
Abstract
Adzuki beans have long been cultivated as a food and folk medicine in East Asia. In this study, we investigated the effect of black adzuki bean (BAB) extract on pancreatic cells and determined their mechanism of action in impaired glucose tolerance in an animal model of type 2 diabetes. In addition, we performed functional gene annotation analysis to identify genes related to the regulation of glucose metabolism and insulin response. Treatment of pancreatic β cells with BAB extract (0.2 mg/mL) led to tolerance of the high glucose-induced glucotoxicity, resulting in a similar viability as cells maintained in normal glucose media. In addition, dietary supplementation with BAB extract significantly (P < .05) improved hyperglycemia and homeostasis model assessment of insulin resistance index (HOMA-IR) in high-fat diet-induced glucose-intolerant obese C57BL/6J mice. Our results suggest that BAB extract ameliorates hyperglycemia and glucose intolerance, and lowers HOMA-IR by regulating insulin secretion and response, and by maintaining the integrity of pancreatic β cells exposed to hyperglycemic conditions.
Introduction
D
T2D and obesity are closely correlated. 5 Diet-induced obesity can cause T2D, and peripheral insulin resistance is common during obesity. 6 It is well known that when C57BL/6J mice are fed a high-fat diet (HD) ad libitum, these mice develop obesity, hyperinsulinemia, and hyperglycemia. 7 This is a consequence of increased adipose tissue mass leading to increased secretion of adipokines, cytokines, and proteins that deregulates glucose homeostasis and insulin sensitivity. 8 Therefore, relevant rodent models and pancreatic β cells represent good starting points for exploring the mechanisms altered in the development of T2D. 4
Recently, natural plant products and naturally occurring polyphenolic compounds have become a focal point of research interests because of their potential for treating diseases, with considerable attention devoted to managing diabetes. 9 Adzuki beans have long been cultivated throughout East Asia, where they have been the subject of extensive investigation because of their biological activities. In this study, we focused on the black adzuki bean (BAB), which has remarkable levels of flavonoids, anthocyanins, and proanthocyanidins compared with the other adzuki beans. 10 Consistent with previous studies of adzuki beans, there were several representative flavonols including kaempferol (Kfr). 11,12 Kfr promotes insulin sensitivity and preserves pancreatic cells, and reverses hyperglycemia and glucose intolerance. 6,9,13 Furthermore, it suppresses lipid accumulation in 3T3-L1 cells and zebrafish, also hypercalorie diet supplemented with Kfr reduces body weight in mice. 14,15
In the past, adzuki beans have been recommended as suitable foods for diabetic patients. 16,17 In addition, although the cascade of events underlying the improvement in insulin response related to adzuki beans remains uncertain, several studies have suggested that adzuki beans have antiobesity and antidiabetes effects. 18 –21
In this study, we investigated the effect of BAB extract on pancreatic cells and investigated their mechanism of action in an animal study. In addition, we performed functional gene annotation analysis to identify genes associated with the regulation of glucose metabolism and insulin response.
Materials and Methods
Preparation of test materials
BABs were obtained from the Rural Development Administration, Republic of Korea. Bean extracts were prepared by adding 80% ethanol to ground BABs, which were then concentrated by evaporating the ethanol using a rotary vacuum evaporator (Eyela, Tokyo, Japan) followed by freeze drying as described in a previous study. 18
Quality control of the extract was performed using high-performance liquid chromatography. Kfr was isolated from the ethyl acetate fraction of BAB ethanol extract, its chemical structure was elucidated by the direct comparison of NMR data to results reported in the literature (data not shown). 22
Cell culture
RINm5F rat insulinoma pancreatic β cells were obtained from ATCC. Cells were cultured in RPMI-1640 medium containing 11.1 mM
Cell viability assay
The effect of BAB extract on cell viability was assessed using EZ-Cytox reagent (Daeil Lab Service, Seoul, Korea). RINm5F cells were seeded in 96-well plates and allowed to adhere overnight. Then, cells were cultured in RPMI-1640 medium with normal glucose levels (11 mM) or high glucose levels (25 mM) in the presence or absence of various concentrations of BAB extract for 96 h in 96-well plates. After the experimental time period, the culture medium was replaced with medium containing EZ-Cytox solution (0.01 mL/well) and incubated at 37°C for an additional 2 h. Absorbance was measured at 450 nm using a microplate reader (MRX II; Dynex Technologies, Chantilly, VA). Cell viability (%) was calculated using the following equation:
Bromodeoxyuridine incorporation assay
The incorporation of bromodeoxyuridine (BrdU) in RINm5F cells was quantified using a colorimetric cell proliferation ELISA BrdU assay (Roche, Mannheim, Germany). In brief, cells were treated as already described for the viability assay, after which time a BrdU solution (10 μM) was added to the culture medium for 2 h. After BrdU labeling, the culture medium was removed and the extent of BrdU incorporation into the DNA was measured.
Quantification of insulin secretion
Cells were preincubated in media containing normal or high glucose levels for 96 h, after which time the cells were stimulated by replacing the medium with normal or high glucose levels with or without BAB extract. After stimulation for 2 h, levels of insulin in the culture medium were quantified using a rat high-range insulin ELISA kit (Alpco, Salem, MA).
Animals and study design
Four-week-old male C57BL/6J mice (Charles River Laboratories, Tokyo, Japan) were housed in plastic cages under controlled conditions. After 1 week, animals were randomly divided into 4 groups of 10 animals each and fed for 12 weeks with either control diets (CD) containing 10% kcal from fat (D12450B), HD containing 60% kcal from fat (D12492), a HD mixed with a 1% freeze-dried ethanolic extract of BAB (BAB), or a HD mixed with 0.08% Kfr. 23,24 Kfr was purchased from Aktin Chemicals (Chengdu, China). The diet compositions are listed in Table 1 and were based on D12450B and D12492 (Research Diets, Inc., New Brunswick, NJ). All animal procedures were conducted in accordance with the Guide for the Care and Use of Laboratory Animals after obtaining approval from the Institutional Animal Care and Use Committees of the Chonbuk National University (CBU 2014-00029).
CD, control diets containing 10% kcal fat (D12450B); HD, high-fat diets control containing 60% kcal fat (D12492); BAB, high-fat diets (D12492) mixed with 1% freeze-dried ethanolic extract of black adzuki bean; Kfr, high-fat diets (D12492) mixed with 0.08% kaempferol.
Blood glucose and oral glucose tolerance test
Fasting blood glucose levels were measured once every 2 weeks using a glucometer (Accu-Check; Roche). An oral glucose tolerance test (OGTT) was conducted at the end of the experiment using a 25% solution of glucose in sterile saline at a dose of 1.0 g/kg. Glucose was administered by orogastric gavage after a 12 h fasting period. Glucose concentration was measured before glucose administration (0 min) as well as at 30, 60, 120, and 180 min after administration. Results are shown as the area under the curve (AUC) of the glycemic index. The AUC was calculated using the trapezoidal rule.
Serum insulin and homeostasis model assessment of insulin resistance index
Serum insulin concentrations were measured using a mouse ultrasensitive insulin ELISA kit (Alpco). The homeostasis model assessment of insulin resistance index (HOMA-IR) was calculated as fasting glucose (mmol/L) multiplied by the fasting insulin level (uIU/mL) divided by 22.5.
Gene expression profiling
Total RNA of liver tissue was isolated using Trizol reagent (Life Technologies, Inc., Carlsbad, CA). For target labeling and microarray hybridization, the synthesis of target cRNA probes and hybridization were performed using Agilent's Low RNA Input Linear Amplification kit (Agilent Technology, Santa Clara, CA). After checking labeling efficiency, the fragmented cRNA was then resuspended with 2× hybridization buffer and directly pipetted onto an assembled Agilent Mouse whole genome 44K microarray.
All data normalization and selection of fold changes in gene expression levels were performed. The differences in significant gene expression were a value of P < .05 and, at least, a two-fold change in gene expression. Genes exhibiting differences in expression levels were classified using the DAVID Bioinformatics Resources with functional categories (
Statistical analyses
All data are presented as the mean ± standard deviation. Statistically significant differences were determined using one-way ANOVA. After ANOVA, differences between means were assessed using Duncan's multiple range test. Values of P < .05 were considered significant.
Results
BAB improves cell viability, proliferation, and glucose-stimulated insulin secretion of pancreatic β cells during incubation in high glucose
We first investigated the viability of pancreatic cells treated with different concentrations of BAB extract ranging from 0.0125 to 0.2 mg/mL in normal glucose medium (11 mM) after an incubation period of 96 h. The results showed that treating cells with 0.05 or 0.1 mg/mL BAB extract significantly increased cell proliferation, and no cytotoxic effect of BAB extract was observed (Fig. 1A). Cells were then exposed to medium containing 25 mM glucose for 96 h to mimic hyperglycemic conditions (Fig. 1B). Exposure of pancreatic β cells to 25 mM glucose significantly reduced cell viability by ∼20%; however, in the presence of BAB extract, the cells were able to tolerate high glucose-induced glucotoxicity, exhibiting a viability similar to cells maintained in normal glucose medium.

BAB improves pancreatic β-cell viability and proliferation. RINm5F cells were incubated in RPMI1640 medium containing glucose at normal levels (NG; 11 mM) or high levels (HG; 25 mM) with or without various concentrations of (BAB; 0.0125, 0.025, 0.05, 0.1, and 0.2 mg/mL) for 96 h. Cell viability and proliferation were assessed by EZ-cytox assay
Next, to determine the effects of BAB extract on cell proliferation, we measured the rate of BrdU incorporation in the presence or absence of BAB extract and under hyperglycemic conditions. Consistent with the cell viability results, the hyperglycemic condition inhibited cell proliferation. However, the rate of BrdU incorporation into cells treated with BAB extract increased in a dose-dependent manner. Ultimately, the rate of proliferation in cells under high glucose conditions treated with BAB extract was improved similar to that of cells incubated with normal levels of glucose (Fig. 1D).
We also measured the level of insulin secretion by pancreatic β cells in the presence or absence of BAB extract. As shown in Figure 2, the insulin secretion of cells cultured in 25 mM glucose was significantly increased compared with the cells cultured in 11 mM glucose. Furthermore, the addition of BAB extract increased the levels of insulin secretion.

BAB improves high glucose impaired insulin secretion. Data are expressed as the mean ± SD. Different letters indicate significant differences among groups according to ANOVA with Duncan's multiple range test (P < .05).
Effect of BAB extract on HD-induced body weight gain
The initial body weights of animals in each of the groups were within the same range (Fig. 3A), and all animals gained weight as the study progressed, as expected. Body weight gain (Fig. 3B) was highest in the HD control group compared with that in the control diet-fed group. Interestingly, supplementation of BAB extract along with HD significantly averted the increase in body weight compared with the HD control group, which was similar to animals supplemented with Kfr.
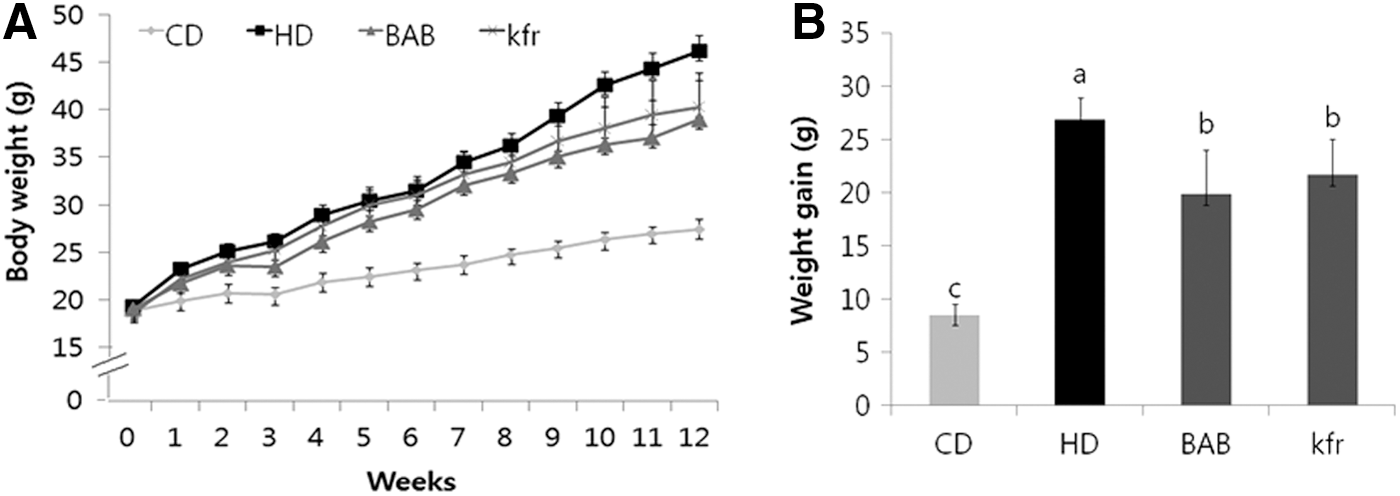
Effects of BAB on
Effect of BAB extract on fasting blood glucose, serum insulin, and HOMA-IR index
All of the mice fed a HD showed significantly increased fasting blood glucose levels compared with the CD group (Fig. 4A). Thus, it was interesting to observe that animals fed a HD along with BAB extract had a significantly lower fasting blood glucose level throughout the experimental period than animals in the HD group. At the end of the experimental period, we observed that animals in the HD group had a significantly higher level of fasting serum glucose than the animals in the control diet-fed CD group (Fig. 4C). Consistent with the said results, supplementing the HD with BAB extract prevented the increase in fasting serum glucose levels compared with the HD group.
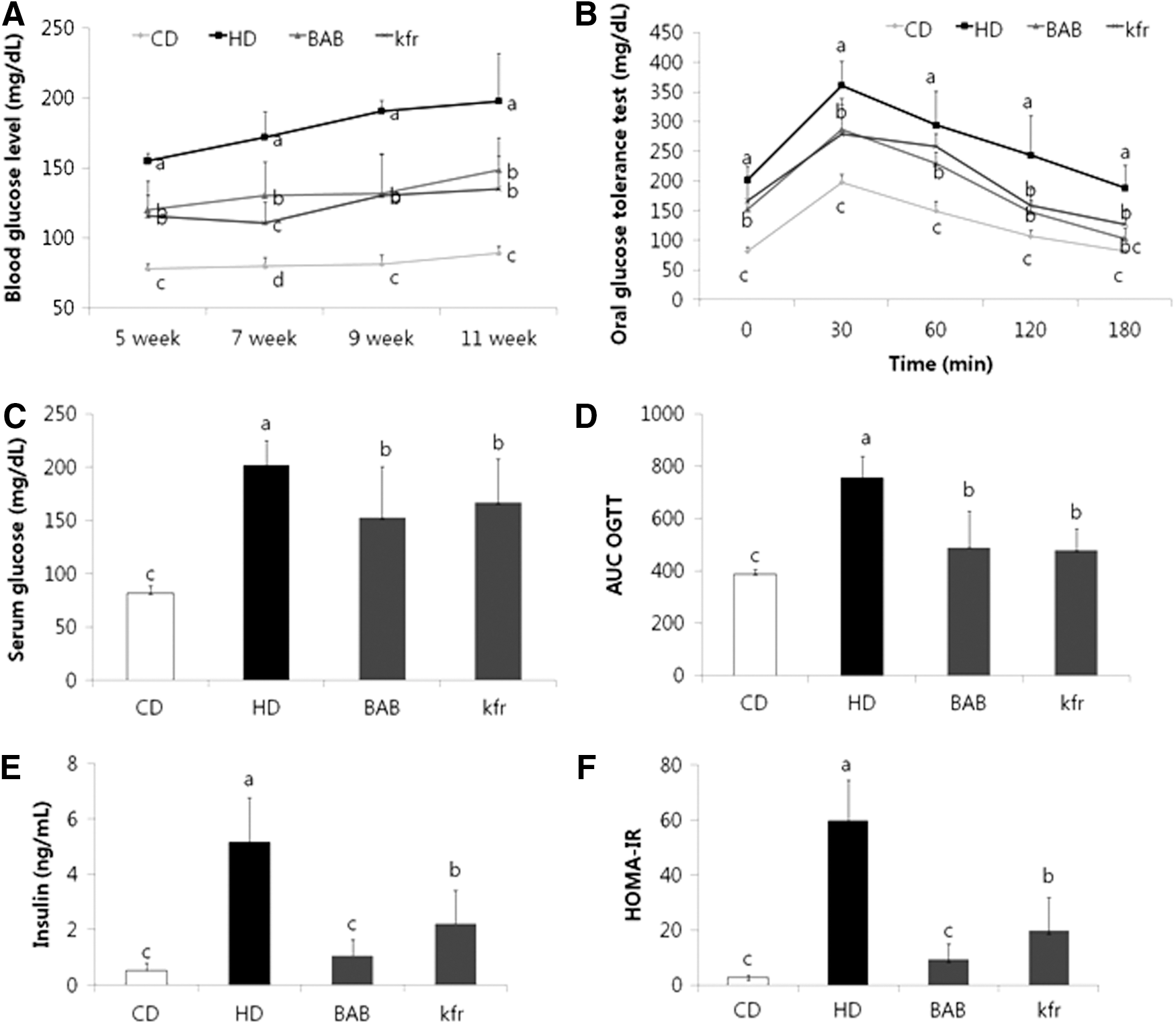
Effects of BAB on serum glucose, glucose tolerance, insulin levels, and HOMA-IR in diet-induced obese mice.
Furthermore, the OGTT showed that the blood glucose levels were significantly lower in BAB extract- and Kfr-treated mice than in HD mice 180 min after glucose loading (Fig. 4B). Likewise, the AUC for the 3-hour blood glucose response was significantly lower in BAB extract-fed mice than in HD mice (Fig. 4D).
In addition to the results already described, HD mice had higher serum insulin concentrations than CD mice, whereas BAB extract-fed mice had lower serum insulin levels than HD mice (by 80%), which was similar to that of CD mice (Fig. 4E). Consistently, the insulin resistance index, HOMA-IR, indicated that HD mice had a higher HOMA-IR than CD mice, whereas BAB extract-fed mice exhibited an 84% lower HOMA-IR than HD mice (Fig. 4F).
Gene expression profiling
More intensive examination of the effects of BAB extract at the molecular level was carried out by analyzing hepatic gene expression profiles between HD mice and BAB extract-fed mice compared with CD mice. Specifically, we identified 4373 genes that were differentially expressed among CD, HD, and BAB groups, among which 1440 genes were differentially expressed by at least two-fold. Further functional annotation analysis emphasized the ability of BAB extract to regulate glucose metabolic processes and alter the expression of insulin response genes. Specifically, expression of pyruvate dehydrogenase kinase (Pdk4), chitinase 3-like 3 (Chi3l3), and peroxisome proliferative activated receptor, gamma, coactivator 1 alpha (Ppargc1a) was lower in mice supplemented with BAB extract than in mice in the HD group, whereas the expression of protein phosphatase 1 regulatory subunit 3B (Ppp1r3b), insulin receptor substrate (Irs2 and 4), sortilin 1 (Sort1), and forkhead box P1 (Foxp1) genes was upregulated by BAB extract treatment compared with animals in the HD group (Table 2).
Fold: Fold changes in CD versus HD and CD versus BAB animals. CD, control diet containing 10% kcal fat; HD, high-fat diet control containing 60% kcal fat; BAB, high-fat diet with 1% freeze-dried ethanolic extract of black adzuki bean.
Discussion
Adzuki beans have traditionally been consumed as food crops and folk medicines in many regions of Asia, 25 and previous studies have demonstrated that adzuki beans have hypoglycemic effects. For example, adzuki bean extract significantly lowers blood glucose and serum insulin levels in the diabetic KK-Ay mouse model. 20,21 In this study, we sought to identify the cascade of events by which BAB alters glucose levels during the insulin response. In pursuing this goal, we took advantage of the fact T2D is strongly associated with hyperglycemia, insulin resistance, and obesity.
Kfr is a flavonol that has been identified in many plants used in traditional medicine,
26
including the adzuki bean
11,12
and BAB (Supplementary Fig. S1; Supplementary Data are available online at
At the end of the experimental period (week 12), we noted significantly increased insulin levels in the HD group together with significantly increased fasting glucose levels, which likely reflected typical hyperinsulinemia related to HD-induced T2D and obesity. Importantly, BAB supplementation reduced fasting glucose and insulin concentrations (Fig. 4), which may be due to increased expression of insulin receptor-related genes (Table 2) with consequential improvments in glucose homeostasis.
Furthermore, hyperglycemia deteriorates peripheral insulin sensitivity and pancreatic insulin secretory capacity. 27,28 In this study, we found that BAB was able to protect pancreatic β cells and regulate genes associated with the insulin response. A previous study reported the effects of culturing pancreatic cells with various concentrations of glucose with respect to insulin secretion. 29 Elevation of glucose levels stimulates insulin release; however, chronic exposure of pancreatic cells to high glucose conditions results in impairment of both cell viability and insulin secretory function. 6 Therefore, we confirmed the ability of BAB to enhance cell viability, proliferation, and glucose-stimulated insulin secretion in pancreatic β cells cultured under high glucose incubations. Furthermore, BAB extract treatment increased glucose-induced insulin secretory capacity. Using an animal model, we found that the HOMA-IR index was increased in HD mice than in CD mice, whereas BAB extract-fed mice showed an 80% lower HOMA-IR than HD mice. Decreasing β-cell function can be modeled as a function of changing β-cell response to blood glucose concentrations. 30 A significant component of the pathogenesis of T2D is low peripheral tissue sensitivity to insulin activity. 1 Importantly, we found that BAB treatment increased the expression of Irs2, 4, and Sort1 genes. Irs2 signaling promotes regeneration of β cells and control of homeostasis, which can prevent obesity and diabetes in mice. 31 Chronic hyperinsulinemia downregulates the mRNA for Irs2, an essential component of the insulin signaling pathway in the liver, thereby resulting in insulin resistance. 32 The expression of Sort increases formation of glucose transporter 4, which is localized in specialized storage vesicles and stimulates insulin-regulated glucose uptake, whereas knockdown of Sort decreases the expression of both genes. 33
In addition to the molecular changes already described, BAB appears to influence glucose homeostasis in other ways. Hepatic glucokinase (Gck) catalyzes the phosphorylation of glucose to glucose 6-phosphate, which is an essential step in glucose metabolism in the liver as well as for the induction of glycolytic genes. 34 In this study, Gck expression was similar between HD and BAB, whereas Pdk4 expression was downregulated by BAB compared with HD. The pyruvate dehydrogenase complex (PDC) regulates the entry of glycolytic products into the tricarboxylic acid cycle by catalyzing the oxidative decarboxylation of pyruvate to acetyl-coenzyme A in the mitochondria. 35 The activity of PDC is strongly inhibited by phosphorylation of its dehydrogenase component by Pdk, 36 and high-fat-fed and insulin-resistant mice lacking Pdk4 exhibit improved blood glucose levels and better glucose tolerance than wild-type mice. 37,38 In addition to Pdk4, Ppp1r3b is a major protein that regulates liver glycogen metabolism. 39 In glycogen metabolism, glucose binds to activated glycogen phosphorylase (GP) and promotes its dephosphorylation and inactivation, thereby terminating the inhibition of glycogen synthase phosphatase by GP such that glycogen can be resynthesized when blood glucose is high after a meal. 40 Mice expressing a mutant form of Ppp1r3b that cannot be inhibited by GP consistently exhibit enhanced activation of hepatic glycogen synthase and conversion of blood glucose into hepatic glycogen. 41 Accordingly, our genome-wide expression analysis suggested that BAB plays a significant role in regulating glucose metabolism.
In contrast, several genes (phosphoenolpyruvate carboxykinase, Pck; glucose-6 phosphatase, G6pc) involved in gluconeogenesis regulations are less suppressed with BAB group than with HD group. Foxp1 is a transcriptional repressor and is crucial for the governance of systemic glucose homeostasis. A recent study showed that hepatic overexpression of Foxp1 triggers the inhibition of the expression of the key gluconeogenic genes, including Ppargc1a, 42 which activates an entire program of gluconeogenic enzymes production. 43 Hormonal and nutrient regulation of hepatic glucose production primarily occurs through modulation of Ppargc1a and related complex networks. 44 In diabetic mice, the hepatic expression of Foxp1 has been found to be downregulated leading to increased hepatic gluconeogenesis. 42 Interestingly in our study, the BAB supplementation was effective in upregulating the Foxp1 expression, thereby downregulating Ppargc1a expression (Supplementary Fig. S2) and suppressing the gluconeogenic pathway. In summary, our in vitro and in vivo data indicated that BAB extract improved glucose tolerance by protecting pancreatic β cells from damage due to chronic hyperglycemia and improving insulin responses. Together, these results suggested that BAB may be useful as a dietary supplement to regulate blood glucose.
Footnotes
Acknowledgment
This work was carried out with the support of “Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ009218)” Rural Development Administration, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.