
Abstract
The ancient Chinese wild rice (Zizania latifolia (Griseb) Turcz) (CWR) has valuable biological and medicinal functions. To assess the advantages lost in modern cultivated rice after domestication, we compared the composition of bioactive compounds and the results of proteomic analysis with those of Indica rice (N22). We used routine methods to determine the protein, total dietary fiber, amino acid, mineral substance, plant secondary metabolites, and amino acid composition of CWR and N22. The protein and mineral contents of CWR were two times that of N22, and the levels of calcium, potassium, magnesium, chromium, iron, and zinc were significantly higher than those of N22 (P < .05). There was ∼7.6 times more dietary fiber in CWR than in N22, but fewer carbohydrates (P < .05). Anthocyanins and chlorophyll were detected in CWR, but were absent from N22. Compared with N22, CWR had 53, 19, and 5.4 times higher (P < .05) levels of saponins, flavonoids, and plant sterols, respectively. The amino acid score of CWR was 66.6, which was significantly higher than N22. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) indicated that the main seed proteins of CWR were glutelins, including both acid and alkaline subunits, which were approximately twice those of N22. To investigate the differences in protein profiles between CWR and N22, we conducted two-dimensional electrophoresis (2-DE) analysis of the total proteins in the seeds of the two rice species. 2-DE gels revealed 19 differentially expressed proteins. Information obtained from peptide mass fingerprinting indicates that glutelin precursor caffeoyl coenzyme A (CoA) O-methyltransferase and putative bithoraxoid-like protein can provide good gene sources for improving rice quality.
Introduction
C
Materials and Methods
Sampling of CWR and N22
CWR was collected in the harvest year from Bao-ying Lake, Jiangsu province, China. N22 was provided by the Rice Research Institute of Nanjing Agricultural University, Nanjing, China (Fig. 1). The samples were sun-dried, hulled by hand, and stored at room temperature (18–23°C) before use.

Seeds of CWR and N22. The left is CWR and the right is N22. CWR, Chinese wild rice. Color images available online at
Analysis of nutrient content, plant secondary metabolites, and amino acid composition
Protein content (nitrogen × 6.25) was determined by the Kjeldahl method, 8 and the direct oven method was used for the determination of moisture at 130°C for 1 h. 8 To measure total fat, the Soxhlet extraction method was used with ether as a solvent. 8 Mineral composition was determined gravimetrically, 8 and although it is now a nutritionally obsolete method, the total dietary fiber, soluble fiber, and insoluble fiber were determined by the enzymatic–gravimetric method according to the People's Republic of China National Standards (PRCNS) (GB/T 5009.88-2008) 9 to enable comparisons with earlier data. The critical steps of these analyses are described in the methods of the AOAC (1984).
Mineral analysis
K, Mg, Ca, Na, Fe, Zn, Cu, Mn, and Cr were detected using Atomic Absorption Spectroscopy (SP-3500AA; Shanghai). Phosphorus content was analyzed by the molybdenum blue method (PRCNS, GB/T 5009.87-2003). 10 All measurements were conducted in duplicate and the means and standard deviations calculated. Only duplicates with standard deviations of <5% were accepted. USBS cattle liver and orange leaf (Nos. SRM 1577 and SRM 1572) were used as standards.
Measurement of plant secondary metabolites
Chlorophyll was measured by a spectrophotometric method (PRCNS, GB/T22182-2008) 11 after the samples were milled and extracted. Lipid components in CWR and N22 were determined by Soxhlet extraction, followed by phytosterol analysis by gas chromatography (GC) 7809A (Aglient Technologies). Flavonoid, saponin, and anthocyanins were analyzed using colorimetry.
Amino acid composition analysis
The grain samples were hydrolyzed in HCl, and 17 amino acids were measured with an automated amino acid analyzer (Hitachi model 835-50). Tryptophan, grain samples of Tryptophan, was measured by hydrolyzing samples in NaOH and determining tryptophan content was using a Hitachi spectrofluorometer (model 850-2). 12
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was performed following the method described by using a 15% (w/w) separating gel and a 7.5% (w/w) stacking gel. 13 Each flour seed was extracted using 1 mL of extraction buffer [50 M Tris/HCl pH 6.8, 8 M Urea, 4% (w/w) SDS, 20% (w/w) glycerol, and 0.004% bromophenol blue] with 0.01 M β-mercaptoethanol. The mixture was agitated on a rotary shaker for 30 seconds at room temperature and centrifuged at 4800 g for 5 minutes. The supernatant containing the soluble protein fraction was used to prepare samples for SDS-PAGE. Supernatant protein concentration was quantified using the Lowry method, 14 and proteins were stained with Coomassie Brilliant Blue G-250 (CBB G-250) (Amersham Biosciences).
Two-dimensional gel electrophoresis
Seed proteins were extracted using a modified trichloroacetic acid/acetone procedure as described previously. 15 To remove pigments and lipids, the precipitated proteins were washed with ice-cold acetone containing 0.07% dithiothreitol (DTT), until the supernatant was colorless. The pellet was vacuum-dried, resuspended in solubilization solution, 7 M urea, 2 M thiourea, 4% [(3-Cholamidopropyl) dimethylammonio] propanesulfonic acid (CHAPS), 65 mM DTT, 0.5% Bio-Lyte 3/10 ampholyte, and 0.1% phenylmethanesulfonyl fluoride (PMSF), and sonicated to extract the proteins. The protein concentration of the final supernatant was measured according to the Bradford method, 16 using bovine serum albumin as the standard.
2D-PAGE analysis of rice glutelin was performed according to previously described methods 17 with minor modifications. The prepared samples were initially analyzed by isoelectric focusing using pH3-10; 17-cm strips of polyacrylamide gel containing 7 M urea, 2 M thiourea, 4% CHAPS, 10 mM DTT, 0.25% Bio-Lyte 3/10 ampholyte, and 0.1% PMSF. Isoelectric focusing was performed for 0.5 hours at 200 V, 0.5 hours at 500 V, 1 hour at 1000 V, 1 hour at 2000 V, and 5 hours at 8000 V and finally held constant at 8000 V until a total of at least 55,000 V was reached. The strips were then loaded onto 12.5% polyacrylamide gels for SDS-PAGE analysis. The gels were stained with CBB, R-250.
In-gel digestion of proteins
Protein spots were excised, dehydrated in acetonitrile, and dried at room temperature. Dried gel pieces were reswollen with 2–3 μL of trypsin (Promega) solution (10 ng/μL in 25 mM ammonium bicarbonate) at 4°C for 30 minutes. All samples were allowed to air dry at room temperature, and 0.1% trifluoroacetic acid (TFA) was used for on-target washing.
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry
Matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry (MS) and tandem TOF/TOF MS were performed on a time-of-flight Autolex III mass spectrometer (Bruker Daltonics). Peptide mass maps were acquired in positive reflection mode, averaging 400 laser shots per MALDI-TOF spectrum (resolution was 15,000–20,000). Bruker calibration mixtures were used to calibrate spectra to a mass tolerance within 0.1 Da.
Database query
Database searching and peptide mass fingerprinting were performed using MASCOT (
Statistical analyses
Data were analyzed by Student's t-test using SPSS for Windows (v. 19.0; SPSS, Inc.). In all analyses, the level of significance was set at P < .05.
Results
CWR had a higher content of all nutrients except carbohydrate than N22
The protein and mineral contents of CWR were two times that of N22, and the levels of calcium, potassium, magnesium, chromium, iron, and zinc were significantly higher than those of N22 (P < .05, Table 1). There was ∼7.6 times more dietary fiber in CWR than in N22, but fewer carbohydrates (P < .05, Table 1). Anthocyanins and chlorophyll were detected in CWR, but were absent from N22. Compared with N22, CWR had 53, 19, and 5.4 times higher (P < .05, Table 1) levels of saponins, flavonoids, and plant sterols, respectively.
Mean values within a row with * were significantly different by Student's t-test for five samples (P < .05).
CWR, Chinese wild rice.
In our study, CWR had higher levels of protein, fat, total dietary fiber, magnesium, phosphorus, potassium, sodium, iron, and manganese, but the total carbohydrate content in CWR was lower than in N22 (P < .05).
CWR had greater and a more diverse amino acid composition than N22
CWR contained higher levels of all amino acids (Fig. 2). The amino acid score of CWR was 66.6, which was significantly higher than 51.1 for N22 (Table 2). The percentage of essential amino acids was 45% in CWR and 50% in N22. The total amounts of essential amino acids in CWR and N22 were 5.21 mg/100 g and 2.98 mg/100 g, respectively.

The amino acid content of CWR and N22. Data are expressed as the mean ± SD for five samples. Bars with * indicate significant differences between the two groups at P < .05.
The second-limited amino acids of CWR.
The first-limited amino acids of CWR.
The first-limited amino acids of N22.
The second-limited amino acids of N22.
AAS, amino acid score; EAA, essential amino acids; NEAA, nonessential amino acids; CWRAA, Chinese wild rice amino acid.
CWR has higher glutelin content than N22
SDS-PAGE indicated that the main seed protein of CWR was glutelin, including both acid (37–39 kDa) and alkaline (22–23 kDa) subunits. Its content was approximately two times that of N22 (Fig. 3).
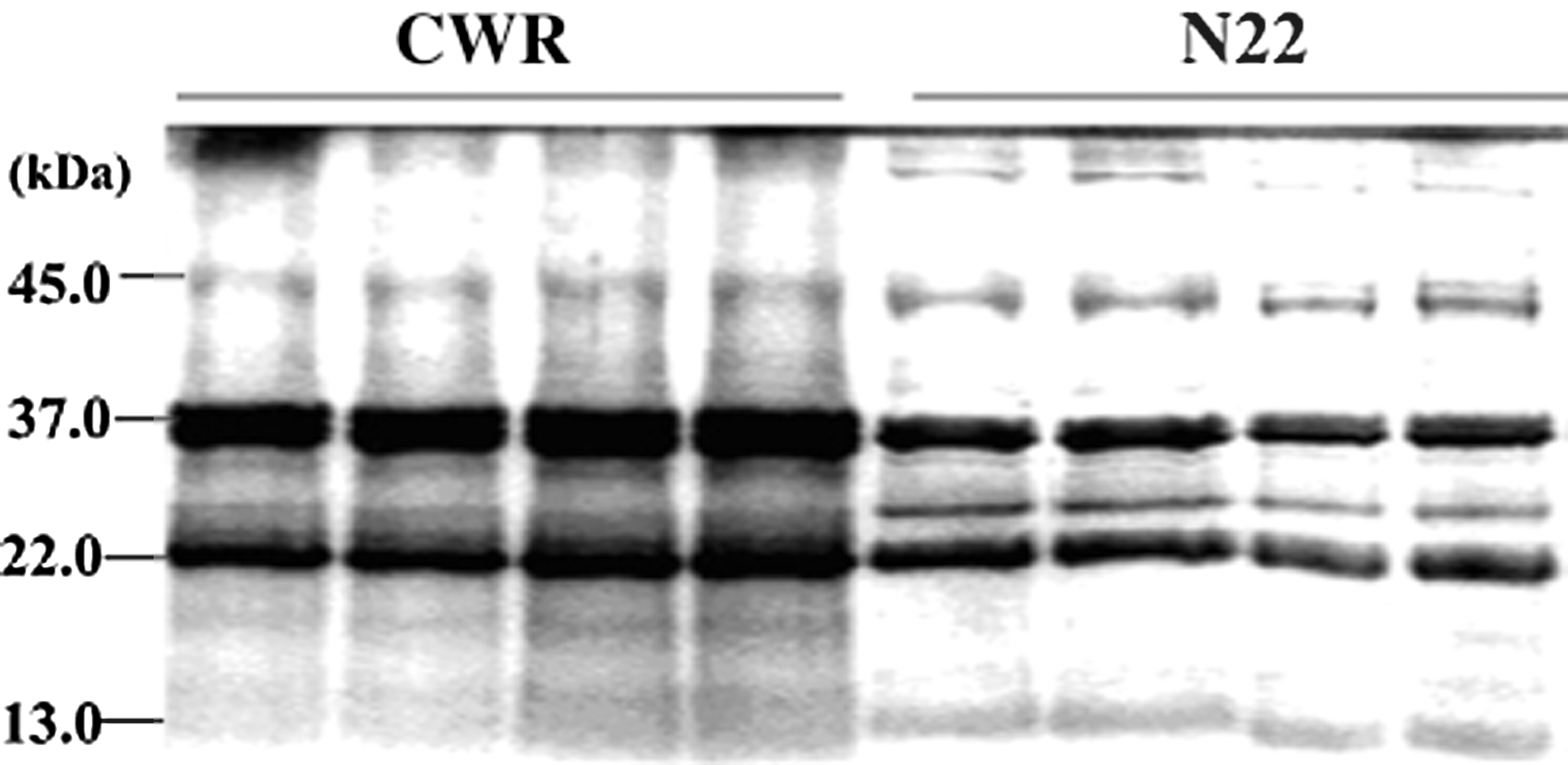
The SDS-PAGE gel of CWR and N22 seed proteins. SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
Differences in protein composition between CWR and N22
To investigate the differences in protein profiles between CWR and N22, we conducted two-dimensional electrophoresis (2-DE) analysis of the total proteins in the seeds of CWR and N22 using pH 3-10NL 17-cm IPG strips with the CBB G-250 staining method. For each sample, the gels were repeated in triplicate and showed a high level of reproducibility. Representative gels are shown in Figure 4. Using PDQest software version 8.0 (Bio-Rad), a total of 68 spots were obtained in the CWR sample and 123 spots in N22. Most of the proteins had molecular weights between 35 and 60 kDa. In each sample, albumins and globulins were mostly distributed in the pH range of six to nine.

Representative Coomassie Brilliant Blue-stained 2D-PAGE gel of separated CWR and N22 proteins. The experiments were totally repeated for three times with three repetitions for each group.
2-DE gels resulted in 19 differentially expressed proteins (Fig. 4). When the predicted sequences of the tryptic peptides from the 19 differently expressed proteins were referenced with the NCBI database, 16 spots were identified, while three spots (1, 3, and 15) remained unidentified (Table 3). Spot 14 results are reported in Figure 5 as an example.
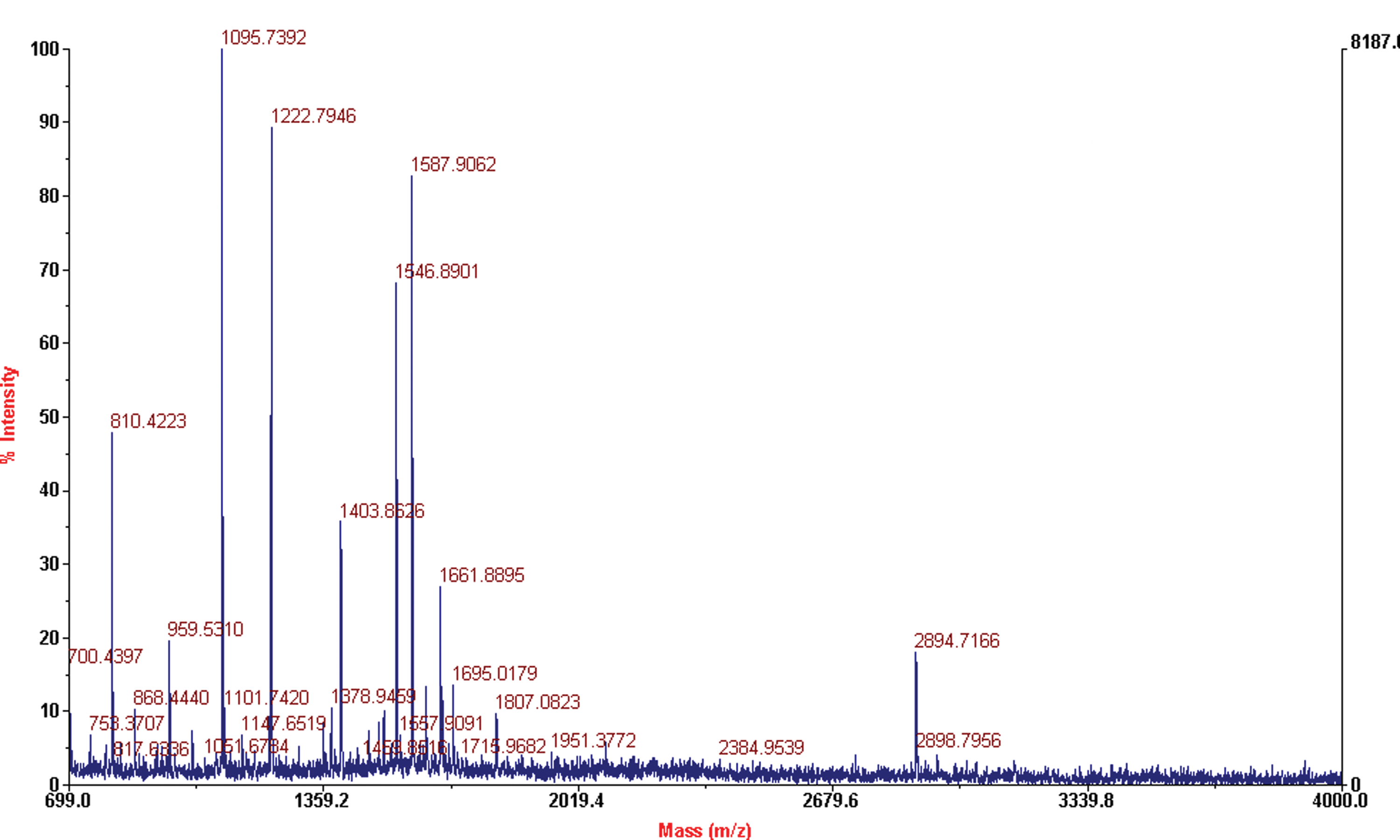
Spot 14 identified as caffeoyl-CoA O-methyltransferase by peptide mass fingerprinting. Color images available online at
Acc. No, Accession number; NM/PI, nominal mass and isoelectric point; PC, protein score; SC, the percentage of sequence coverage; SID, spot ID.
Discussion
CWR has a long history as a food component and it is recorded in the annals of all dynasties in China. As early as 3000 years ago in the Zhou dynasty (11th B.C.), it had become a tribute food eaten by the emperor. 1,2 It was used as a herbal medicine to treat diabetes and regulate gastrointestinal function. 5 This long historical record suggests that the ancient Chinese people had already recognized the healthy benefits of CWR for the body. The results of the current study further improved our previous research data and hypothesis. We showed that proteins, minerals, and dietary fiber contents of CWR were significantly higher than those of N22 rice (Table 1). In our study, two types of active substances (anthocyanins and chlorophyll) were found in CWR, but were not detectable in N22. There was a much higher frequency of saponins, flavonoids, and phytosterols in CWR than in N22 (Table 1). The appearance of CWR was remarkably different from that of the modern cultivated Indica rice, in that the surface of CWR was covered with green or brown material, while the surface of N22 was ivory (Fig. 1). However, it is currently unknown whether the aforementioned anthocyanins, chlorophyll, saponins, flavonoids, and plant sterols derive from this surface layer. Our previous study indicated that CWR could improve the serum lipid profile and antioxidant status of rats fed a high-fat/cholesterol diet as well as improve glucose metabolism and insulin sensitivity. 3,7 These beneficial biological functions of CWR may be associated with the pigmented surface material.
The amino acid composition of food proteins provides important information and perspective about their nutritive quality. 18 The first-limiting amino acid of both CWR and N22 is Lysine, but the chemical score of CWR was 66, which was substantially higher compared with N22. In addition to the dietary requirement for protein, there are also requirements for nonessential amino acids, nitrogen, and essential amino acids. Consequently, proteins differ in nutritional value to the extent that their individual essential amino acid profile satisfies the body's requirements. The ratio between essential and nonessential amino acids in the diet is also important. The ratio of CWR approaches 1:1, the ratio recommended for optimal retention and utilization. 19 In conclusion, CWR has higher protein quality than N22.
The results of SDS-PAGE also showed that CWR has superior nutritional value. The content of glutelins in CWR was approximately two times that of N22. As the major storage protein in rice, 20 a high content (∼75% of total proteins on a weight basis) of glutelins in the starchy endosperm differentiates CWR from other cereal crops, with the exception of oat. Compared with prolamins, glutelins are more easily digestible and contain a higher amount of the essential amino acid lysine. 21 In general, the quality and quantity of glutelins determine the nutritional value of rice. In addition, our data showed that CWR has a very good amino acid composition, which further explains our previous finding that CWR has the high Protein Efficiency Ratio (PER) value of 2.75, 22 which was much higher compared with N22.
Using the NCBI database, we identified nine spots of CWR as rice proteins. This meant that the homology between CWR and N22 is 64%, but significant differences also exist between the two. We gained new insights by analyzing the biological function of the high reliability (credibility score 67 or higher, P < .05) peptide mass fingerprint of CWR.
Spots 4, 5, and 9 were identified as glutelin (Oryza sativa [japonica cultivar-group]), a major storage protein in rice endosperm cells that is transported to a protein storage vacuole. 20
Spots 6, 7, and 12 were identified as a glutelin precursor [Z. latifolia], which is synthesized by glutelin in the endosperm endoplasmic reticulum and then deposited within the protein storage vacuole protein body-II (PB-II) as an aggregate, with a high degree of polymerized higher order structure comprising mature acidic and basic subunits after post-translation processing cleavage. 23 Additionally, two full-length cDNA sequences of CWR glutelin were cloned and added in the Genbank database (DQ665811.1 and DQ659137.1), 24,25 thereby providing new genetic resources for applying the transfer gene technique to rice, which could be advantageous for improving rice nourishment quality.
Spot 14 was identified as caffeoyl coenzyme A (CoA) O-methyltransferase (CoAOMT). It has been found that CCoAOMT is essential for lignin biosynthesis. 26 Lignin provides the mechanical strength to the walls of conducting tracheary elements (TEs), which is necessary for enduring the negative pressure generated from transpiration. It also renders the walls of maturing TEs indigestible by the hydrolytic enzymes released during autolysis of xylogenesis. In addition, as an insoluble dietary fiber, lignin with high water retention capacity and water swelling capacity enhances satiety, increases fecal volume, and shortens defecation time, thereby reducing the risk of obesity, constipation, diabetes, and colon cancer. 27 The protein expression provides the substance evidence in accordance with biological characteristics of CWR, such as preventing insulin resistance.
Interestingly, Spot 16 was identified as a putative bithoraxoid-like protein [Spermatozopsis similis]. Bithoraxoid (Bxd) is a low-molecular-weight late transcript of the Bxd region of several functional domains, such as ultrabithorax domain. Although Bxd-like proteins appear to be widespread in animals, for example, in Caenorhabditis elegans and in different mammals (including humans), no function had been attributed to the Bxd-like proteins until recently. Similar proteins from higher plants have not yet been identified. 28 It may prove that CWR is an ancient plant and that the plant and animal cells have the same mechanism for the transmission of energy and information.
Spot17 was identified as Os06g0604500 (O. sativa [japonica cultivar-group]), belonging to Ca2+-binding proteins. In plants, intracellular calcium levels are modulated in response to various signals, including abiotic stresses, light, pathogens, hormones, and nutrition. Nevertheless, an additional level of regulation in calcium signaling is achieved through the function of calcium-binding proteins. 29 The sensor proteins of CWR likely recognize specific calcium signatures and relay these signals into downstream responses, such as phosphorylation cascades and regulation of gene expression, and also show special functions different from N22.
In summary, we found that compared with CWR, the modern cultivated N22 obtained some new biological characteristics after domestication, such as synchronized maturation and high yield and ease of harvest. Meanwhile, some biologically active components were also lost in the process of evolution, including proteins, minerals, and dietary fiber, especially anthocyanin, chlorophyll, and phytochemicals, such as saponins, flavonoids, and plant sterols. In addition, the nutrients and phytochemical components play an important role in the mechanism underlying the protective effect of CWR against obesity and lipotoxicity as we observed recently. The identification of differentially expressed proteins between CWR and N22, such as Spots 14, 16, and 5, also provides an excellent resource for improving the quality of cultivated rice, which requires further study.
The current study was limited in the number of proteins analyzed as well as in the use of quantitative techniques, such as western blotting. Therefore, further research on these proteins is essential.
Footnotes
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (Nos. 30471451 and 81372986). C.-K.Z. designed the study and interpreted the data. M.X.J. and H.Y. conducted the study. M.-X.J. collated the data, performed all of the statistical analyses, interpreted the data, and wrote the manuscript. L.-J.Z. and S.-M.Z. interpreted the data and corrected the English. All authors read and approved the findings of the study.
Author Disclosure Statement
No competing financial interests exist.