was observed. Pain threshold pressure was defined as the intensity of colorectal distension that elicited an observable AWR, that is, a sudden and persistent abdominal muscle contraction with abdomen lift off the platform. The AWR test was repeated for five times in each rat with intervals of at least 5 min for recovery. Data of each rat were averaged, and the results were compared across groups.
EC cell counting
Tissue sections (5 μm thick) were deparaffinized and rehydrated for immunohistochemistry. Briefly, sections were incubated with primary antibody (anti-serotonin, 1:4000; Sigma-Aldrich) overnight at 4°C, followed by the labeled streptavidin biotin (the LSAB + System HRP Kit; Dako). The primary antibody was omitted on one section of each reaction as a negative control. All the sections were counterstained with Hematoxylin for nuclei labeling. Five random fields at 200× magnifications per section were counted by a researcher blinded to the treatment; the number of EC cells per mm2 of the mucosa was quantified using ImageJ software (National Institutes of Health, Bethesda, MD).
5-HT content determination
The 5-HT content in colon tissue was determined by an Agilent1260 Infinity HPLC (Agilent Technologies, Santa Clara, CA) coupled to an Agilent 6460 triple quadrupole mass spectrometer (Agilent Technologies) equipped with the electrospray ionization interface, the procedure was conducted as previously reported. 19 Briefly, colon tissue was homogenized in 15% iced trichloroacetic acid, then centrifuged at 10,000 g for 10 min. The supernatant was added to acetonitrile (containing 1 μg/mL methimazole) and the mixtures were vortexed for 5 min. For derivatization, borate buffer (sodium tetraborate, 100 mM in water) and benzoyl chloride (2% in acetonitrile, v/v) were added to the mixtures, and then vortexed for 5 min. After centrifugation at 10,000 g for 10 min, a 20 μL aliquot was transferred to the vial and a 5 μL sample was injected for liquid chromatography–tandem mass spectrometry (Agilent Technologies) analysis.
Western blot analysis
Colon tissues were homogenized and sonicated on ice for protein extraction. The protein content was determined, and then samples were denatured at 100°C for 5 min. Proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membrane (Bio-Rad Laboratories, Inc., Hercules, CA). The membrane was blocked with 5% nonfat milk, and then incubated with primary antibody (anti-TPH, 1:1000; Santa Cruz Biotechnology, Santa Cruz, CA; anti-SERT, 1:1000; Merck Millipore Corporation, Darmstadt, Germany) overnight at 4°C, then incubated with the appropriate secondary antibodies. The immunoreaction was detected using the Amersham ECL Western Blotting Detection Kit (GE Healthcare Life Sciences, Amersham, United Kingdom), and the optical density of each band was semi-quantified by software (ImageJ). The detected protein bands were normalized to actin or GAPDH levels.
ELISA assay
The colon samples were lysed in iced Tris-EDTA solution (10 mM Tris-HCl, 1 mM EDTA, pH7.4) containing 0.5% Triton X-100 and protease inhibitors (Cocktail Tablets, Roche, Mannheim, Germany). The samples were homogenized and sonicated on ice. After centrifuging at 10,000 g for 10 min at 4°C, the supernatant was collected and stored at −80°C until detection. Colonic cytokine levels of TNF-α, IFN-γ, IL-4, and IL-13 were determined using the ELISA Kits according to the manufacturer's protocol and instruction. All the results were presented as pg/mg total protein.
Statistical analysis
Data are presented as mean ± standard error of the mean. Statistical analysis was conducted using SPSS 12.0 Software (SPSS, Inc., Chicago, IL). Tests of homogeneity of variances were carried out first. For the normal distribution data, differences between two groups were analyzed by Student's t-test, when multiple groups were compared, data were analyzed using one-way ANOVA, followed by Student–Newman–Keuls test. For the abnormal distribution data, a nonparametric test (Mann–Whitney U-test) was used for two-group comparison. Differences were considered significant when P < .05.
Results
Effect of oridonin on pain threshold pressure in PI-IBS rats
As shown in Figure 1A, pain threshold pressures in PI-IBS rats were significantly decreased when compared with that of the control (P < .05). Compared with the PI-IBS rats, the pain threshold pressures in oridonin-treated rats were significantly and dose-dependently elevated with an EC50 of 8.2 mg (P < .05).
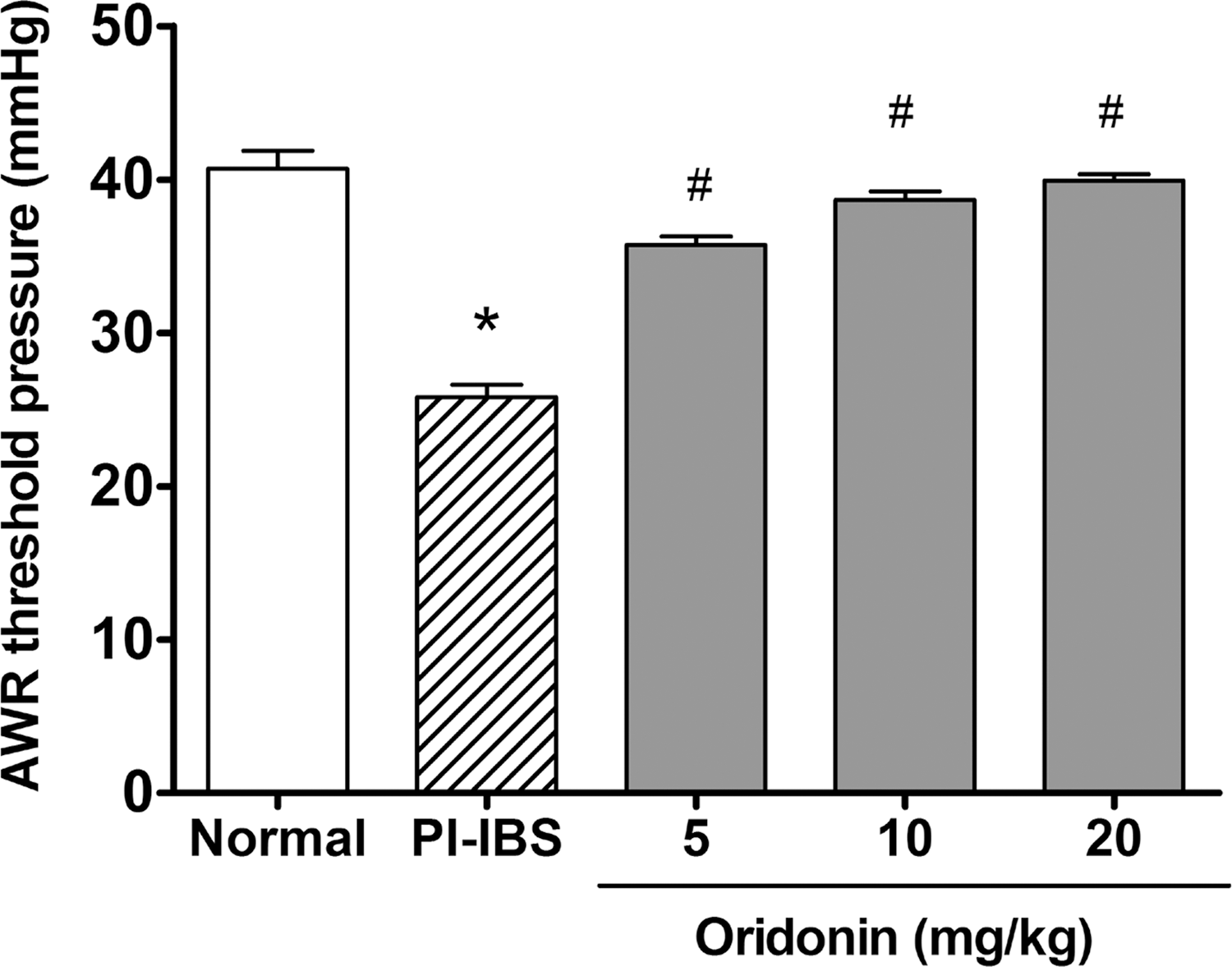
Effects of oridonin on visceral hypersensitivity of TNBS-induced PI-IBS rats. Data are presented as mean ± SEM, n = 6 per group. *P < .05 versus normal rats, # P < .05 versus TNBS-induced PI-IBS rats. PI-IBS, postinflammatory irritable bowel syndrome. SEM, standard error of the mean; TNBS, 2, 4, 6-trinitrobenzenesulfonic acid.
Effects of oridonin on colonic EC cell number and 5-HT content in PI-IBS rats
As shown in Figure 2, the colon EC cell density and 5-HT content were both significantly increased (−47% increase in EC cell density, −69% increase in 5-HT content) in PI-IBS rats when compared with that of the control (P < .05). Compared with the PI-IBS rats, oridonin treatment significantly and dose-dependently decreased the colon EC cell density (16–30%) and 5-HT content (15–38%) in PI-IBS rats.
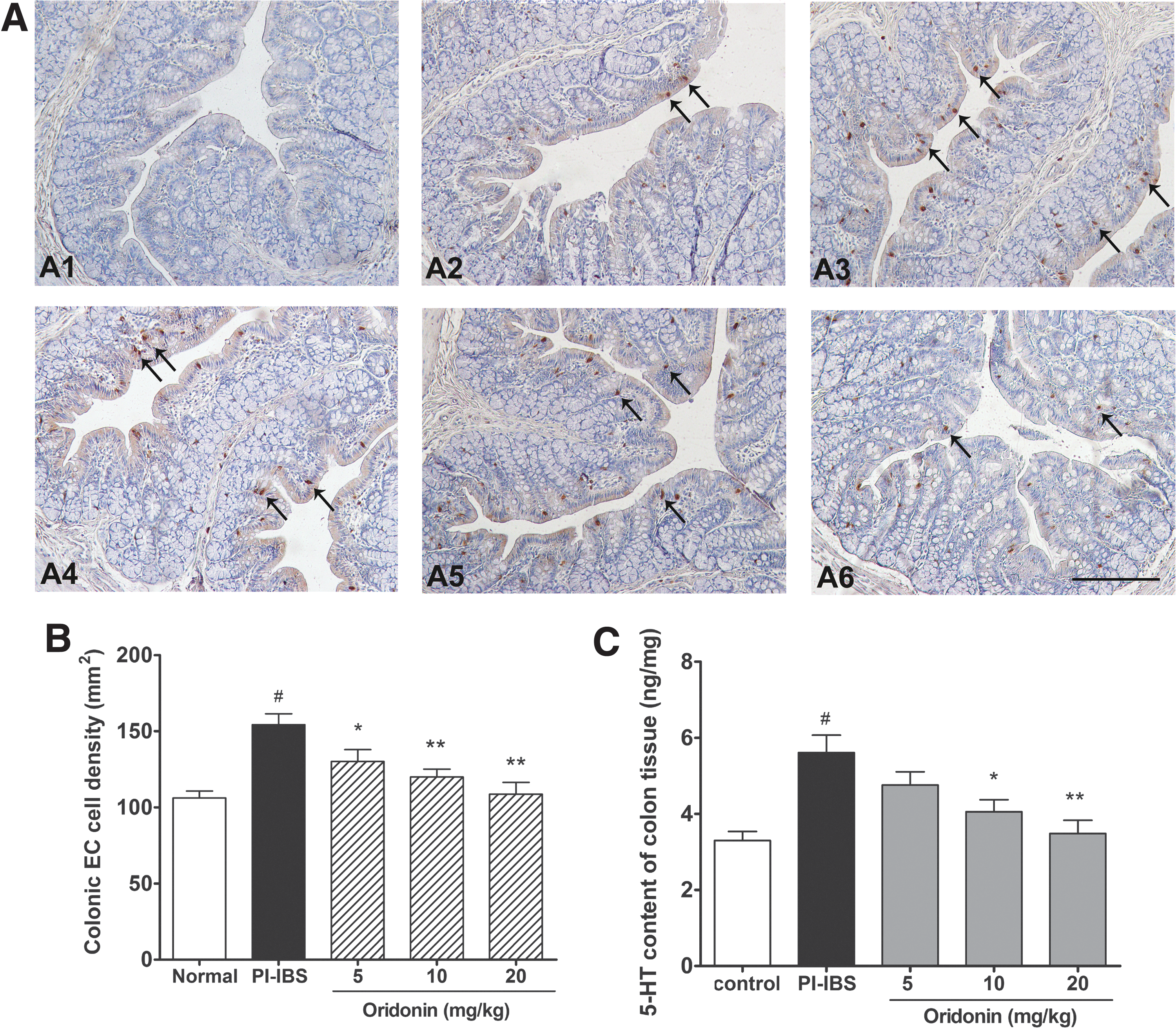
Effects of oridonin on colonic EC cell density and 5-HT content in TNBS-induced PI-IBS rats.
Effect of oridonin on protein expression of TPH and SERT in PI-IBS rats
As shown in Figure 3A, colonic protein expression of TPH in PI-IBS rats was significantly greater when compared wih that of the normal rats (P < .05), whereas high-dose oridonin treatment significantly decreased the protein expression of TPH in PI-IBS rats. These findings are consistent with the alteration of EC cell number in PI-IBS rats. As shown in Figure 3B, the protein level of SERT in PI-IBS rats was markedly decreased when compared with that of the normal rats (P < .05), but high-dose oridonin treatment did not show any effect on the protein expression of SERT in PI-IBS rats (P < .05).
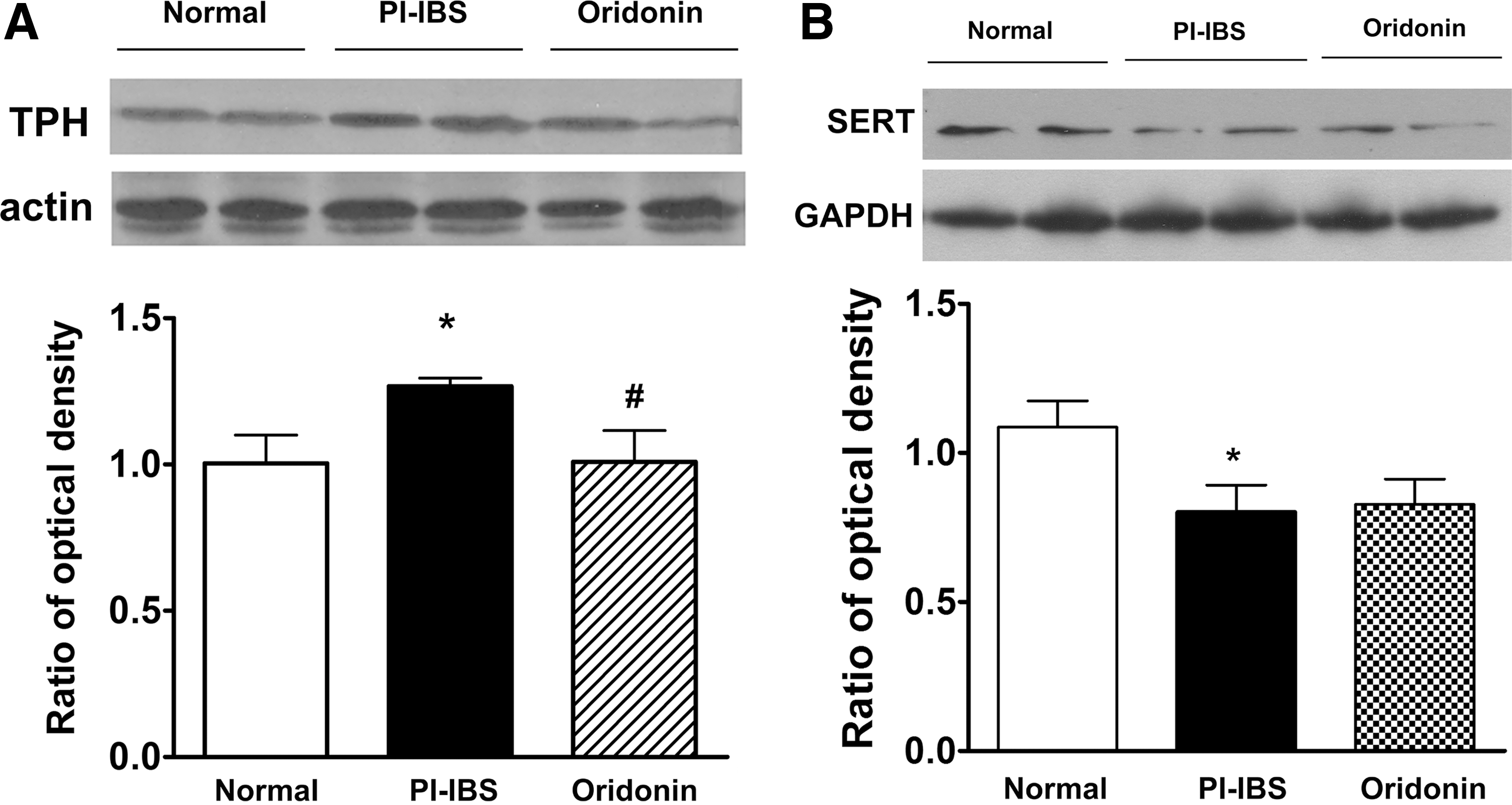
Effects of oridonin on protein expression of TPH and SERT in the colon tissue of PI-IBS rats. Representative western immunoblots of TPH and SERT as well as the statistical analysis of protein level are shown in
Effect of oridonin on spleen and thymus indices in PI-IBS rats
As shown in Table 1, the thymus indices in PI-IBS rats were slightly decreased, but no differences were found between PI-IBS rats and normal rats. High dose of oridonin treatment slightly, but not significantly, elevated the thymus indices in PI-IBS rats. Moreover, the spleen indices in PI-IBS rats were significantly decreased when compared with that of the normal rats (P < .05), whereas oridonin treatment dose-dependently and markedly elevated the spleen indices in PI-IBS rats.
Data are presented as mean ± SD (n = 6).
P < .05 versus normal rats.
P < .05 versus PI-IBS rats.
PI-IBS, postinflammatory irritable bowel syndrome; SD, standard deviation.
Effect of oridonin on colonic cytokine production in PI-IBS rats
As shown in Figure 4A, compared with the normal rats, the levels of TNF-α, IFN-γ, IL-4, and IL-13 in the colon of PI-IBS rats were all significantly decreased (P < .05). High dose of oridonin treatment significantly elevated the levels of TNF-α, IFN-γ, IL-4, and IL-13 in PI-IBS rats (P < .05). As shown in Figure 4 (B), the detected Th1- and Th2-related cytokines were all decreased significantly in the colon of PI-IBS rats. The levels of Th1-related cytokines, that is, TNF-α and IFN-γ, were decreased more markedly (49% decrease in TNF-α, 58% decrease in IFN-γ) than Th2-related cytokines (19% decrease in IL-4, 14% decrease in IL-13), and oridonin treatment dose-dependently and significantly reversed the decreases in colon cytokine levels in PI-IBS rats.
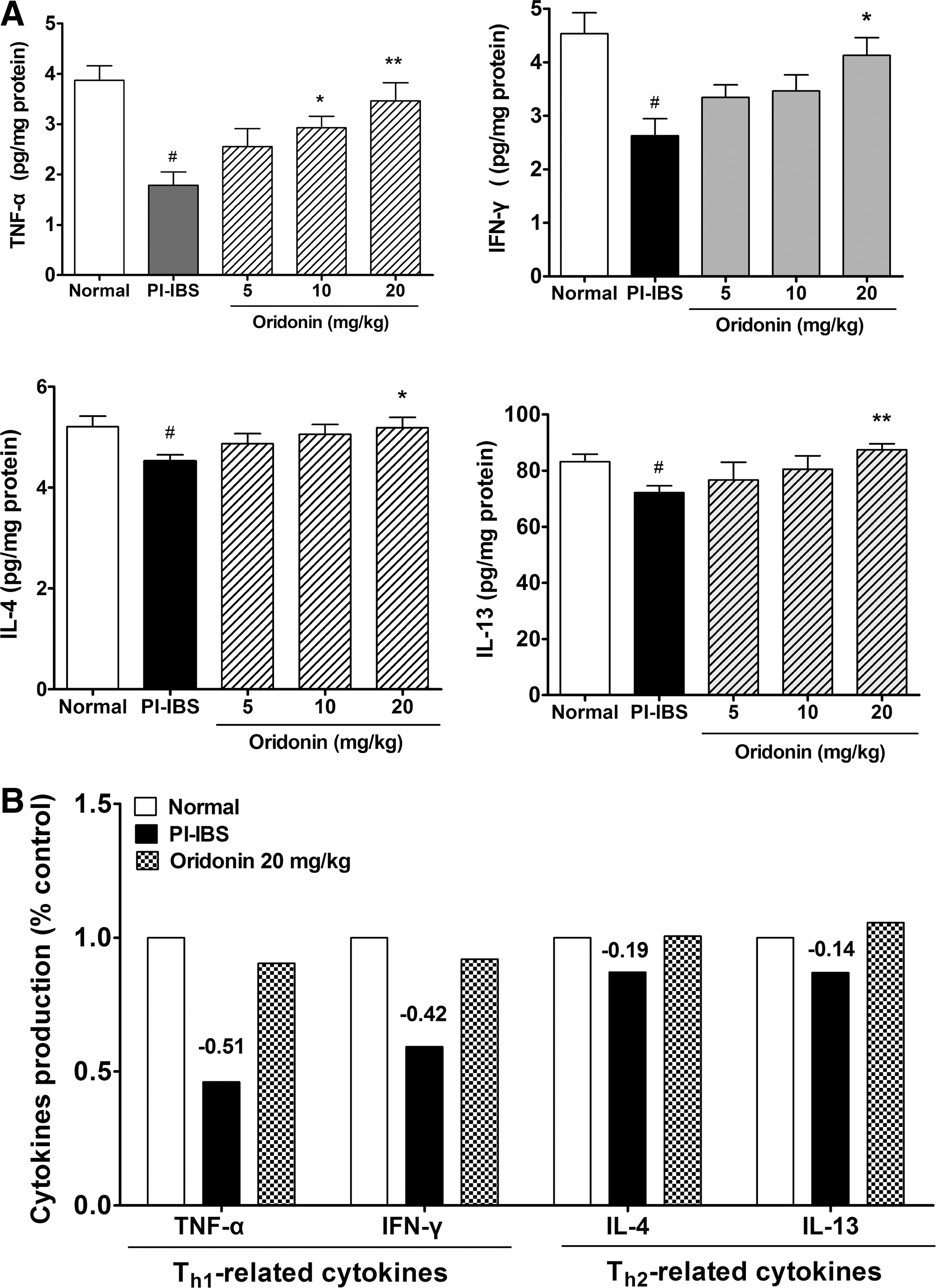
Effects of oridonin on cytokine production in the colon of PI-IBS rats. Statistical graphs about the levels of TNF-α, IFN-γ, IL-4, and IL-13 are shown in
Discussion
PI-IBS is a subset of IBS. PI-IBS patients present with IBS symptoms that occur after an acute intestinal infection or inflammation. The major symptoms of PI-IBS include loose stool, visceral hypersensitivity, and accelerated colon transit. This present study investigated the effect of oridonin, the main active component of Rabdosia rubescens, on visceral hyperalgesia in TNBS-induced PI-IBS rats, the analgesic effect and the underlying mechanism of oridonin were further investigated based on colonic EC cell counting and 5-HT bioavailability study. Our results found that oridonin treatment can attenuate visceral hypersensitivity, decrease colonic EC cell number, and 5-HT availability. Moreover, oridonin treatment can regulate colonic cytokine production, and thus rebalance the Th1/Th2-related cytokine ratio in the colon of PI-IBS rats.
5-HT is a neurotransmitter and a paracrine messenger; it plays an important role in the perception of nociceptive stimuli in the gastrointestinal tract. It has been reported that the increase of colonic EC cell number and 5-HT content are involved in the development of visceral pain and motility dysfunction of IBS. 9 Results from our study are consistent with previous findings; we found that EC cell number and 5-HT content in PI-IBS rats were markedly increased in the colon tissue. 20,21 We also found that oridonin treatment dose-dependently reduced visceral hypersensitivity in PI-IBS rats, and this effect was concomitant with the lower colonic EC cell number and 5-HT content. Given the vital role of 5-HT in the symptom generation of IBS, it is possible that the decreased colonic EC cell number and 5-HT content in oridonin-treated rats may be responsible for the attenuated visceral hyperalgesia of PI-IBS rats.
Moreover, in the gastrointestinal tract, 5-HT content comes from synthesized 5-HT in EC cells and the released 5-HT that has not been taken up by SERT, 22 we further investigated the effect of oridonin on protein expression of TPH and SERT in PI-IBS rats. We found that colonic TPH expression in PI-IBS rats was increased, whereas SERT expression was decreased; this result is in agreement with previous reports, which revealed that SERT expression is reduced in IBS patients and animal models. 23,24 We also found that high dose (20 mg/kg) of oridonin treatment reduced the protein expression of TPH in colon tissue of PI-IBS rats, but did not show any effect on SERT expression. These findings indicated that oridonin may reduce colonic 5-HT bioavailability in PI-IBS rats by decreasing colonic EC cell number and TPH expression.
To date, the mechanism concerning EC cell hyperplasia in PI-IBS is still not clear, but it is reported to be correlated with mucosal T lymphocytes, especially the Th1 or Th2 cytokine predominance. 10 Moreover, EC cell hyperplasia was observed in deficient mice with impaired Th1 cytokine production. 11 Our results revealed that the level of TNF-α, IL-1β, IL-4, and IL-13 in the colon of PI-IBS rats were all significantly decreased, but Th1-related cytokine levels were decreased more obviously than Th2-related cytokine levels. It is still not clear why colonic cytokine level was reduced in PI-IBS rats, but this finding is consistent well with a previous report showing reduced intestinal cytokine production in PI-IBS patients. 25 Our result also showed that the spleen indices in PI-IBS rats were significantly decreased; this finding is in agreement with the reduced colonic cytokine levels in PI-IBS rats, and both these data indicate that intestinal immune response may be suppressed in PI-IBS rats. Oridonin has been found to modulate the production of proinflammatory cytokines, such as TNF-α, IL-1β, and IL-6, 15 suggesting that oridonin possess immunoregulatory effects. In this present study, we found that oridonin treatment significantly upregulated the colonic cytokine levels of TNF-α, IL-1β, IL-4, and IL-13 in PI-IBS rats, and thus rebalanced colonic Th1/Th2-related cytokine levels.
Oridonin treatment also significantly increased the spleen index in PI-IBS rats, these data indicating that oridonin indeed had an immunoregulatory effect and oridonin treatment can upregulate the suppressed intestinal immune response in PI-IBS rats. Considering the possible role of T lymphocytes in regulating EC cell number, we hypothesize that the rebalanced colonic Th1/Th2-related cytokine production in oridonin-treated rats may be responsible for the reduced colonic EC cell number in PI-IBS rats. Up to now, the mechanism of oridonin in immune regulation has not been very clear; however, a previous study suggests that it is associated with inhibited proliferation of CD4+ T cells and upregulating apoptosis of lymphocytes through nuclear translocation of transcription factor nuclear factor-kappa B. 26 Even though, oridonin treatment modulated colonic Th1/Th2-related cytokine production in this study, it is still not clear whether oridonin treatment can regulate the quantity of Th1 or Th2 lymphocytes in the colon of PI-IBS rats, and further studies are still needed to explore the effect of oridonin on the subgroups of T lymphocytes.
It is well known that oridonin is the main component of Rabdosia rubescens. According to the reported extraction rate, 27 the dose of oridonin tested in this study (5–20 mg/kg in rats), when converted into the dose of raw herbal material, is 0.7–2.8 g Rabdosia rubescens per kg body weight of rats. The dosage of oridonin selected in this study is based on our previous result, in which an oridonin tablet (4.8 g/kg) showed a therapeutic effect in mice with ulcerative colitis. 16 One limitation of this study is that we only observed the effects of three doses (5, 10, and 20 mg/kg) of oridonin in PI-IBS rats, and the effect of higher doses (exceeding 20 mg/kg) of oridonin has not been tested. Nevertheless, this study supports the hypothesis that oridonin is a potential therapeutic drug that can be developed for the treatment of PI-IBS.
In conclusion, the analgesic effect of oridonin in TNBS-induced PI-IBS rats may be mediated through reducing colonic EC cell hyperplasia and 5-HT bioavailability, the regulatory effect of oridonin on colonic Th1/Th2-related cytokine production may be responsible for the alterations induced by oridonin in PI-IBS rats. Results from this present study, for the first time, provide experimental evidence about the therapeutic potential of oridonin in PI-IBS.
Footnotes
Acknowledgments
This work was supported by the National Science Foundation of China (81341145) and the Fundamental Research Funds for the Central Universities (lzujbky-2014-223).
Author Disclosure Statement
No competing financial interests exist.