
Abstract
Dietary fiber and proteins are individually known to decrease feeding, but could result greater weight management benefit when both are combined. We hypothesized that supplementing the diet with fermented barley, being rich in both dietary fiber and proteins, could lower energy intake by modulating the mRNA expression level of hypothalamic genes associated with the regulation of feeding behavior and satiety; thereby decreasing body weight gain. To test our hypothesis, four groups of Sprague Dawley rats were arranged in a 2 × 2 factorial design (n = 6), low-fat diet with either guar gum (LFD-G) or fermented barley (LFD-FB) and high-fat diet with either guar gum (HFD-G) or fermented barley (HFD-FB). Using oral gavage, fermented barley was given at a dosage of 1500 mg/kg body weight and guar gum was supplemented in an equivalent quantity to that of the fiber in the fermented barley. After 19 weeks, the fermented barley-supplemented groups showed a significant reduction in energy intake, triglyceride, body weight gain, and serum leptin, compared to the guar gum-supplemented groups in both the low- and high-fat diet groups. Likewise, the anorexigenic gene proopiomelanocortin (POMC) and cocaine and amphetamine-regulated transcript (CART) mRNA level were significantly higher in the fermented barley-supplemented groups compared to the guar gum-supplemented groups in rats fed on both high- and low-fat diets. In conclusion, fermented barley supplementation upregulated hypothalamic POMC/CART, decreased energy intake in both low- and high-fat diet groups, and prevented excessive weight gain in rats.
Introduction
O
Materials and Methods
Animals and diets
Male SD rats aged 4 weeks were purchased from Central Lab. Animal, Inc. (Seoul, Republic of Korea) and acclimatized for 10 days. Four groups were arranged in a 2 × 2 factorial design (n = 6): low-fat diet with either guar gum (LFD-G) or fermented barley (LFD-FB, 1500 mg/kg BW); high-fat diet with either guar gum (HFD-G) or fermented barley (HFD-FB, 1500 mg/kg). Guar gum was supplemented to equilibrate the fiber content to that of the fermented barley sample, which was 28%. The low-fat and high-fat diets (Research Diets, New Brunswick, NJ) were modified AIN-93 diets with 4% fat (10% calories from fat) content (OpenSource diet-D12450B) and 35% fat (60% calories from fat) content (OpenSource diet-D12492), respectively (Table 1). The animals were housed in a controlled environment at 23°C ± 1°C, relative humidity of 50% ± 5%, and 12-h light–12-h dark cycles. Animals were given free access to food and water during the entire 19-week experimental period. The experimental protocol was approved by the Animal Care and Use Committee of Chonbuk National University.
Low-fat diet: AIN-93 diet with 10% kcal from fat (OpenSource diet- D12450B).
High-fat diet: AIN-93 diet with 60% kcal from fat (OpenSource diet- D12492).
Preparation of fermented barley
Fermented barley was provided by Mealtobalance, Co., Ltd. (Jeongeup, Korea) and it was prepared as described earlier. 13 Briefly, the barley grain powders were dispersed in water and treated with α-amylase (Termamyl 120 L; Novozyme, Copenhagen, Denmark) and further liquefied at 93°C for 2 h; after cooling to 63°C, the mixture was treated with glucoamylase (AMG 300L; Novozyme) for 2 h for saccharification. The resultant product was centrifuged at 3000 g for 10 min (Combi-514R; Hanil Science Industrial Co., Ltd., Seoul, Korea) and the supernatant was transferred to the fermenter for fermentation (30°C, pH 5.5, 48 h) by Saccharomyces cerevisiae. After fermentation, the fermented material was centrifuged at 3000 g for 10 min and concentrated by vacuum evaporation (60°C, 1 h). The composition of fermented barley is given in Table 2.
Serum and tissue sample collection
After 12 h (21:00–9:00) of overnight fasting, each rat was deeply anesthetized with diethyl ether inhalation and blood was collected by decapitation. Serum was isolated from the clotted blood by centrifugation at 1100 g for 15 min at 4°C (Micro 17R; Hanil Science Industrial Co., Ltd.). Tissues were carefully excised, rinsed, weighed, and stored along with serum at −80°C until analyzed.
Serum hormones and lipid profile
Hormones were measured using commercially available kits, leptin; the Quantikine® Immunoassay Kit (R&D Systems, Inc., Minneapolis, MN), the Insulin Kit (Shibayagi, Shibukawa, Japan), and the Ghrelin Kit (Millipore, Billerica, MA) following the manufacturer's instructions. Serum lipid profiles were measured using a commercially available kit (Asan Pharmaceutical Co., Seoul, Republic of Korea).
Quantitative real time PCR analyses
Total RNA was extracted from the hypothalamus and liver tissue using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA) and the concentration was measured spectrophotometrically. The extracted RNA was reverse transcribed into complementary DNA (cDNA) using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA); the RNA expression level was quantified by using SYBR Green Polymerase Chain Reaction (PCR) Master Mix (Applied Biosystems) and the 7500 Real Time PCR system (Applied Biosystems) according to the manufacturer's protocol. Gene specific primers used in this study are given in Table 3. Relative quantification of gene expression with real-time PCR data was calculated relative to GAPDH and β-actin for hypothalamic and hepatic genes, respectively.
NPY, neuropeptide Y; AgRP, agouti-related peptide; POMC, proopiomelanocortin; CART, cocaine- and amphetamine-regulated transcript; MC4R, melanocortin receptor 4; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; PPARg, peroxisome proliferator-activated receptor gamma; PPARa, peroxisome proliferator-activated receptor alpha; CPT-Iα, carnitine palmitoyltransferase I alpha; ACO, acyl-CoA oxidase; β-actin, beta-actin.
Statistical analyses
Data from individual experiments are expressed as the mean ± SD. Analyses of variance were conducted to compare the effects of dietary fat and fermented barley as well as their interaction on the metabolic parameters. All of the statistical analyses were determined using SPSS 17.0 (SPSS, Inc., Chicago, IL). Statistical significance was considered at P < .05.
Results
Bodyweight, epididymal adipose tissue weight, anthropometric parameters, and energy intake
The final body weight (Table 4) was higher in HFD-fed animals than the LFD-fed animals indicating the diet effect (P < .05), but interestingly the final body weight was lowest in fermented barley-supplemented groups (P < .05) compared to the guar gum groups in both the diet regimen. The lower bodyweight gain as a result of fermented barley supplementation resulted in lower BMI (P < .05) and Lee Index (P < .05) in the fermented barley-supplemented groups compared to guar gum groups. Higher epididymal adipose tissue (EAT) weight was observed in the HFD groups than LFD groups P < .05).
All values are given as mean ± standard deviation (SD) (n = 6). The degree of significance resulting from the two-way ANOVA is shown with the effect if Supplement (S), Diet (D), and the interaction of Supplement and Diet (S×D) being expressed as a numerical value or as NS when P > .05. Energy intake (kcal/rat) = [Total diet intake (g) of the group × Energy per gram diet (kcal/g)] ÷ N. Energy per gram for LFD = 3.8 kcal/g; HFD = 5.2 kcal/g
HFD-FB, fermented barley; HFD-G, high-fat diet with either guar gum; LFD-FB, fermented barley; LFD-G, low-fat diet with either guar gum; NS, not significant.
The energy intake (Table 4) was lower in the LFD groups compared to the HFD groups (P < .05); however, when compared between the two supplements, it was observed that the energy consumption was lower in the fermented barley-supplemented groups as compared to guar gum supplementation (P < .05).
Effect of fermented barley on serum lipid profile
The serum triglyceride (TG), total cholesterol (TC), and high-density lipoprotein (HDL)-cholesterol (Table 5) levels were higher in the low-fat diet-fed group compared to high fat-diet group suggesting the diet effect (P < .05). The TG level was also influenced by the fermented barley supplementation (P < .05) and the interaction between the diet and the supplementation (P < .05); however, supplementation and its interaction with diet did not change TC and HDL-c.
All values are given as mean ± SD (n = 6). The degree of significance resulting from the two-way ANOVA is shown with the effect if Supplement (S), Diet (D), and the interaction of Supplement and Diet (S×D) being expressed as a numerical value or as NS when P > .05.
HDL, high-density lipoprotein; TC, total cholesterol; TG, triglyceride.
Hormone levels
Serum insulin, leptin, and ghrelin levels are shown in Figure 1. Serum insulin concentrations did not show any significance. We observed lower circulating serum leptin in the LFD than HFD group (P < .05); however, on comparing the two supplements it was noticed that fermented barley-supplemented groups in LFD and HFD regimen had less circulation leptin as compared to the guar gum in their respective diets (P < .05). The serum total ghrelin level was lower in the HFD groups compared to the LFD groups (P < .05); however no significant effect of fermented barley was observed in either group.
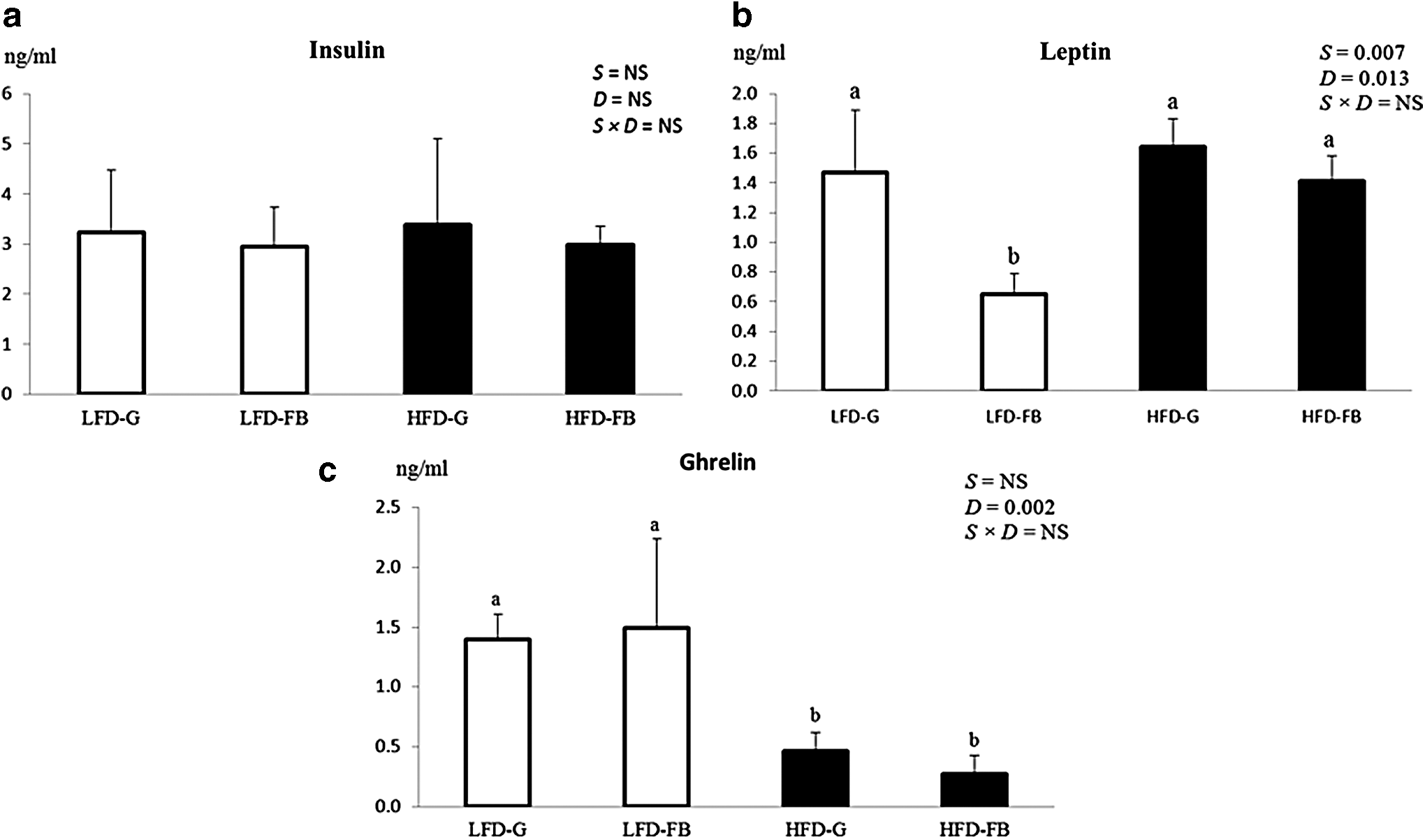
Serum levels of
Hypothalamic gene expression
The hypothalamic genes involved in appetite regulation are shown in Figure 2. In our study the agouti-related peptide (AgRP) mRNA level of animals in fermented barley-supplemented group was higher in the LFD background, whereas lower in the HFD background indicating the effect of diet (P < .05) as well as the interaction between diet and the supplementation (P < .05). The level of neuropeptide Y (NPY) and melanocortin 4 receptor (MC4R) mRNA did not show any significant difference among any of the groups. The mRNA expressions of anorexigenic neurons releasing proopiomelanocortin (POMC) and cocaine and amphetamine-regulated transcript (CART) were higher in the fermented barley-supplemented group than guar gum-supplemented group under both LFD and HFD diet regimen, respectively (P < .05); however the CART mRNA expression level was also influenced by the interaction between diet and supplement (P < .05).

Hypothalamic gene expression of
Hepatic lipid metabolism gene expression
The hepatic genes involved in lipid metabolism are shown in Figure 3. In our study fatty acid synthase (FAS) mRNA expression was downregulated in fermented barley supplementation group compared to guar gum fed under both diet regimens (P < .05). Peroxisome proliferator-activated receptor (PPARG) γ mRNA expression was higher in the HFD group than LFD groups, indicating the diet effect (P < .05) rather than the supplementation. The mRNA levels of PPARα along with carnitine palmitoyltransferase 1α and acyl-CoA oxidase (ACO) were lower in the fermented barley-supplemented group than the guar gum group (P < .05) in both diets; however, ACO mRNA expression was also influenced by diet (P < .05) and the interaction between diet and supplement (P < .05).
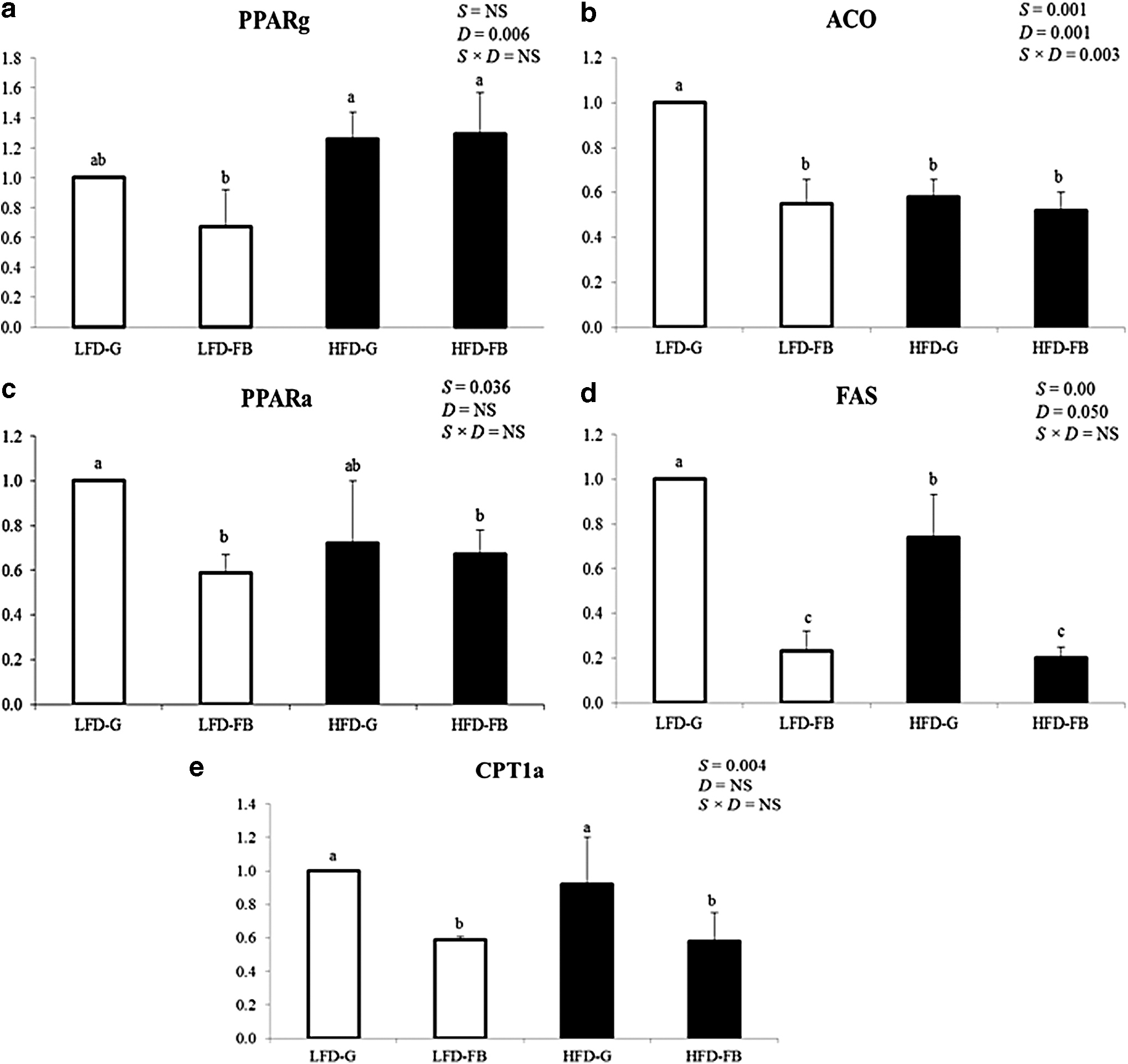
The hepatic lipid metabolic gene expression
Discussion
We investigated the effect of fermented barley supplementation on obesity-related parameters in rats fed with two different diet regimens, LFD and HFD. The fermented barley-supplemented LFD and HFD groups gained 11.27% and 12.43% less body weight, respectively, as compared to the guar gum-supplemented group. Consumption of high-calorie diets promotes adiposity. 20,21 Although not significant, the epididymal adipose mass was lower by 11.51% and 6.62% in animals supplemented with fermented barley compared to guar gum under both LFD and HFD regimen, respectively. As a lower body weight was observed in the fermented barley groups with insignificance in epididymal adipose weight, the supplementation might have affected adipose tissue from other depots. A similar study showed that the intake of dietary fiber was effective in lowering the mesenteric fat, but not epididymal fat. 22
Lesser energy consumption has a strong correlation with lower body weight gain. It was observed that supplementation of fermented barley reduced the energy intake by 4.37% in LFD and 7% in HFD rats as compared to guar gum. The fermented barley contained dietary fiber (28%) and protein (25.4%); dietary fibers, such as dextrin, nutriose, and flaxseed fiber added to beverages have been shown to decrease appetite and energy intake in humans. 23 –26 Similarly, proteins modulate appetite-regulating gene expression causing decreased food intake and increased satiety; a study with mice showed that whey protein isolate decreased energy intake. 27 In accordance with these studies, fermented barley-supplemented animals in our study showed less energy intake than guar gum with both diets.
The serum insulin level did not show any significant differences among the groups; this result was similar to the findings of Artiss et al. 28 suggesting that soluble dietary fiber intake did not result in any significant changes in serum insulin levels. Leptin is an adipokine secreted by adipose tissue with EAT as a major contributor. 29 In this study we observed that the fermented barley group had 55.78% and 14.02% of lower circulating serum leptin than guar gum in both in LFD and HFD diets, respectively. The reason behind low levels of circulating leptin in the two fermented barley groups could be due to the lower energy intake and EAT weight. Ghrelin, secreted by gastric oxyntic mucosal cells acts on hypothalamus and induces appetite in both animals and humans. 30 Diets rich in high fat have been shown to reduce serum ghrelin levels, 31 and foods rich in dietary fiber reduce the expression of ghrelin genes. 32 The ghrelin-suppressing nature of dietary fiber in both fermented barley and guar gum groups might have resulted in similar serum ghrelin levels between the fermented barley and guar gum-supplemented groups fed on both the LFD and HFD.
The hypothalamic circuitry plays an important role in regulating energy intake by governing the appetite-regulating genes. The AgRP and NPY are orexigenic neuropeptides, 33 colocalized in arcuate nucleus and stimulate feeding in animals. 34 A study by Gaysinskaya et al. 19 suggested that sucrose has a strong stimulatory effect on AgRP mRNA, therefore, it could be possible that sucrose in LFD might have masked the effect of fermented barley supplementation resulting in its upregulation. In one study, mice supplemented with a protein-enriched diet showed no significant alterations in NPY expression levels in comparison with those fed with isocaloric starch-enriched diet. 35 Similarly, in our study, animals supplemented with protein and fiber-rich fermented barley did not show any significant difference in the NPY mRNA expression level as compared to guar gum.
The hypothalamic melanocortin system consists of MC4R—a common receptor for agonist αMSH (melanocyte-stimulating hormone) and antagonist AgRP 36 Adan et al. studies with knockout mice showed that MC4R is essential for the coordination of energy intake and substrate oxidation. 37 Although in our study we did not see any significance in the MC4R mRNA expression level, the outcome was dependent on POMC, the precursor of its agonist to αMSH. 35
POMC and CART are involved in reducing the food intake. 38 In this current study, the fermented barley-supplemented group showed 98% and 14% higher expressions of CART along with 58% and 35.7% increased POMC mRNA expression than the guar gum group under both LFD and HFD diet, respectively; the higher expression of CART/POMC mRNA might be due to the presence of dietary fiber in the fermented barley along with protein. We believe that there might be at least two possible reasons; diets rich in proteins and amino acids change the expression of hypothalamic appetite-regulating gene expression through mTOR (mammalian target of rapamycin) and AMPK (AMP-activated protein kinase)-dependent mechanism in the hypothalamus, causing increased expression of POMC and decreased expression of NPY and AgRP resulting in decreased feed intake. 12,39 –41 The other possible reason could be the fermentation of dietary fibers in the animal's gut. A study using an in vitro model of the porcine gastrointestinal tract suggested that, barley, upon fermentation in the intestinal tract, produces short-chain fatty acid (SCFA) including acetate. 42 While another study, using a canine model, suggested that intestinal fermentation of guar gum produces propionate as a major SCFA. 43 A recent study reported that gut fermentation of dietary fibers suppresses appetite, 44 through production of acetate that stimulates the production of POMC mRNA and suppresses AgRP mRNA expression. 45 Therefore, the high amounts of dietary fiber (28%) and protein (25.4%) in the fermented barley might act synergistically to decrease energy intake.
Apart from analyzing hypothalamic genes regulating feeding, we also measured the expressions of hepatic genes involved in lipid metabolism. The lipid anabolic genes PPARγ and FAS are crucial for TG synthesis. LFD rich in sucrose have been reported to increase the FAS mRNA expression level, 46 and similar effects were observed in our study, as compared with HFD. Interestingly, supplementing fermented barley in both diets lowered the expression of FAS. High fat upregulated the PPARγ mRNA expression in the HFD group, which was reflected in a higher EAT weight in the HFD group compared to the LFD group. Higher PPARγ expression has been found to correlate with lower circulating TG and higher adiposity. 47,48 The PPARα gene regulates lipid catabolism through governing the expression of genes for the enzymes involved in β-oxidation of fatty acids in mitochondria and peroxisome, such as CPT-1α and ACO, respectively. We observed that, PPARα expression was lower in the fermented barley-supplemented group than the guar gum-supplemented group. A lower PPARα was reflected in the down-regulation of the CPT-1α and ACO in the fermented barley-supplemented group in both diet regimens. The differential expression of the lipid metabolic gene between the guar gum and fermented barely group could be due to the difference in the end product resulting from gut fermentation of the supplements. Gut fermentation of guar gum and barley produces propionate 43 and acetate, 42 respectively. In vitro and in vivo studies using propionate revealed that SCFA upregulates the expression of various genes involved in lipid metabolism, including PPARα, 49 FAS, and PPARγ. 50
The serum TG, TC, and HDL cholesterol levels were higher in the LFD group compared to HFD group suggesting the diet effects. Interestingly, fermented barley-supplementation group showed lower levels of TG in both diet regimens indicating ameliorating effects of fermented barley supplementation. One of the reasons behind higher TG levels in the low-fat diet could be due to the higher sucrose content in LFD than HFD. Our data are in congruence with the report 51 published earlier suggesting TG level raising effect of a sucrose diet. On the other hand, lower TG level in the fermented barley groups could be due to less energy intake by the group as compared to the guar gum-supplemented group.
In conclusion, fermented barley with dietary fiber and protein promoted the upregulation of the POMC/CART mRNA levels in animals more than the guar gum, resulting in less energy intake leading to lower weight gain and obesity. Further study, however, will be carried out with an acetate-supplemented group for elucidating the metabolic pathway.
Footnotes
Acknowledgments
This research was financially supported by the Ministry of Knowledge Economy (MKE) and Korea Testing & Research Institute (KTR) through the support project of healthcare material industry for international mutuality authentication. The author thanks Dr. Janardhan P. Bhattarai, Department of Oral Physiology, Chonbuk National University, for his technical assistance.
Author Disclosure Statement
No competing financial interests exist.