
Abstract
Nonalcoholic fatty liver disease (NAFLD) is a common chronic liver disease that is rising in prevalence worldwide. Therapeutic strategies for patients with NAFLD are limited by a lack of effective drugs. In this report, we show that Dolichos lablab water extract (DLL-Ex) protects against free fatty acid (FFA)-induced lipid accumulation and attenuates expression of genes involved in lipid droplet accumulation in cellular NAFLD models. The hepatoprotective effects and underlying mechanism of DLL-Ex were assessed using an in vitro cellular model in which NAFLD was simulated by inducing excessive FFA influx into hepatocytes. HepG2 cells were treated with DLL-Ex and FFAs for 24 h, after which intracellular lipid content was observed by using Nile Red and Oil Red O staining. Quantitative real-time polymerase chain reaction was used to measure expression levels of genes related to FFA-mediated cellular energy depletion. Western blotting was used to measure protein levels of phosphorylated c-Jun N-terminal kinase, AMP-activated protein kinase alpha (AMPKα), and peroxisome proliferator-activated receptor γ coactivator 1 alpha. In HepG2 cells, DLL-Ex inhibited expression of CD36, which regulates fatty acid uptake, as well as BODIPY-labeled fatty acid uptake. Additionally, DLL-Ex significantly attenuated FFA-mediated cellular energy depletion and mitochondrial membrane depolarization. Furthermore, DLL-Ex enhanced phosphorylation of AMPK, indicating that AMPK is a critical regulator of DLL-Ex-mediated inhibition of hepatic lipid accumulation, possibly through its antioxidative effect. These results demonstrate that DLL-Ex exerts potent anti-NAFLD activity, suggesting that it could be a potential adjuvant treatment for patients with NAFLD.
Introduction
N
Although the pathogenesis of NAFLD remains unclear, it is characterized by the accumulation of lipids, primarily triglycerides (TGs), in hepatocytes, as well as damage to liver cells, which is referred to as hepatic steatosis. 5,6 Increased delivery and uptake of free fatty acids (FFAs) in hepatocytes due to excess dietary intake or release from adipose tissue, increased de novo hepatic FFA and TG synthesis, failure of very low-density lipoprotein synthesis and TG export, or failure of FFA elimination due to impaired hepatic mitochondrial β-oxidation may induce NAFLD. 7 –9 Therefore, therapies that inhibit hepatic lipid accumulation may be useful for controlling NAFLD. NAFLD treatments primarily target the main components of metabolic syndrome, including obesity, diabetes, hypertension, and dyslipidemia. 10 Although numerous drugs are currently employed to treat NAFLD, no specific therapy is available. 11
Traditional medicines, particularly herbal products, have been widely used to treat liver diseases. In recent years, the frequency of studies utilizing these alternative approaches has increased with the popularity of traditional medicine as an alternative to Western medicine. 12,13 Herbal products with multiple components have diverse effects because of the synergistic activities of their constituent herbs, which may prevent adverse effects. Dolichos lablab Linne, commonly referred to as field bean or hyacinth bean, is a legume that produces a pulse that is consumed by humans and used as livestock forage. 14 D. lablab has been used to improve digestive absorption in patients with chronic diseases and is an effective therapy for patients with diarrhea.
The purpose of this study was to elucidate the hepatoprotective effects and underlying mechanism of D. lablab water extract (DLL-Ex) in an in vitro cellular NAFLD model. The results of the present study indicate that DLL-Ex inhibits cellular lipid accumulation, downregulates mRNA expression of fatty acid synthesis-associated genes, and significantly reduces hepatic FFA uptake by inhibiting fatty acid translocase (FAT and CD36) expression. Moreover, inhibition of FFA signaling by DLL-Ex was mediated by AMP-activated protein kinase (AMPK) activation.
Materials and Methods
Preparation of DLL-Ex
Dried D. lablab was purchased from Naemome DAH Co. (Ulsan, Korea). To prepare DLL-Ex, dried D. lablab (200 g) was ground into a fine powder and extracted with 2 L of water at 90–100°C for 2 h. The crude extract solution was filtered, evaporated, and lyophilized in a freeze dryer. The dry weight of the DLL-Ex was 32.3 g (16.12% w/w yield). DLL-Ex was stored at the Korea Institute of Oriental Medicine (Daejeon, Korea) and dissolved in distilled phosphate buffered saline (DPBS) before its application in cells.
Cell culture and treatment
HepG2 cells were obtained from the American Type Culture Collection (Manassas, VA, USA) and maintained in a 1:1 mixture of Dulbecco's modified Eagle's medium (DMEM) and F-12 (50:50; Invitrogen, Carlsbad, CA, USA) supplemented with 1% penicillin/streptomycin (P/S; Gibco, Grand Island, NY, USA) and 10% heat-inactivated fetal bovine serum (Invitrogen) at 37°C in an atmosphere containing 5% CO2. Oleic acid (OA) and palmitic acid (PA) were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA) and prepared in 0.1 M NaOH at 70°C, after which they were filter sterilized using a syringe filter (Sartorius Stedim Biotech, Goettingen, Germany). An FFA (0.66 mM OA and 0.33 mM PA) solution was prepared at a concentration of 1 mM with bovine serum albumin (BSA) and used at a final concentration of 1% in the culture medium. Cells were starved in DMEM and F-12 (50:50; Invitrogen) supplemented with 1% P/S (Gibco-BRL, Grand Island, NY, USA) and 0.5% heat-inactivated fetal bovine serum (Invitrogen) at 37°C in an atmosphere containing 5% CO2. Cells were used when they reached 75% confluence.
Cell viability assay
DLL-Ex or FFA cytotoxicity was assessed using the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium inner salt (MTS) assay as described previously. 15 HepG2 cells in 96-well plates were treated with various concentrations of DLL-Ex or FFA. After incubation for 24 h, MTS solution (CellTiter Aqueous One Solution; Promega, Madison, WI, USA) was added to each well, after which the plates were incubated at 37°C for 4 h. The optical absorbance of the solution in each well was determined at 490 nm with a microplate spectrophotometer (Molecular Devices, Inc., Sunnyvale, CA, USA). Each treatment was performed in triplicate.
Nile Red and Oil Red O staining
Nile Red, a fluorescent hydrophobic dye, was applied to assess the presence of phospholipids in HepG2 cells. 16,17 Intracellular fat content was determined through staining with Oil Red O, a lipophilic dye, used to label fat accumulation in the cytosol. After 24 h of FFA exposure with or without DLL-Ex, cells were washed twice with PBS and incubated with 0.75 μg/mL Nile Red dye for 15 min at room temperature. To minimize dye photobleaching, the plates were wrapped in aluminum foil. The fluorescence intensity of the solution was measured with a fluorescence spectrophotometer (excitation: 485 nm; emission: 572 nm; SpectraMax M2, Molecular Devices, Inc.). For the Oil Red O staining, cells were exposed to FFAs for 24 h with or without DLL-Ex, washed twice with PBS, and fixed with 10% formalin for 1 h. Cells were stained with Oil Red O solution and examined under a light microscope. After observing the lipid droplets, 100% isopropanol was added to each well, after which the fluorescence intensity of the solution at 520 nm was measured with a spectrophotometer (Molecular Devices, Inc.). Each treatment was performed in triplicate.
Measurement of intracellular TG content
Cellular TG content was measured enzymatically with a commercial kit (FUJI DRI-CHEM, 3500; Fujifilm, Tokyo, Japan) following the manufacturer's instructions. After 24 h of FFA exposure with or without DLL-Ex, cells were washed twice with PBS, collected with trypsinization, and homogenized in 500 μL of 5% Triton X-100 in PBS with a homogenizer. Total cell extracts were incubated at 80°C for 5 min and cooled to room temperature. After further centrifugation, the supernatant of each sample was collected and used for TG measurement. TG content was expressed in μg TGs per μg protein. Each treatment was performed in triplicate.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was isolated using TRIzol (Invitrogen) according to the manufacturer's instructions. Reverse transcription and quantitative real-time polymerase chain reaction (Q-PCR) were performed to detect relative mRNA expression using TaqMan probes (Applied Biosystems, Foster City, CA, USA) as described previously. 18 Relative amounts of cDNA were calculated by the relative quantification (ΔΔCt) method. Each sample was run in triplicate. The relative abundance of each transcript was normalized to that of ubiquitin C.
Measurement of fatty acid uptake
The effect of DLL-Ex on fatty acid uptake was examined using BODIPY-labeled fatty acid (C16; Invitrogen). HepG2 cells were incubated with 1 μM BODIPY-labeled fatty acid (C16), in the presence or absence of DLL-Ex at 37°C for 30 min, after which the cells were washed at 4°C with PBS containing 0.2% BSA to remove surface-associated BODIPY. BODIPY fluorescence was measured using a SpectraMax i3 fluorescence plate reader (Molecular Devices, Inc.).
ATP measurement
Cells were cultured in 96-well white plates. The total cellular ATP content was determined using an ATP Luminescence Detection Kit (PerkinElmer, Waltham, MA, USA) and an internal standard according to the manufacturer's instructions.
Mitochondrial membrane potential
The JC-1 Mitochondrial Membrane Potential Detection Kit (Biotium, Hayward, CA, USA) was used to measure the mitochondrial membrane potential (MMP) of HepG2 cells. Cells were seeded (1 × 104 cells/well) in 96-well white plates and incubated with 1× JC-1 staining solution at 37°C for 20 min, after which they were rinsed with PBS. Red fluorescence (excitation, 550 nm; emission, 600 nm) and green fluorescence (excitation, 485 nm; emission, 535 nm) were measured using a SoftMax Pro 5 fluorescence plate reader (Molecular Devices, Inc.). The ratio of green: red fluorescence, an index of mitochondrial membrane depolarization, was increased in dead cells and in cells undergoing apoptosis.
Western blotting analysis
HepG2 cells were lysed and total protein was extracted. Protein samples (20 μg) were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes, which were blocked for 1 h in 5% skim milk at room temperature. The membranes were incubated overnight at 4°C with specific antibodies for the genes of interest and β-actin (Cell Signaling Technology, Danvers, MA, USA). The blots were washed three times for 10 min each with PBS-T, after which the membranes were incubated for 2 h with anti-rabbit secondary antibodies (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The immunoblots were visualized using an ECL chemiluminescence detection system (GE Healthcare, Buckinghamshire, United Kingdom) and the bands were visualized using a chemiluminescence imaging system (ImageQuant LAS 4000 mini; GE Healthcare). Total ERK, JNK, p38, and AMPK, as well as β-actin expression, which was used as the loading control, were detected. The resulting image was acquired and the intensity of individual bands was quantified using the ImageJ (Institutes of Health, New York, NY, USA) densitometry software.
Statistical analysis
All experimental data are presented as the mean ± SEM. Multiple groups were tested by one-way analysis of variance followed by Dunnett's test to determine significant differences versus the control group. A P value less than .05 was considered to be significant (*,# P < .05 and **,## P < .001).
Results
Effect of DLL-Ex on FFA-induced lipid accumulation in HepG2 cells
An FFA-induced in vitro cellular hepatic steatosis model was used to investigate the efficacy of DLL-Ex against hepatic steatosis. FFA treatment for 24 h induced dramatic intracellular lipid accumulation in HepG2 cells (Fig. 1a, b). FFA had no apparent cytotoxic effect at concentrations up to 1 mM (Fig. 1c).
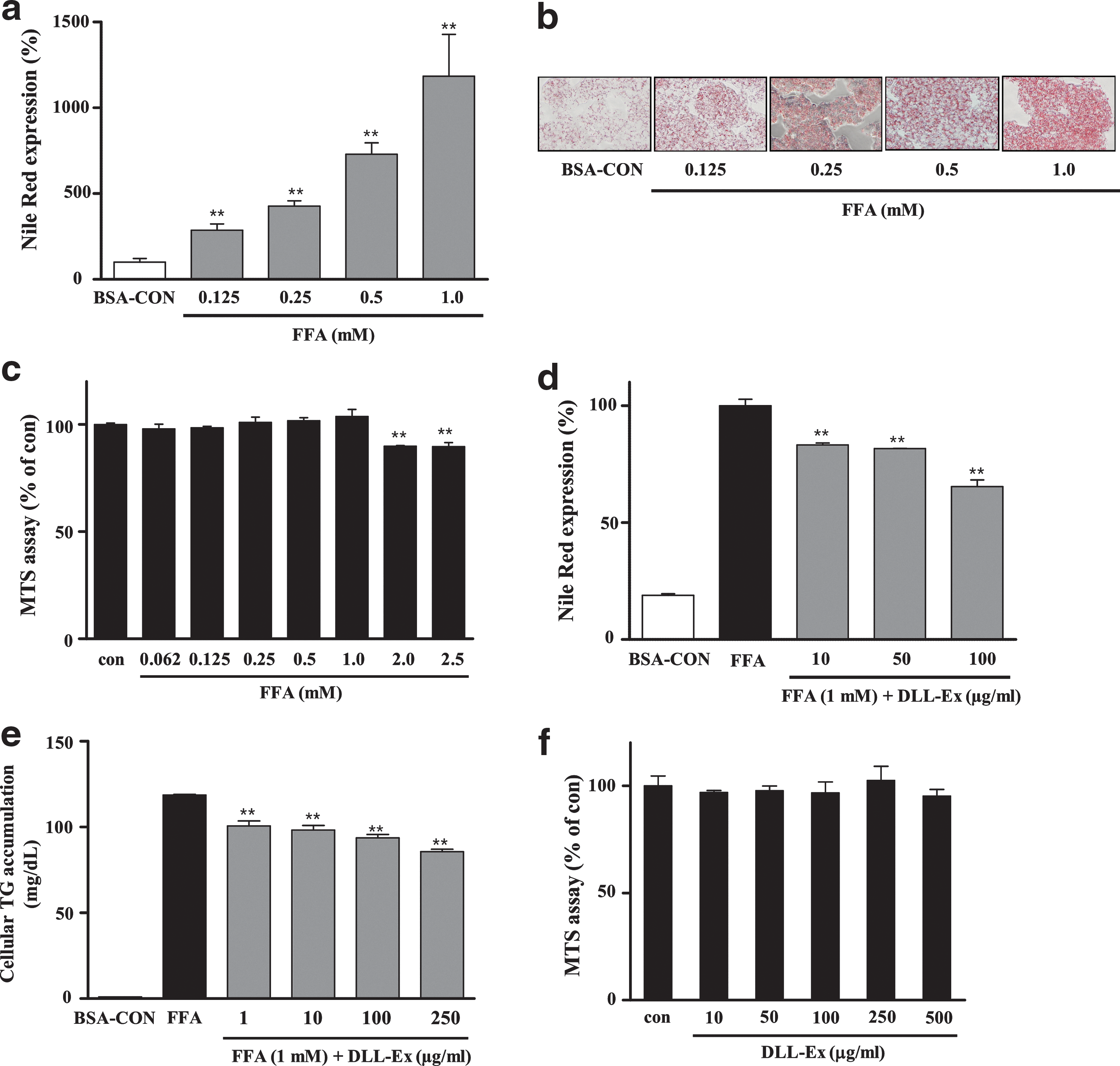
Inhibition of free fatty acid (FFA)-induced lipid accumulation by Dolichos lablab water extract (DLL-Ex) in HepG2 cells. FFA treatment for 24 h induced dramatic intracellular lipid accumulation in HepG2 cells
Nile Red staining indicated that treatment with DLL-Ex (10, 50, and 100 μg/mL) dose-dependently suppressed intracellular lipid accumulation (Fig. 1d). DLL-Ex (1, 10, 100, and 250 μg/mL)-mediated inhibition of FFA-induced lipid accumulation in HepG2 cells was assessed by quantification of intracellular TGs (Fig. 1e). DLL-Ex at concentrations less than 500 μg/mL did not significantly affect HepG2 cell viability, indicating that DLL-EX-mediated inhibition of intracellular lipid accumulation did not result from cytotoxicity (Fig. 1f).
Effects of DLL-Ex on lipid droplet accumulation and fatty acid uptake
Q-PCR analysis of lipid-associated pathway-specific genes showed that expression levels of genes involved in hepatic lipid uptake and lipid droplet accumulation were significantly reduced in DLL-Ex-treated cells in comparison with their expression levels in FFA-treated cells. Expression of adipose differentiation-related protein (ADRP, perilipin2, Plin2) was dramatically increased by FFA treatment and decreased by DLL-Ex treatment (1, 10, 100, and 250 μg/mL) (Fig. 2a). The expression of fat-specific protein 27 (Fsp27, Cidec) and fat-inducing transcript 2 (Fit2, fat storage-inducing transmembrane protein 2, Fitm2), which are involved in lipid droplet accumulation, was increased by FFA treatment and decreased by DLL-Ex (100 and 250 μg/mL) (Fig. 2b, c).
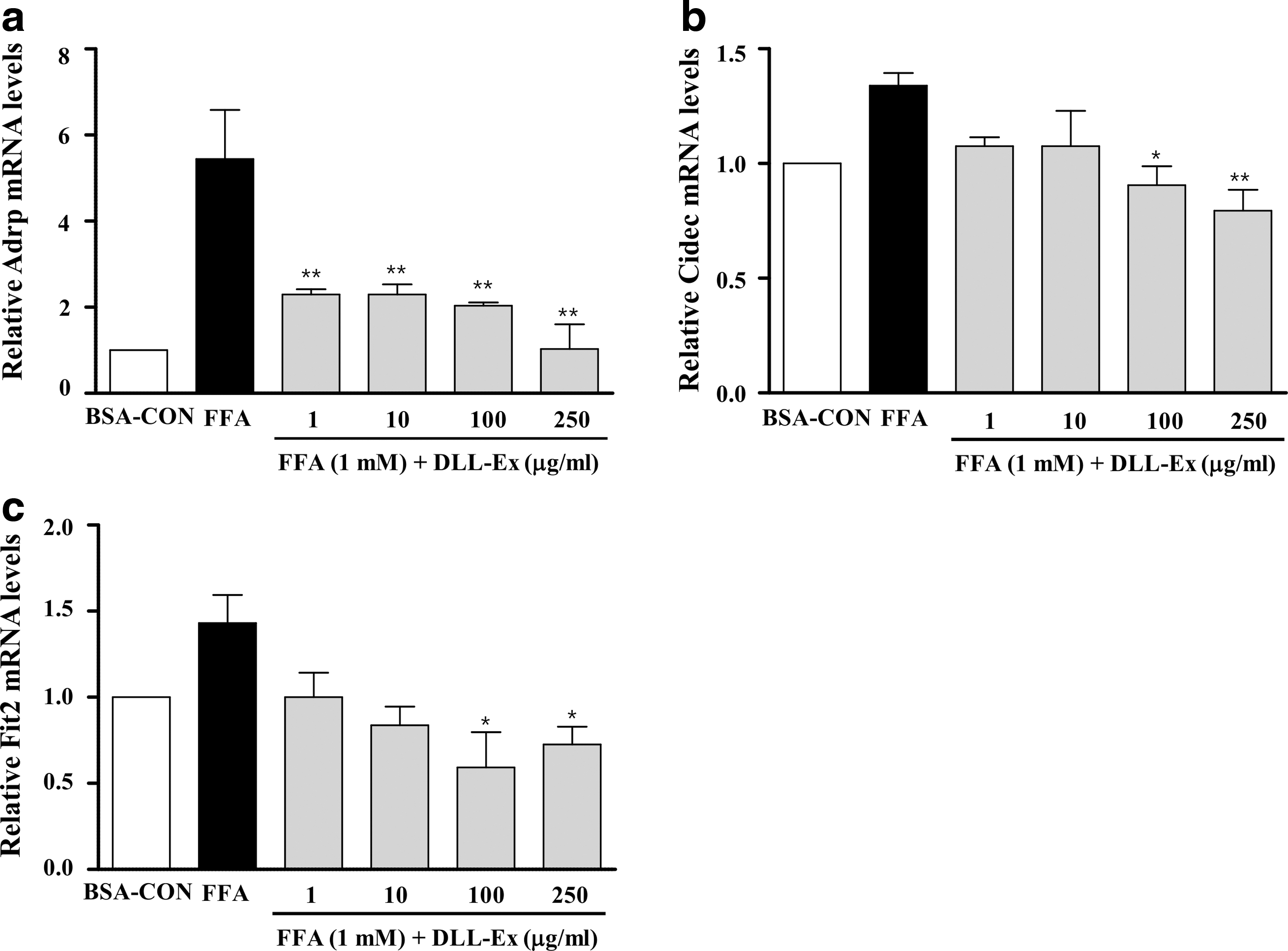
DLL-Ex attenuates lipogenic mRNA expression induced by FFAs. Cells were treated with 1 mM FFAs in the presence or absence of DLL-Ex (1, 10, 100, and 250 μg/mL) for 24 h. Total RNA was isolated for real-time quantitative polymerase chain reaction (PCR) analysis of
BODIPY-labeled FFA uptake in DLL-Ex-treated cells (10, 50, and 100 μg/mL) was significantly reduced in comparison with that of the control cells (Fig. 3a). Expression of FFA transporter CD36 was significantly and dose-dependently decreased by DLL-Ex treatment (100 and 250 μg/mL) (Fig. 3b). Furthermore, very low-density lipoprotein receptor (VLDLR) expression was significantly induced by FFA treatment and attenuated by DLL-Ex treatment (10, 100, and 250 μg/mL) (Fig. 3c).
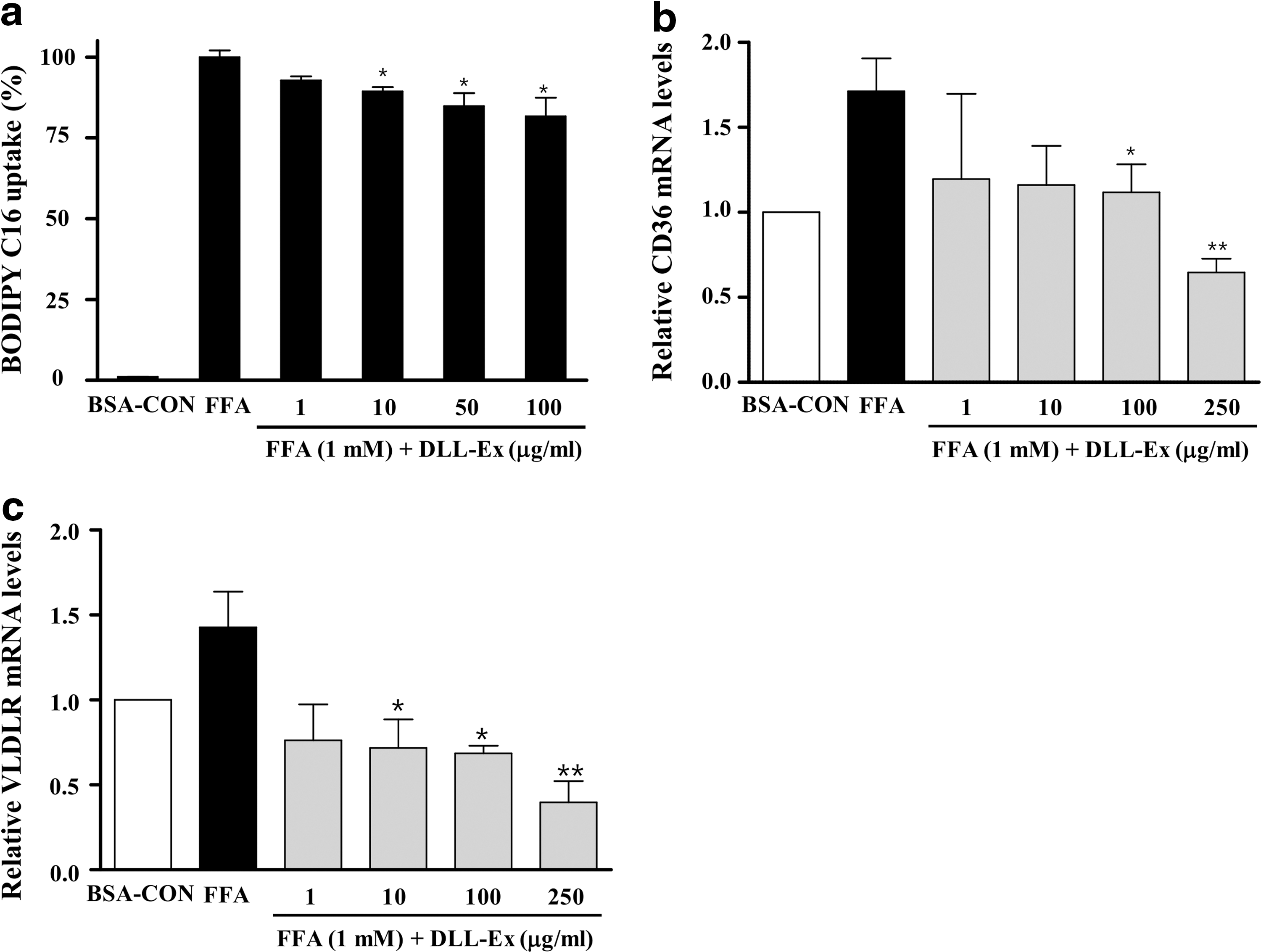
Effect of DLL-Ex on fatty acid uptake.
DLL-Ex prevents FFA-mediated cellular energy depletion and mitochondrial membrane depolarization
Measurement of ATP content in HepG2 cells revealed that the ATP level of the FFA-treated cells was decreased in comparison with that of the control cells; however, DLL-Ex (10, 50, and 100 μg/mL) attenuated this effect (Fig. 4a). Cells treated with FFA showed decreased red fluorescence intensity and increased green fluorescence intensity in comparison with the fluorescence observed in the untreated control cells, indicating decreased MMP and depolarization of the mitochondrial membrane (Fig. 4b). DLL-Ex-treated (10, 100, and 250 μg/mL) cells showed significantly greater MMP than that of cells treated with FFAs alone and close to that of control cells.
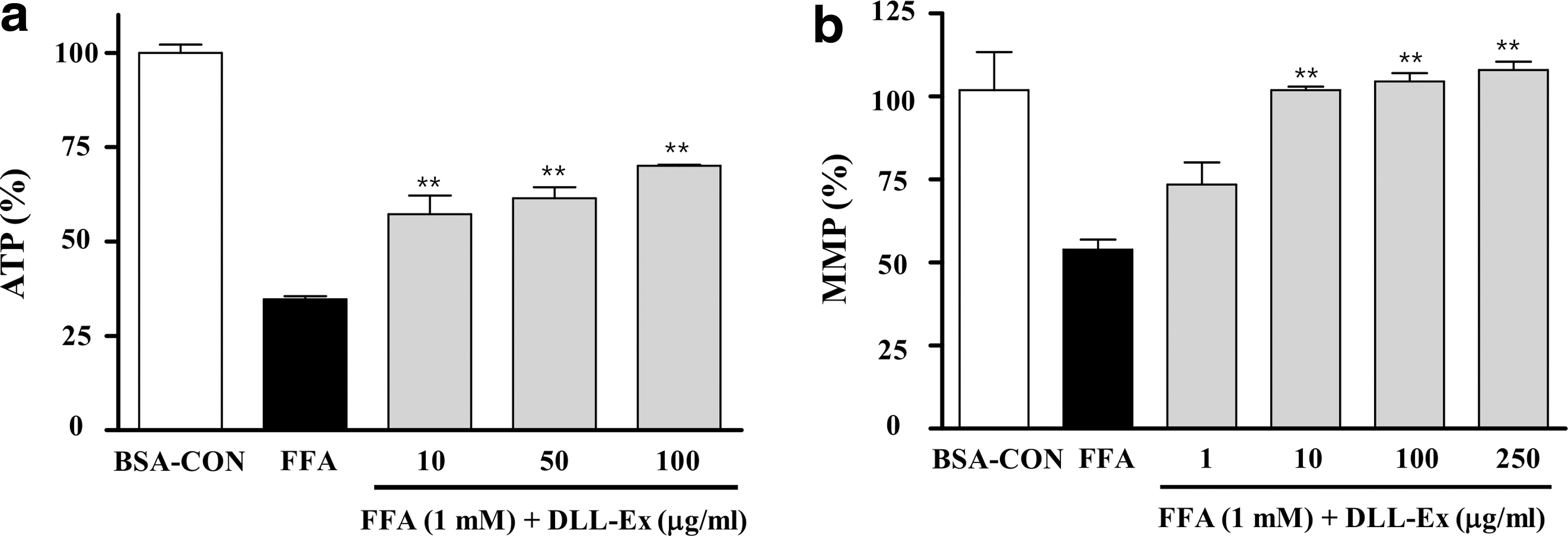
Effects of DLL-Ex on total cellular ATP levels and mitochondrial membrane potential (MMP).
DLL-Ex regulates FFA-induced mitogen-activated protein kinase
Investigation of the effect of DLL-Ex on FFA-induced cell signaling revealed that FFAs enhanced phosphorylation of ERK, JNK, and p38; however, DLL-Ex treatment (10, 50, 100, and 250 μg/mL) significantly reduced levels of phosphorylated ERK, JNK, and p38 in comparison with those of the cells treated with FFAs alone (Fig. 5a). Expression of peroxisome proliferator-activated receptor γ coactivator 1 alpha (PGC-1α) was also increased by DLL-Ex treatment (10 and 50 μg/mL) in HepG2 cells (Fig. 5b). Moreover, DLL-Ex treatment (10, 50, 100, and 250 μg/mL) significantly increased the abundance of phosphorylated AMPK in FFA-treated cells (Fig. 5c).
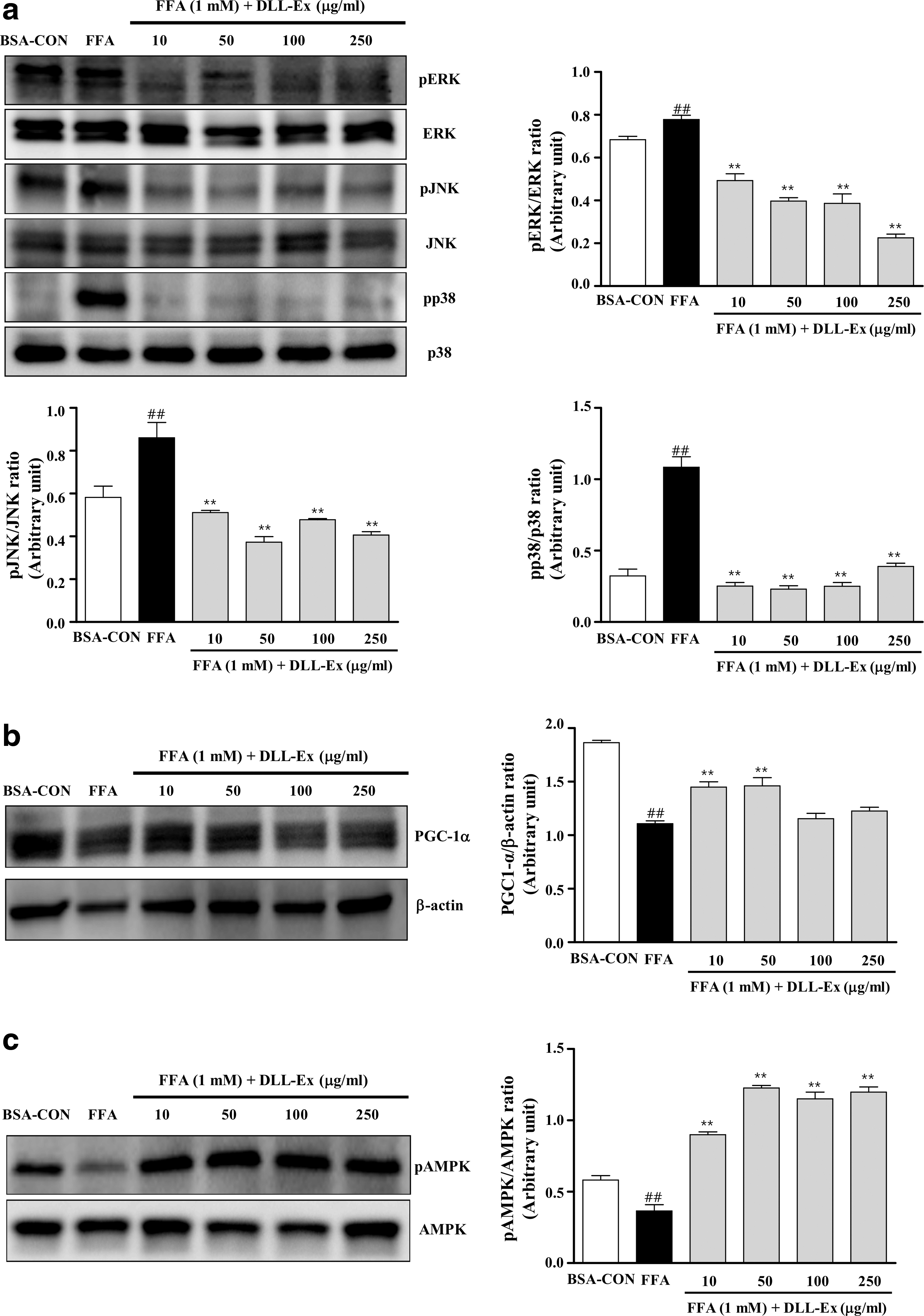
DLL-Ex activates AMP-activated protein kinase (AMPK) phosphorylation. Total protein was extracted from HepG2 cells and lysed.
Discussion
NAFLD is one of the most common forms of liver disease and is strongly related to other pathological conditions, including obesity, diabetes, cardiovascular diseases, and metabolic syndrome. 19 NAFLD encompasses a broad spectrum of liver diseases, ranging from simple fatty liver to NASH and cirrhosis. 9 All stages of NAFLD are associated with accumulation of TG droplets within hepatocytes, which is characterized by steatosis, inflammation, necrosis, and ultimately fibrosis and cirrhosis. 9 Although current NAFLD treatments focus on weight loss and management of cardiometabolic risk factors through correcting elevated cholesterol, TG, and blood sugar concentrations, no effective therapy has been developed.
The screening that led to the present study assessed more than 120 herbal medicine formulas and was aimed at facilitating the development of a novel, cytotoxicity-free herbal medicine to treat patients with NAFLD. This screening led to the development of DLL-Ex, a decoction of a bean that has been used in traditional Asian medicine for centuries. 20 In this study, an in vitro cellular model of hepatic steatosis was used to investigate the hepatoprotective activity of DLL-Ex in cells with FFA overload-induced steatosis. Although herbal medicines have been used by humans for millennia, the cytotoxicity of such materials must be considered before systematic clinical use. Therefore, the safety of DLL-Ex was confirmed using an in vitro MTS assay, whereas the efficacy of DLL-Ex was confirmed using an in vitro cellular model of hepatic steatosis. 21,22
The results described above demonstrate that pathophysiologically relevant concentrations of PA and OA induce lipid accumulation in hepatocytes. Morphological alterations were observed using a phase contrast microscope, whereas quantitative analysis was performed using Nile Red staining. Intracellular lipid content was increased dose-dependently by treatment with FFAs, which are generated by lipolysis in adipose tissue through the action of hormone-sensitive lipases, and provide a major source of fatty acids for liver TG secretion. Furthermore, changes in cellular lipid content were confirmed by Oil Red O staining and measurement of intracellular TG levels.
Previous studies have shown that expression of Fsp27/CIDEC is low in the livers of normal patients, whereas its expression is dramatically induced in patients with fatty liver. Furthermore, overexpression of Fsp27 in mouse primary hepatocytes leads to TG accumulation. 23 Intracellular lipid storage droplets are regulated by the perilipin family of lipid droplet coat proteins, including perilipin, ADRP, and myocardial lipid droplet protein/oxidative tissue-enriched PAT protein/lipid storage droplet protein 5. 24 In this study, DLL-Ex reduced lipid accumulation in hepatocytes by regulating genes that are important for hepatic lipid droplet accumulation and FFA uptake (Fig. 2). Cell-based assays also revealed that DLL-Ex repressed FFA uptake through repression of FAT/CD36 expression. Because CD36 deletion protected against hepatic steatosis and hepatic CD36 gene expression was increased and correlated with liver fat content in patients with NAFLD, FFA uptake inhibition in hepatocytes could theoretically be influenced by inhibition of CD36 expression. 23,25,26
Mitochondrial ATP production is highly dependent on the inner MMP during coupled respiration. 27 Mitochondria are involved in many processes essential for liver cell survival, including energy production, redox control, calcium homeostasis, and certain metabolic and biosynthetic pathways. 28 Hepatic mitochondrial dysfunction is well documented in the progression of metabolic disease, with impairments in electron transport chain complexes and subsequent depression of ATP levels contributing to the progression of NAFLD and NASH. 29,30 HepG2 cells exposed to FFA exhibited suppressed oxidative phosphorylation, as indicated by reduced MMP, ATP content, and oxygen consumption. 31 Additionally, a previous study showed that incubation of human and murine hepatocytes with FFAs dose-dependently induced mitochondrial dysfunction. 28 Our results showed that FFA treatment induced cellular oxidative stress and ROS generation in parallel with decreased cellular ATP levels (Fig. 4a); however, these changes were ameliorated by DLL-Ex treatment. These results are consistent with previous reports that DLL-Ex exhibits antioxidative effects and free radical scavenging activity. 14
Fatty acids are hypothesized to promote the progression of steatohepatitis by promoting fatty acid-induced cellular stress and organelle toxicity. 26 In previous studies, exposure to palmitate promoted JNK and p38 phosphorylation in HepG2 cells. 32 In this study, mitogen-activated protein kinases (MAPKs) were activated by FFA; however, DLL-Ex attenuated this effect. AMPK acts as a cellular energy gauge and phosphorylates target proteins to increase or decrease their activity in response to low cellular energy levels. 33 In the present study, DLL-Ex markedly enhanced AMPK phosphorylation after FFA overloading, indicating that AMPK activation by DLL-Ex contributes to DLL-Ex-mediated repression of lipid accumulation in FFA-overloaded HepG2 cells. The results described above show that newly developed herbal extract DLL-Ex represses cellular lipid accumulation and downregulates mRNA levels of genes associated with lipid droplet accumulation and FFA uptake in HepG2 cells. The sustained anti-NAFLD effects of DLL-Ex included alleviation of cellular energy depletion and mitochondrial membrane depolarization, which contributed to its antioxidative effect. The results of the present study also suggest that activation of the AMPK signaling pathway and suppression of MAPK activation by FFAs may play important roles in the lipid-lowering effect of DLL-Ex.
DLL-Ex contains several phytochemicals with strong antioxidant activity. A basic separation of steroidal plant growth regulators, dolicholide, dolichosterone, homodolicholide, homodolichosterone, brassinolide, castasterone, 6-deoxocastasterone, 6-deoxodolichosterone, and saponin conjugated with 2,3-dihydro-2,5-dihydroxy-6-methyl-4H-pyran-4-one from immature seed of D. lablab was accomplished using various chromatographic techniques. 34 –37 Four soy isoflavones, daidzein, daidzin, genistein, and genistin, were identified in the leaves of D. lablab by high-performance liquid chromatography coupled with high-resolution mass spectrometry/tandem mass spectrometry. 37 We speculate that the multitarget anti-NAFLD activity of DLL-Ex might be related to these multiple active components. Preclinical in vivo trial results are required to confirm the results described above. Studies using mice with diet-induced obesity will be carried out by our research group to confirm the anti-NAFLD activity of DLL-Ex.
Taken together, the results of the present study demonstrate that the sustained anti-NAFLD effects of DLL-Ex included alleviation of lipid accumulation, improvement of hypertriglyceridemia, decreased of FFA uptake into hepatocytes, reversal of mitochondrial dysfunction, and activation of AMPK. These results provide insight into the molecular mechanisms underlying the therapeutic effects of DLL-Ex and suggest that DLL-Ex could be used to reduce the risk of NAFLD and related diseases such as obesity and dyslipidemia in humans.
Footnotes
Acknowledgment
This research was supported by a grant for the “Development of Herbal Medicine for Aging-Related NAFLD” (K15302) funded by the KM Convergence Research Division of Korea Institute of Oriental Medicine (KIOM).
Author Disclosure Statement
No competing financial interests exist.