
Abstract
Nonalcoholic fatty liver disease (NAFLD), which is characterized by >5% deposition of triglycerides in hepatocytes, is often referred as a major risk factor for obesity, type 2 diabetes, and hypertension. We investigated the hepatoprotective effect of whole soybean embryos containing bioactive substances such as isoflavones and soyasaponins. For this study, mice were randomly allocated into four groups that were fed different diets for 10 weeks: normal diets and high-fat and high-cholesterol diets (HD), and HD with 10% or 20% soybean embryo powder (10SE-HD and 20SE-HD). Hepatic superoxide dismutase and glutathione peroxidase activity of the experimental groups increased during the period of the study (P < .05). Hepatic mRNA expressions of tumor necrosis factor α, nuclear factor (erythroid-derived 2)-like 2, and Caspase 3 were decreased when soybean embryos were increased in the mice's diets. Both of the soybean embryo-treated groups showed significantly decreased serum and liver triglyceride and total cholesterol. Adiponectin, AMP-activated protein kinase (AMPK) α, hydroxymethylglutaryl-CoA reductase, sterol regulatory element-binding protein-1c, fatty acid synthase, and apolipoprotein B mRNA expressions were decreased in the mice that were fed soybean embryos. We suggest that the regular supplementation of soybean embryos might be a useful treatment for preventing NAFLD and associated complications through upregulation of adiponectin-mediated AMPKα pathway parameters, which are implicated in antioxidant, anti-inflammatory, and lipid metabolism activities.
Introduction
N
The prevalence of NAFLD is explained by a “two-hit hypothesis”. 9,10 The first hit is presumed to come from triglyceride (TG) accumulation within hepatocytes, which ends in simple steatosis. Hepatic steatosis is easily exacerbated due to an accelerated peripheral lipolysis and increased cellular vulnerability to a variety of stress factors, increasing further injury to the liver. 8,11 –13 Examples of this include fatty acid-derived oxidative stress, environmental toxins, or proinflammatory cytokines arising from the portal venous system leading to endoplasmic reticulum stress and the activation of hepatic stellate cells, which is the key event of hepatic fibrosis. 11,14
The second hit includes primary lipotoxicity caused by oxidative stress from increased lipid peroxidation, high reactive oxygen species production within hepatocytes, mitochondrial dysfunction, and inflammation. Other factors leading to liver injury include oxidative stress and lipid peroxidation, decreased antioxidant defenses, early mitochondrial dysfunction, iron accumulation, imbalance of adipokines, adipose-derived cytokines with chronic inflammation, and gut-derived microbial adducts. 8,15 These biological processes may compromise liver status and progress into steatohepatitis and fibrosis due to the activation of hepatic stellate cells. 16 –18
Adiponectin can enhance AMP-activated protein kinase alpha (AMPKα) actions 19 and suppress proinflammatory cytokines like tumor necrosis factor (TNF) α as well as nuclear factor kappa B (NF)-κB. 20 In a previous study, the downregulation of adiponectin can lead to an increase in the induction of inflammation and lipid accumulation in the liver. 20,21 Upregulation of AMPKα leads to further increase of fatty-acid oxidation through upregulating carnitine palmitoyltransferase (CPT)-1 expression and impedes lipid synthesis. 22 –24 In addition, AMPKα activation, which reduces expression of acetyl-CoA carboxylase (ACC) and fatty acid synthase (FAS) through suppression of sterol regulatory element-binding protein (SREBP)-1c, triggers decreased hepatic lipid synthesis (Fig. 1).

Mechanism of antioxidant activity and lipid metabolism-associated factors by soybean embryos in NAFLD. We hypothesized that supplementation of soybean embryos could improve parameters, which are implicated in antioxidant, anti-inflammation, and lipid metabolism, and related NAFLD. NAFLD, nonalcoholic fatty liver disease. Color images available online at
Previous studies have demonstrated that a significant correlation exists between the abundant intake of soy foods and low incidence of several chronic diseases, for example, sex hormone-dependent cancers, coronary heart disease, osteoporosis, and postmenopausal discomfort. Scientists presume that isoflavones contained in soy foods are major components responsible for preventive actions against these chronic conditions. 25,26
Soybeans contain three primary isoflavones, genistein (4′,5,7-trihydroxyisoflavone), daidzein (4′,7-dihydroxyisoflavone), and glycitein (7,4′-dihydroxy-6-methoxyisoflavone) and the β-glycosides, genistin, daidzin, and glycitin. Soybean embryos also have a high percentage of oils, proteins, phytochemicals, phytosterols, and phytic acid. Soybean embryo contains more isoflavones (30–60 mg), saponins, phytosterols, linolenic acid, iron, zinc, folate, and vitamin E on a per gram basis than soy flour, which contains only 1–2 mg of isoflavones. 27 In addition, Soybean embryos contain 7.5 times more isoflavone and 3.1 times more soyasaponin than cotyledon. 28 –30 Recently, soy germ was in the spotlight of several studies demonstrating an estrogenic activity, 31 osteoporosis, 32 menopausal symptom relief, 33 cholesterol control, 34 and antioxidant effects. 35
Soyasaponin is known to have important effects on the antioxidant activity, such as reducing oxidant stress, 36 antivirus activities leading to virus suppression, 37,38 a protective effect on the liver against liver damage, 39,40 cancer prevention by reducing DNA damage, 41 and inhibiting the incidence of aberrant crypt foci in the colon of mice. 42
This study aimed to identify the positive effects of soybean embryos on the antioxidant activity and lipid metabolism in high-fat and high-cholesterol diet-induced NAFLD in C57/BL6 mice. Many studies have suggested that the consumption of isoflavone and soyasaponin has positive effects on NAFLD-related metabolism. Judging from these results, we hypothesized that isoflavone and soyasaponin-rich soybean embryo intake could have positive effects on the development of NAFLD.
Material and Methods
Animals and diets
Six-week-old C57/BL6 male mice were purchased from Saeronbio, Inc. (Gyeonggi, Korea). Mice were housed under controlled temperature (21°C ± 1°C) and humidity (55% ± 5%) with a 12-h light–12-h dark cycle. All animal procedures were performed in accordance with the guidelines issued by Sookmyung Women's University for the care and use of laboratory animals (SMWU-IACUC-1404-006-001). After acclimatization for 1 week, the C57/BL6 mice (n = 7 per group) were randomly allocated to four groups (Table 1): normal diet (ND), high-cholesterol and high-fat diet (HD), HD with 10% soybean embryo powder (10SE-HD), and HD with 20% soybean embryo powder (20SE-HD). The feed was purchased from FEEDLAB Co. (Gyeonggi, Korea) and supplemented with soybean embryo powder for 10 weeks. Mice were fed an AIN-93G diet, or an HD. The dried soybean embryo powders were mixed with their diets. The food intake and body weight of the mice were measured daily and weekly.
ND (n = 7); HD (n = 7); 10SE-HD, HD +10% SE powder (n = 7); 20SE-HD, HD +20% SE powder (n = 7).
HD, high-cholesterol/fat diet; ND, normal diet; SE, soybean embryo; TBHQ, tert-butylhydroquinone.
Preservation of blood and tissue samples
At the end of the study, following overnight fast, final body weight was measured, and the mice were euthanized. The blood samples were collected in tubes and centrifuged at 1600 ×g for 30 min at 4°C allowing the serum to be collected. The serum was stored at −70°C until analysis. The heart, kidney, lung, liver, abdominal adipose tissue, and epididymis adipose tissue were removed, rinsed with distilled water, weighed, and stored at −70°C until analysis.
Superoxide dismutase, glutathione peroxidase, catalase activity in the liver
Superoxide dismutase (SOD), glutathione peroxidase (GPx), and catalase activities were measured using an SOD Assay kit-WST (S311-10, Dojindo Molecular Technologies, Inc., Rockville, MA, USA), a Glutathione Peroxidase Activity Colorimetric kit (K762-100, BioVision, Inc., Milpitas, CA, USA), and a Catalase Activity Colorimetric/Fluorometric kit (K773-100, BioVision, Inc.), respectively. All experiments were analyzed in triplicate, and the average values are presented.
Lipid parameter measurements
Total cholesterol (TC) and TG levels were determined using a TG-S kit (3I1570; Asanpharm, Hwaseong, Korea) and a T-CHO kit (3I2020; Asanpharm). The high-density lipoprotein (HDL) and low-density lipoprotein (LDL)/very low-density lipoprotein (VLDL) cholesterol levels were determined using an HDL and LDL/VLDL Quantification Colorimetric/Fluorometric kit (K613-100; BioVision, Inc.). The free fatty acid (FFA) content was determined by the Free Fatty Acid Quantification Colorimetric/Fluorometric kit (BioVision, Inc.). Liver TC and TG levels were determined using Folch's method. 43 Serum TC, TG, HDL, LDL/VLDL, and FFA levels were measured with an Epoch Microplate Spectrophotometer and an ELISA reader (BioTek Instruments, Inc., Winooski, VT, USA).
Reverse transcriptase polymerase chain reaction
The RNA was isolated from liver homogenates using an Easy-Blue™ [DNA free] Total RNA Extraction kit (iNtRON biotechnology, Inc., Gyeonggi, Korea) following the manufacturer's protocol. Reverse transcriptase polymerase chain reaction (RT-PCR) was performed in a Thermal cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA) using a Maxim RT-PCR Premix kit (iNtRON biotechnology, Inc.). Band intensities were evaluated using gel quantifier image analysis (GelQuant), version 2.7 beta. Details of all primers used are provided in Table 2.
AMPKα, AMP-activated protein kinase alpha; Apo B, apolipoprotein B; FAS, fatty acid synthase; HMG-CoA-r, 3-hydroxy-3-methylglutaryl-coenzyme A reductase; NF-κB, nuclear factor kappa B; Nrf2, nuclear factor (erythroid-derived 2)-like 2; SREBP-1c, sterol regulatory element-binding protein-1c; TNFα, tumor necrosis factor alpha.
Statistical analysis
The results were expressed as the means ± standard errors (SE). The statistical analyses of the parametric data were based on a one-way ANOVA followed by Tukey's test. The limit of significance was set at P < .05. The statistical analyses were conducted using GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA) software.
Results
Weight and intake parameters
At the beginning of this study, there were no significant differences in body weight in the four groups of mice. At the end of the experimental period, the animals fed the HD showed an ∼42% increase in weight gain compared to the ND group. In contrast to the mice fed the HD, body weight was significantly decreased in the mice fed the 10SE-HD and 20SE-HD (P < .0001). There was also a significant increase in body weight gain of the mice fed the HD (P < .0001) compared with the mice fed the ND. Soybean embryo supplementation of the mice's diet (P < .0001) reduced body weight gain, compared with the HD (P < .0001). There was a statistical difference in total food intake among the groups; Mice from groups 10SE-HD and 20SE-HD (P < .0001) demonstrated a higher food intake than the HD group (Table 3). The kidney (P < .01), liver (P < .0001), visceral (P < .0001), and epididymis adipose tissue (P < .0001) weight significantly decreased in a dose-dependent manner in all experimental groups, compared to those fed the HD. The relative liver weight was significantly decreased in mice fed 20SE-HD compared to those fed HD (P < .0021) (Fig. 2). Visceral (P < .0001) and epididymal adipose tissue (P < .0001) weights decreased in the experimental groups supplemented with soybean embryo (Table 4).
Values are expressed as mean (SEM). ND (n = 7); HD (n = 7); 10SE-HD, HD +10% SE powder (n = 7); 20SE-HD, HD +20% SE powder (n = 7).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD; P < .05).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD, P < .001).
Values are expressed as mean (SEM). ND (n = 7); HD (n = 7); 10SE-HD, HD +10% SE powder (n = 7); 20SE-HD, HD +20% SE powder (n = 7).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD, P < .05).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD, P < .01).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD, P < .001).
BW, body weight.
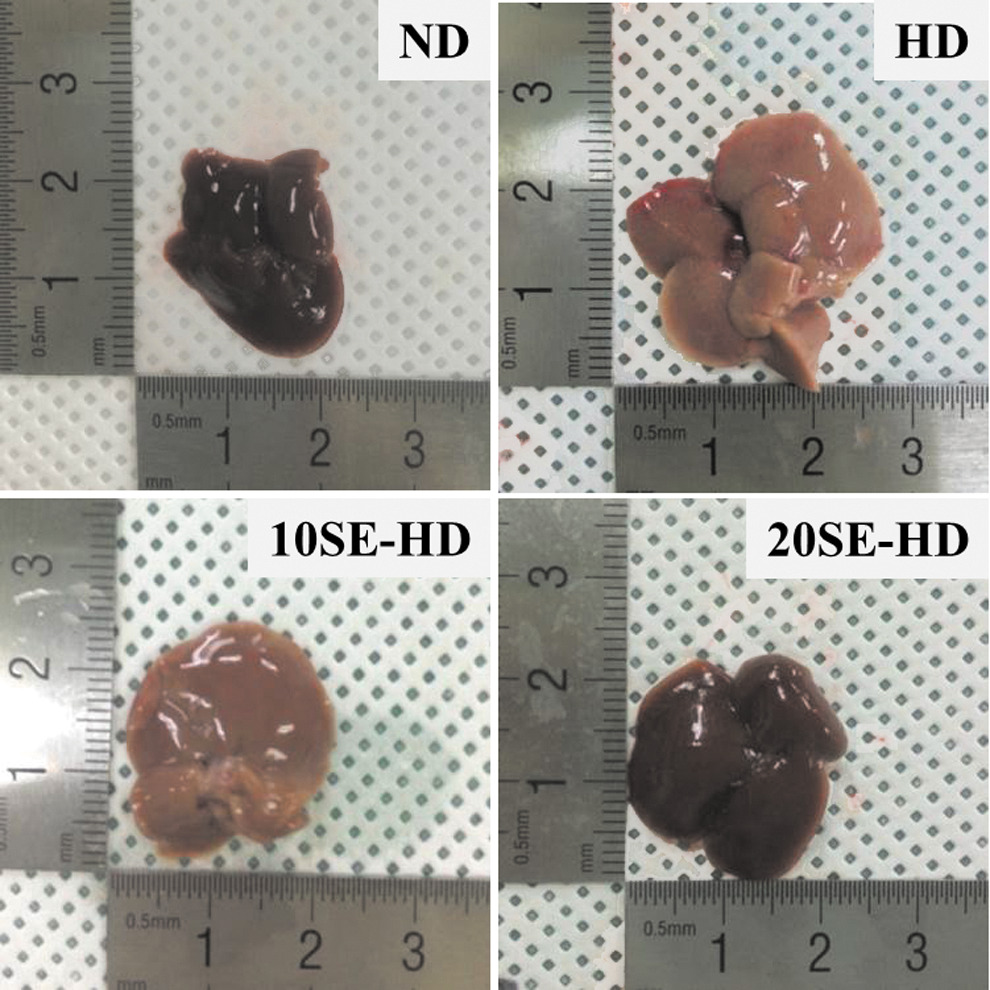
Liver morphology of mice fed Experimental diets for 10 weeks. 10SE-HD, HD +10% SE powder, and 20SE-HD, HD +20% SE powder. HD, high-cholesterol/fat diet; ND, normal diet; SE, soybean embryo. Color images available online at
SOD, GPx, and catalase activity
The antioxidant values increased in the animals fed the soybean embryo powder-supplemented food. The 10SE-HD and 20SE-HD groups showed significant increases in the SOD activity (P < .05 and P < .0001), compared to the HD group (Fig. 3A). The hepatic GPx activity showed a 1.83 times increase in the 20SE-HD (P < .0001), compared to the HD-fed mice (Fig. 3B). However, the hepatic catalase activity was not significantly different (Fig. 3C).
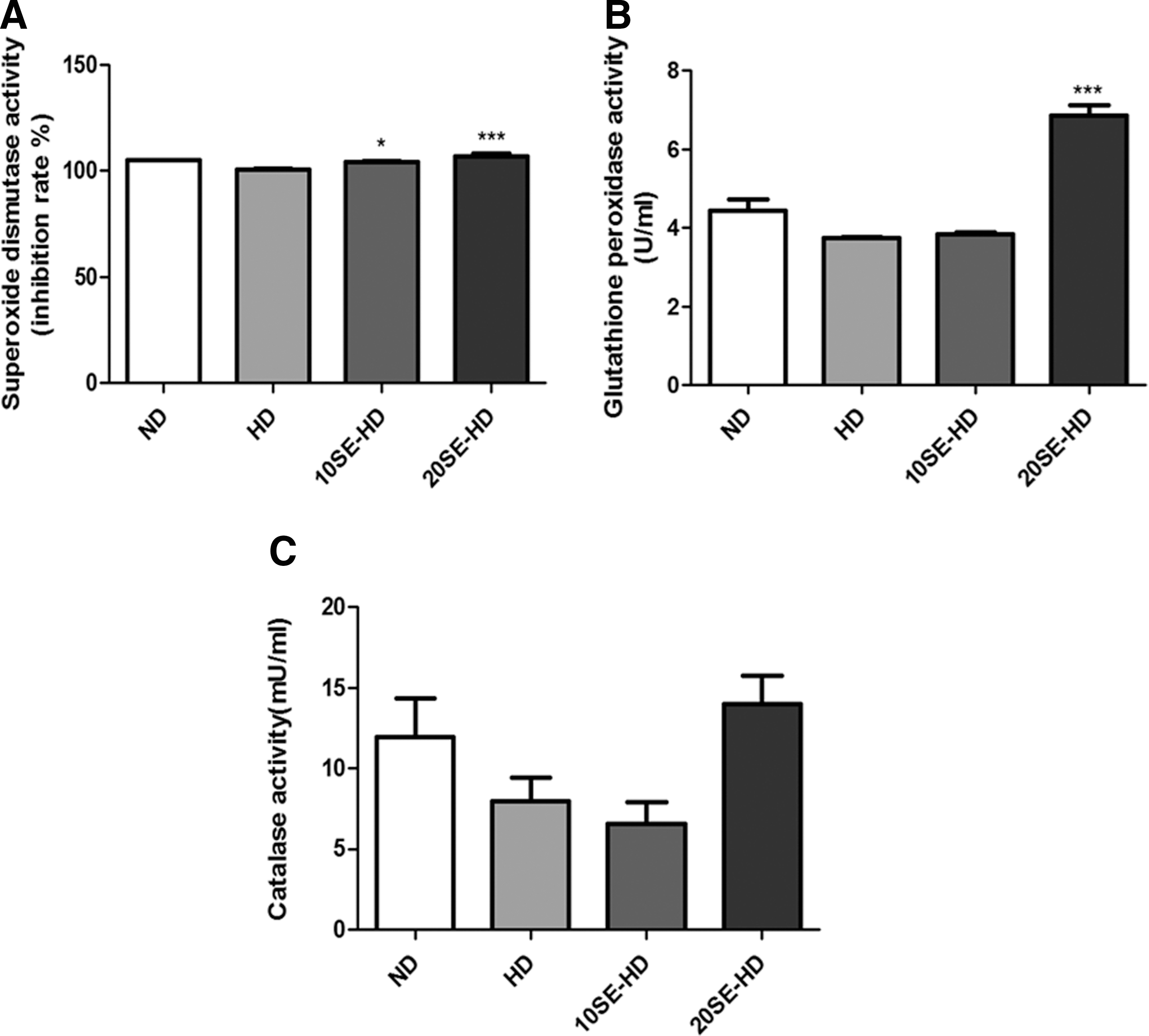
Antioxidant parameters in the liver.
TNFα, Nrf2, NF-κB, and caspase 3 mRNA expression
Expression of TNFα, nuclear factor (erythroid-derived 2)-like 2 (Nrf2), and caspase 3 was significantly lower in the experimental groups. The TNFα expression in the liver showed a significant decrease in the two experimental groups (P < .0007), compared to the HD group (Fig. 4A). Hepatic Nrf2 expression was lower in the treatment groups, 10SE-HD and 20SE-HD (P < .05 and P < .0044), compared to the HD group (Fig. 4B). Caspase 3 expression in the experimental groups (P < .01 and P < .0001) decreased when compared to the HD-fed group (Fig. 4D). However, NF-κB expression was not significantly different (Fig. 4C).
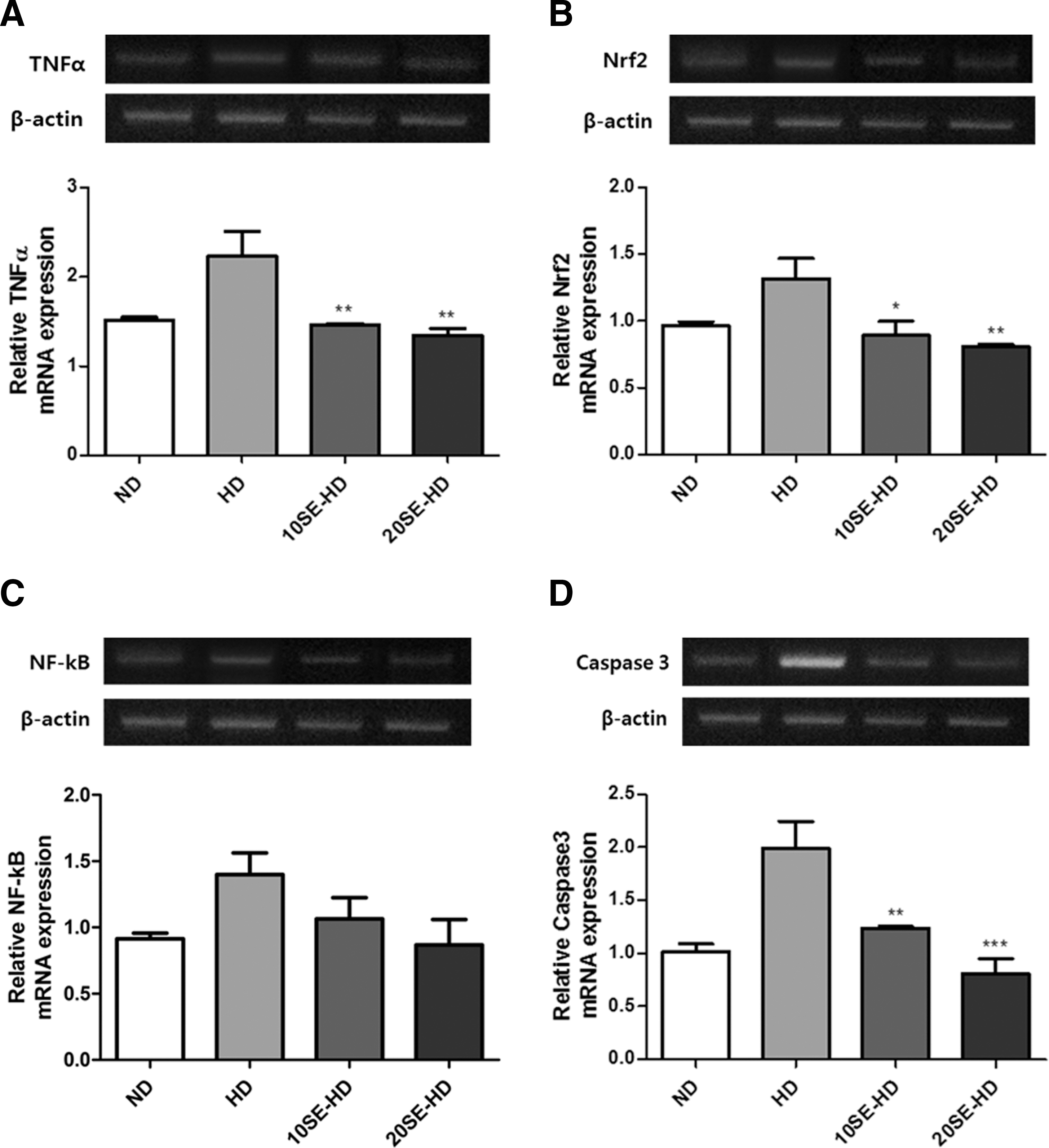
Oxidative stress-related mRNA expression.
TG, TC, FFA, HDL, and LDL/VLDL cholesterol
The TGs in the serum were significantly lower in the group fed 20SE-HD (P < .001) than that in the group fed HD. Serum TC showed a decrease in the mice fed 10SE-HD and 20SE-HD (P < .0001). Hepatic TC and TG were significantly lower in the mice fed 10SE-HD (P < .05 and P < .0001) and 20SE-HD (P < .0001) than in the mice fed HD. The HDL cholesterol in all experimental groups, except the HD group (P < .001 and P < .0001), was increased significantly. The LDL/VLDL cholesterol was not significantly changed in the experimental groups. Serum FFA in 10SE-HD and 20SE-HD (P < .001 and P < .0001) were lower compared to the HD group (Table 5).
Values are expressed as mean (SEM). ND (n = 7); HD (n = 7); 10SE-HD, HD +10% SE powder (n = 7); 20SE-HD, HD +20% SE powder (n = 7).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD, P < .05).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD, P < .01).
Significant differences are indicated by (HD vs. 10SE-HD/20SE-HD, P < .001).
HDL, high-density lipoprotein; LDL, low-density lipoprotein; VLDL, very low-density lipoprotein.
Adiponectin, AMPKα, HMG-CoA reductase, SREBP-1c, FAS, and Apo B mRNA expressions
Adiponectin expression in the experimental groups (P < .01 and P < .0001) was significantly higher compared to that of the HD group (Fig. 5A). However, AMPKα expression in the experimental group (P < .01) was significantly higher than in the HD group (Fig. 5B). Expression of 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase (P < .02) (Fig. 5C) and SREBP-1c (P < .001) (Fig. 5D) was significantly lower in the 20SE-HD group. In addition, expression of FAS (P < .001 and P < .0003) (Fig. 5E) and apolipoprotein B (Apo B) (P < .0001) (Fig. 5F) was significantly lower in the experimental groups, compared with those fed the HD.
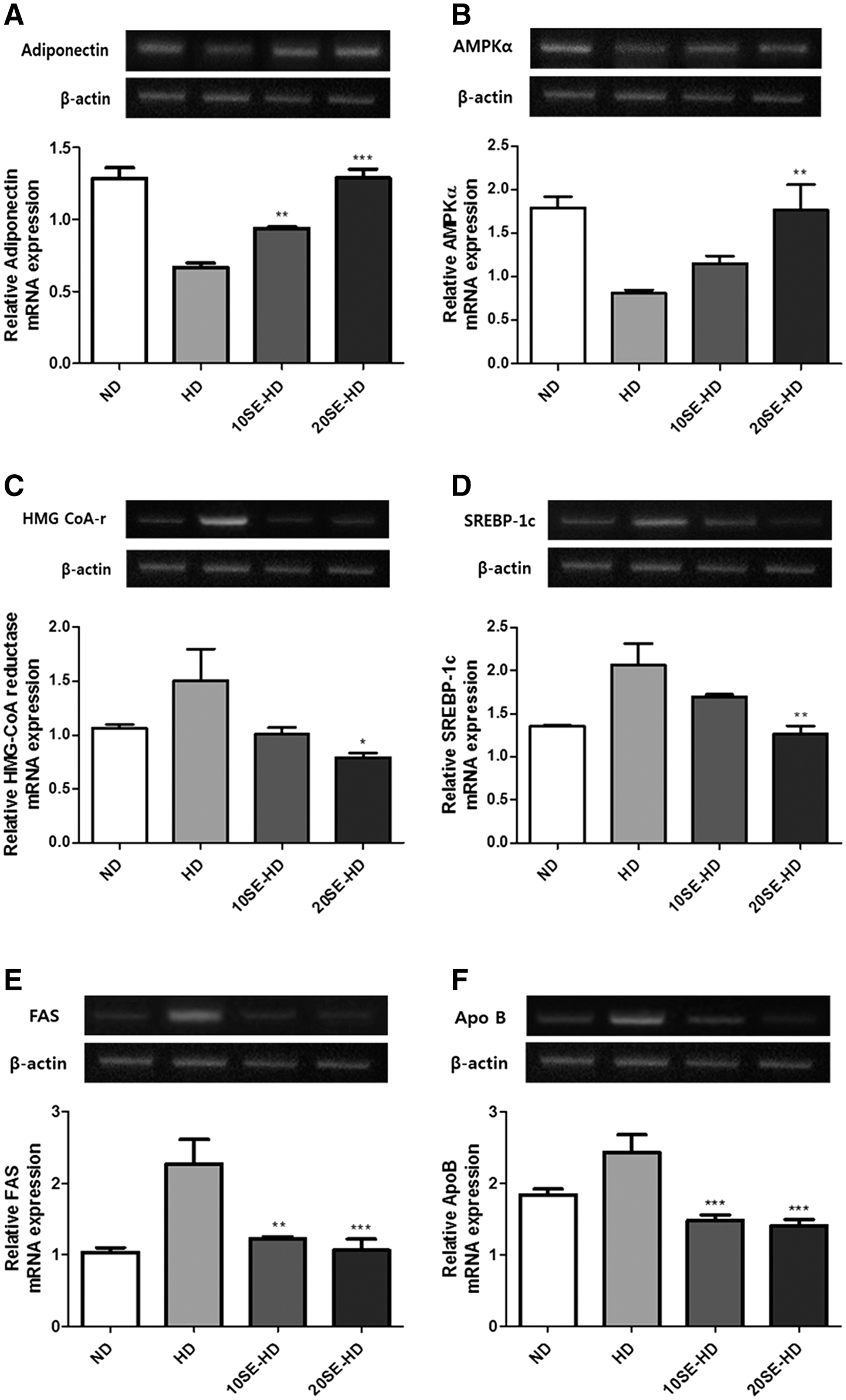
Lipid metabolism-related mRNA expression.
Discussion
One of the popular methods of inducing NAFLD in mice is a high-fat diet, which has been used for the development of diet-induced steatosis and NASH. Increasing the lipid content of experimental diets triggers metabolic diseases, such as obesity, hyperinsulinemia, hypercholesterolemia, and dyslipidemia. 4,5 Mice fed a high-fat diet show significant increases in body weight gain. 5 The results of this study are in agreement with previous findings of studies on mice fed high-fat diets. 2,44 The NAFLD is characterized by hepatic TG accumulation. It is caused by metabolic imbalance, which results from an increase in hepatic TGs and FFAs and a decrease in the release of TGs and FFAs from the liver. Involvement of FFAs in such cases is typically in the blood; some FFAs are provided from meal or hepatic fatty acid synthesis. 45 In our study, NAFLD-induced mice showed an identical trend to those used in previous studies in that a significant increase is seen in the liver weight of NAFLD-induced mice. 2,5 In addition, we suggest that consumption of soybean embryos inhibits hepatic TG accumulation, and high-dose soybean embryos were more effective than low-dose consumption of soybean embryos. We reached this conclusion based on the significantly lower liver weights of mice fed soybean embryos.
We further investigated the antioxidant effect of soybean embryos by measuring the antioxidant enzyme defense systems, such as SOD, catalase, and GPx. Diabetic mice fed genistein and daidzein showed enhanced activation of catalase and GPx. 46,47 These results indicated a similar trend to what was seen in the HD-induced mice fed tofu. A significant increase in hepatic SOD, catalase, and GPx occurred in hyperlipidemic rats that were fed bean sprouts containing rich isoflavones compared to the control group. Based on these results, isoflavones have been reported to improve lipid metabolism. Furthermore, isoflavones protect against oxidative stress and free radical damage by inhibiting oxidative damage and promotion of cell proliferation. 47,48 Our data and previous results showed similar findings, which was that isoflavone in soybean embryos decreased tissue damage in the liver during the NAFLD-induced period and interrupted cascades on the development of fatty liver disease. In addition, the inflammatory response in the liver is caused by the overexpression of NF-κB, which increases the expression of TNFα. 49 The expression of NF-κB is an important factor regulating pathogenesis of NAFLD by endoplasmic reticulum stress in the liver. 4 In this study, mice fed soybean embryos showed a decrease in TNFα and NF-κB gene expression compared to the NAFLD group. We concluded that consumption of soybean embryos has an anti-inflammatory effect in the liver. Nrf2, which has anti-inflammatory effects, is involved in the activation of immunomodulating processes and the effector function of the T cell and B cell. 50,51 We conclude that consumption of soybean embryos has an effect on the expression of Nrf2 through inhibition of the formation of reactive oxygen species in the liver. Previous observations in cases of fatty liver suggested that progression of fatty liver disease is caused by overexpression of apoptosis-stimulated protein, such as caspase-3 or fas-L. 52 The inhibitory effect of fatty liver disease demonstrated in the soybean embryo-fed group exhibited significantly decreased apoptosis-stimulated protein expression, compared to the HD-fed group. The HD-induced obesity and NAFLD is associated with hyperlipidemia, which is caused by increased TGs, LDL, and TC. 2,44,53 Previous studies have demonstrated that inhibition of lipid accumulation is caused by the activation of β-oxidation and downregulation of fatty acid synthesis through the consumption of soybeans. 44,54 In addition, the studies reported that TG, TC, and HDL cholesterol levels are changed by increases in cholesterol and synthesis of bile acid. 44 A study on improvement of fatty liver by chongkukjang consumption indicated that the improvement of lipid profiles is due to the suppressed LDL cholesterol oxidation and activation of lipoprotein lipase by biocompounds, such as phenols and flavonoids, in soybeans. 55 Furthermore, many studies demonstrated that isoflavones in soybeans have positive effects on cholesterol metabolism by decreasing cholesterol levels, protecting LDL cholesterol against oxidation, and enhancement of LDL cholesterol receptor activity. 56,57 Elevation of oxidative stress and stimulation of inflammatory responses due to an increase in FFAs can trigger liver injury, which is commonly associated with hepatic lipid accumulation. However, hepatic fat accumulation in obesity is caused by beta-oxidation and esterification through an increased inflow of FFA to the liver. 58,59 We observed a significant decrease in the lipid profile such as serum and hepatic TG, TC in serum and liver, serum LDL/VLDL cholesterol levels, serum HDL cholesterol, and serum FFAs, which are important indicators of NAFLD. In addition, we established that soybean embryos had a positive effect on lipid metabolism of NAFLD through the improvement of the lipid profiles.
The soybean embryo-fed groups showed significant improvements in lipid metabolism parameters, such as adiponectin, AMPK, SREBP-1c, and FAS. Based on these results, we postulate that consumption of soybean embryos improves hepatic lipid metabolism. When cholesterol levels increase, downregulation of HMG-CoA reductase has been shown to decrease hepatic cholesterol biosynthesis, which is caused by the continual activation of the SREBP2 and LDL receptor. These factors participate in cholesterol metabolism and act as regulators of the blood cholesterol level through stimulation of LDL cholesterol absorption. We evaluated the effect of cholesterol reduction through inhibition of HMG-CoA reductase in the soybean embryo-fed group compared with NAFLD-induced mice. Many studies have found that the ApoB/ApoA-I ratio is highly associated with metabolic syndrome and cardiovascular diseases. The effect of the soybean peptide on cholesterol lowering, following previous findings, is caused by increased excretion of cholesterol through downregulation of Apo B binding to the LDL receptor and accelerated clearance of LDL. The results for Apo B demonstrated that supplementation with soybean embryo can inhibit the progression of fatty liver disease. 60,61 This study was performed as an initial and fundamental one with whole soybean embryos. Therefore, as mentioned above, the levels of serum and hepatic lipid contents and mRNA expression were measured to prove any effect of soybean embryos on NAFLD. However, follow-up research measuring the levels of protein expression is needed.
We believe that this research represents a significant advance in the utilization of functional foods because it opens the very gate to utilizing whole soybean embryos as a supplementation, which is far different from such laboratory-produced extracts and supplements as used in most of the previous studies. The supplementation used in the article supports the use of soybean embryos for ameliorating NAFLDs because it comes directly from whole soybean embryos, which bear both isoflavone and soyasaponin, whereas the majority of previous studies were mainly divided into isoflavone-oriented and soyasaponin-focused ones.
We conclude that there is a protective effect of soybean embryos against NAFLD through the reduction of cholesterol and TG in serum and liver. The results of the study indicate that antioxidant and anti-inflammatory activities of soybean embryos significantly deter the progression of NASH by increased adipose tissues, and the regular supplementation of soybean embryos could improve adiponectin-mediated AMPKα pathway parameters, which are implicated in antioxidant, anti-inflammatory, lipid metabolism, and related NAFLD complications, such as obesity, type 2 diabetes, and metabolic syndrome.
Footnotes
Acknowledgment
This Research was supported by Sookmyung Women's University Research Grants 2014 (1-1403-0054).
Author Disclosure Statement
No competing financial interests exist.