
Abstract
The objective of the present study (
Introduction
I
The skeletal muscle is made up of heterogeneous muscle fibers with distinct metabolic and contractile properties. 9 Changes in gene expression is an integral component of skeletal muscle physiological adaptations to exercise and nutritional supplementation. 9 Shilajit has been reported to improve physical performance and relieve fatigue with enhanced adenosine triphosphate (ATP) production. 4,8,10 The primary aim of the present longitudinal study (the same person's initial visit serves as the baseline for subsequent visits) is to find out the effect of oral Shilajit supplementation and exercise training on human skeletal muscle adaptation in a group of overweight/class I obese human subjects following 12 weeks of study period. This particular cohort was selected for the study because physiological muscle performance is often compromised in this group of subjects. 11 –13
Materials and Methods
Natreon Inc.'s patented ingredient, PrimaVie® [US 6,969,612; 6,440,436; 6,558,712; 8,894,993; and EP 1 387 614], is a purified and standardized Shilajit extract for nutraceutical use. It is standardized to have not less than 60.3% fulvic acid equivalents with DBP and associated chromoproteins (50% fulvic acid + 10.3% free DBPs and DBPs conjugated with chromoproteins) in Shilajit extract. The test product, PrimaVie Shilajit (PVS) capsules, 250 mg, was supplied by Natreon, Inc., New Brunswick, NJ, USA. PVS is manufactured by a process to reduce the heavy metals to less than 1 ppm of lead, 1 ppm of arsenic, and less than 0.1 ppm of mercury. Quality control is achieved through high-performance liquid chromatography (HPLC) analysis as previously reported. 14
Ethics, consent, and permissions
The Western Institutional Review Board (WIRB) approved the study protocols (
Study subjects and experimental design
Overweight/Class I obese, adult human subjects (21–70 years) of both genders with body–mass index (BMI) 25–35 were entitled to participate in this study. They were asked to fast overnight, following which collection of blood samples was done. Any self-reported variations in diet or exercises were recorded. The subjects were excluded from the study if any one of the following medications was used for management/treatment of cardiovascular disease (CVD)-related disorders: steroids (Prednisone, etc.), beta-blockers, hydrochlorothiazide, statins (Crestor, Lipitor, etc.), aspirin, and angiotensin-converting enzyme (ACE) inhibitors. Pregnant females as well as individuals who were therapeutically immunocompromised were also excluded from the study. The experimental study design consisted of four study visits during the 12-week study period: visit 1, baseline visit; visit 2, after 8 weeks of oral supplementation of PVS; visit 3a, additional (following 8 weeks of initial supplementation) 4 weeks of oral supplementation and exercise, sample collection before the final bout of exercise; and visit 3b, same as study visit 3, sample collected 30 min post-final bout of exercise. At each study visit, 50 mL of blood, 5 mm muscle biopsy, and demographic information, including age, gender, weight, BMI, blood pressure, and pulse, were taken (Table 1).
Values are expressed as mean ± SEM.
Supplementation regimen and compliance
Each subject received 250 mg of PVS capsules twice a day for the first 8 weeks of study period. For the last 4 weeks of the study, subjects took 250 mg of oral PVS supplement twice a day while also completing exercise on a treadmill (70–75% of maximum heart rate for 20 min monitored using Polar FT4, plus 5 min of warm-up and 5 min of cool-down exercises for a total of 30 min a day, 3 days a week). The same exercise regimen was followed during visits 3a and 3b. All subjects participated in the total 12 weeks of study, including one baseline and three follow-up visits. The dose of PVS was chosen based on an earlier human supplementation study. 14 The 8-week period of supplementation was selected based on earlier studies on biochemical/genetic adaptations in skeletal muscles. 15,16
Safety monitoring
No adverse effect directly related to the dietary supplement was reported by clinical research staff.
Blood sampling and analysis
During each visit, peripheral venous blood was collected in heparinized tubes and transported on ice immediately for analysis. Among lipid profile total cholesterol, high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), triglyceride levels, calculated LDL-C, and non-HDL-C were measured using standard clinical lipid profile, 17 and creatine kinase (CK), glucose, and serum myoglobin were measured at the clinical laboratory of Ohio State University Wexner Medical Center.
Muscle biopsy collection
A biopsy was collected by a board-certified physician after application of local anesthetics to the site of biopsy (vastus lateralis) using a 100–120 V, 50–60 Hz, 600VA biopsy machine having 12-gauge SenoRx, stereotactic ultrasound Encor Probe (BARD Encor Ultra, breast biopsy system, Tempe, AZ, USA). Muscle samples were stored in liquid nitrogen for further analysis.
Affymetrix GeneChip® probe array analysis
GeneChip® probe array analysis was performed on RNA extracted from muscle biopsies collected on baseline (visit 1) and visit 2 (8 weeks post-supplementation) using Affymetrix GeneChip Human Transcriptome Array 2.0 as described previously.
18,19
Briefly, total RNA was isolated and RNA integrity was interrogated using the Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). Total RNA was linearly amplified, labeled, and fragmented using the GeneChip WT PLUS reagent kit (Affymetrix, Santa Clara, CA, USA). Labeled cRNA targets hybridized to Affymetrix GeneChip Human Transcriptome Array 2.0 (HTA 2.0) for 16 h at 45°C in hybridization oven (Affymetrix model 640) rotating at 60 rpm were washed, stained, and scanned in our own facilities as described earlier.
18,19
The expression data have been submitted to the Gene Expression Omnibus (GEO) at NCBI (
Quantification of mRNA expression by real-time polymerase chain reaction
Real-time polymerase chain reaction (PCR) was carried out to validate the extracellular matrix (ECM)-related genes identified using GeneChip probe array analysis. Real-time polymerase chain reaction (RT-PCR) was performed using double-stranded DNA binding dye SYBR Green-I as described previously. 21,22 GAPDH was used as a reference housekeeping gene.
Statistical methods
Multivariate linear regression was used to test if all 11 gene expression (ΔΔCT) values were jointly different across adjacent time points. Five comparisons were generated across various time points. The multivariate regression produces estimated differences along with their 95% confidence interval for each gene with a single P-value testing if all 11 ΔΔCT values of the genes were jointly different across adjacent time points. Multivariate normality was checked using standardized normal probability plots. If any values were not normal, then they were transformed using natural logarithms. A new multivariate linear regression model was used to check if patient lipids/glucose/muscle damage marker values were jointly different across adjacent time points. Lipids/glucose/muscle damage marker values were summarized using means and standard deviations for each of the three time points. All analyses were run using Stata 13.1; StataCorp, College Station, TX, USA.
Results
Analysis of glucose, lipid profile, CK, and serum myoglobin levels following oral PVS supplementation
Lipid profile measurements displayed no significant changes in the cholesterol, HDL-C, calculated LDL-C, total cholesterol/HDL, non-HDL–C, and triglycerides following 8 weeks of oral PVS supplementation compared with the baseline levels, suggesting the supplementation was well tolerated (Tables 2 and 3). Additionally, lipid profile levels at the week 12 pre and post-final exercise (visits 3a and 3b) showed no significant changes compared with the baseline levels at 8 weeks (Tables 2 and 3). Moreover, no changes were observed in other variables, including blood glucose and muscle damage markers, including CK and serum myoglobin levels, at all follow-up visits (Tables 2 and 3).
HDL, high-density lipoprotein; LDL, low-density lipoprotein.
Transcriptome profiling of skeletal muscle following oral PVS supplementation
To determine the changes in the transcriptomes of human skeletal muscle in response to oral PVS supplementation, muscle samples were collected during each visit. RNA extraction, target labeling, and GeneChip data analysis were performed using Affymetrix® Human Transcriptome Array 2.0 (HTA 2.0) as described previously. 18,19 The HTA 2.0 contains >6.0 million probes covering coding transcripts and exon–exon splice junctions and noncoding transcripts. 23 A total of 175 annotated probe sets were differentially (P < .01) regulated following 8-week supplementation compared with baseline visits (Fig. 1A). Top 20 candidates based on fold change (compared with baseline) that was altered (up or downregulated) in the supplementation group have been provided in Tables 4 and 5. Pathway analysis revealed an ECM-related cluster of probe sets that was significantly upregulated in the 8-week supplementation group compared with corresponding baseline visit (Fig 1B). This probe set included ECM genes: tenascin XB (TNXB), decorin (DCN), collagen (COL) (type I, III, V, VI, XIV), fibrillin 1 (FBN1), elastin (ELN), myoferlin (MYOF), and fibronectin 1 (FN1) (Table 6). Among these upregulated genes, TNXB was increased by ∼1.7-fold and DCN was increased by 2.23- and 1.09-fold, respectively (Table 6). COL1A1, COL1A2, and COL3A1 were increased by 4.61-, 5.13-, and 5.18-fold, respectively, and ELN, FBN1, and FN1 were increased by 1.13-, 3.05-, and 3.65-fold, respectively (Table 6).
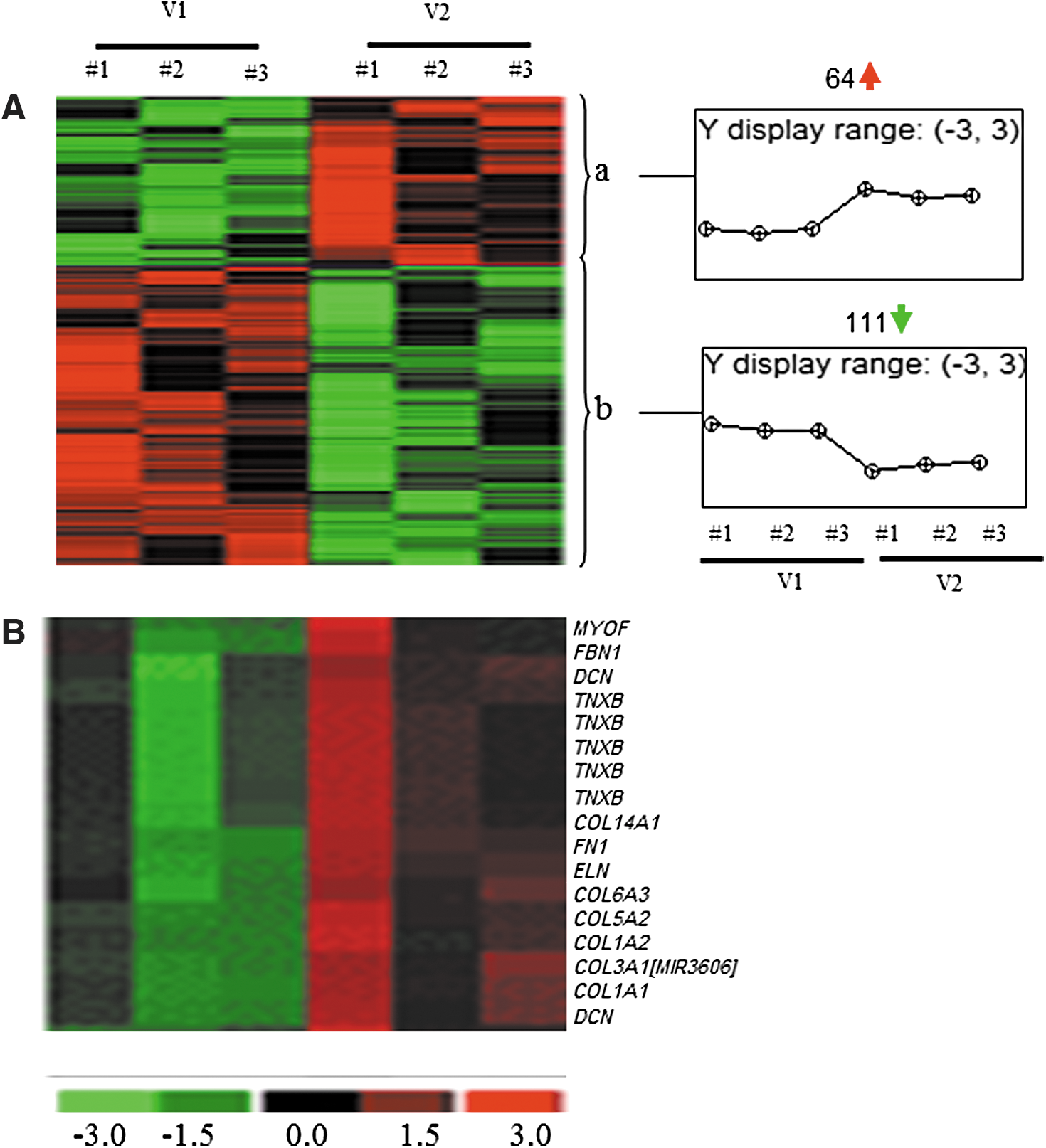
Heat map illustrating cluster of transcripts that were sensitive to PVS supplementation. PVS-sensitive transcripts were subjected to hierarchical clustering.
Data presented indicate fold changes in expression of PVS-sensitive genes following 8 weeks of oral PVS supplementation compared with corresponding baseline visits. Transcripts cluster ID, Affymetrix probe identifications. Data correspond to Figure 1A, cluster a (P < .05; FDR <5%).
Data presented indicate fold change in expression of PVS-sensitive genes following 8 weeks of oral PVS supplementation compared with corresponding baseline visits. Transcripts cluster ID, Affymetrix probe identifications. Data correspond to Figure 1A, cluster b (P < .05; FDR <5%).
Data presented indicate fold change in expression of ECM-related genes following 8 weeks of oral PVS supplementation compared with corresponding baseline visits. Data correspond to Figure 1B (P < .05; FDR <5%).
ECM, extracellular matrix.
Validation of GeneChip data using RT-PCR
To validate the genes identified using microarray analysis, RT-PCR was performed. Since the above-mentioned ECM-related genes were upregulated, those genes were reexamined. Consistent with microarray results, significant upregulation of collagen and other ECM-associated genes was noted in the muscle samples following 8 weeks of oral PVS supplementation compared with the baseline visit (Fig. 2 and Table 7). Interestingly, additional 4 weeks of oral supplementation with exercise further induced the expression of the microarray-identified genes (Fig. 2 and Table 7).

RT-PCR validation of ECM-related genes derived from microarray analysis following oral PVS supplementation. Expression levels of selected collagen genes identified using GeneChip® analyses were independently verified using real-time quantitative (Q) PCR. The effects of oral PVS supplementation (250 mg/b.i.d.) were measured during the course of all visits; V1, baseline; V2, after 8 weeks of oral supplementation; V3a, additional (following 8 weeks of initial supplementation) 4 weeks of oral supplementation and exercise, sample collection before the final stint of exercise; and V3b, same as study visit 3, sample collected 30 min post-final bout of exercise. Data are mean ± SEM (n = 16); *P < .05 compared with the baseline visit and † P < .05 compared with 8 weeks. No significant changes were observed between pre and post 30-min final exercise on week 12. PCR, polymerase chain reaction.
Discussion
Beneficial effects of Shilajit in skeletal muscle adaptation have been highlighted for centuries in ancient Ayurveda medicine. 4,5 The underlying mechanisms of such adaptation have not been elucidated. The current study, for the first time, presents a mechanism of action of Shilajit in improving skeletal muscle adaptation in overweight/obese subjects exercising 30 min a day 3 days a week for 4 weeks. ECM plays an essential role in the development, maintenance, and regeneration of skeletal muscles. 24,25 Chronic loading of muscles such as physical training leads to augmented synthesis and turnover of ECM. 26 Oral supplementation of PVS markedly enhanced ECM-related gene expression in overweight/class I obese human subjects. PVS is one of the very few nutritional supplements that induce skeletal muscle adaptation through upregulation of ECM genes.
During obesity, there is an increase in circulating lipids (free fatty acids, triglycerides) that accumulate in muscle as triacylglycerol as well as fatty acid metabolites such as ceramide, diacylglycerol, and long-chain acyl CoA. 27 No alterations in blood lipid profiles of subjects after supplementation suggest that PVS was well tolerated in this class of human subjects. Phosphocreatine (PCr) is one major source of ATP replenishment in tissues with rapidly shifting energy demand. 28 The CK reaction mediates this supply, in which creatine and adenosine diphosphate (ADP) are reversibly phosphorylated to PCr and ATP, respectively. 28 Functioning as a spatial and temporal buffer of ATP levels, the PCr–CK system requires a high level of total cellular creatine in mammal skeletal muscle. 29 So, reduction in CK may disturb ATP formation within skeletal muscle. High intracellular creatine concentrations are achieved by a combination of exogenous dietary intake and endogenous production, followed by cellular uptake of creatine from blood vessels. 29 The unaffected levels of serum CK in our study provided evidence for skeletal muscle integrity following oral supplementation. The high amounts of myoglobin in the skeletal and cardiac muscles enable storage and diffusion of oxygen in these tissues. 30 The unchanged levels of serum myoglobin and blood glucose on all visits confirmed that PVS was well tolerated and maintained physiological body glucose metabolism, homeostasis, and muscle integrity in the skeletal muscle of overweight/class I obese human subjects.
Skeletal muscle is highly plastic and well known to undergo significant adaptive modifications in response to both endurance and resistance exercise. 31 –34 Increasing evidences now indicate that in response to nutrition, skeletal muscle undergoes adaptive changes through regulatory processes driven by changes in gene expression and cell signaling. 35,36 ECM of skeletal muscle mainly comprises glycoproteins, collagen, and proteoglycans and plays a major role in mechanotransduction, that is, conducting force laterally between fibers and tendons. 37 –39 In our study, both microarray and RT-PCR results revealed elevated mRNA expression of collagen (type I, III, V, VI, and XIV) in response to oral PVS supplementation. The major structural protein in skeletal muscle ECM, collagen, comprises 1% to 2% of the muscle tissue and represents 6% of the weight of tendinous muscles. The skin comprises mainly collagen type I, which constitutes about 70% of collagen, with type III being 10% and trace amounts of collagen types IV, V, VI, and VII. With strenuous exercise, a rapid increase in the synthesis of collagen in tendons and muscles has been noted in mice and humans. 26,40 Increase in the expression of COL1A2, COL3A1, and COL5A1 genes enhances cell proliferation and active remodeling of ECM in tissue repair. 41 Exercise-induced ECM synthesis leads to protein degradation through increased matrix metalloproteinase (MMP) activity. 26 Microarray data show an increase in MMP-2 gene expression in muscles following oral supplementation, suggesting that the effects of PVS supplementation on skeletal muscle adaptation are comparable with exercise by mediating specific synthesis and degradation of ECM. Growth factors such as TGF-β and IGF-1 are involved in regulation of ECM synthesis in connective tissue. 26 The concentration of these growth factors has been shown to increase following exercise. Interestingly, increased expression of IGF1R was noted through microarray analysis. It is plausible that PVS induces ECM gene expression through comparable mechanisms. Further studies are required to determine exact mechanisms of PVS-induced ECM gene expression changes.
Although collagen provides the main structure, other ECM components also play an important role in skeletal muscle adaptation. In our study, both microarray and RT-PCR results revealed elevated mRNA expression of other ECM components including decorin, fibronectin, fibrillin, tenascin XB, myoferlin, and elastin, in response to oral PVS supplementation. Decorin is a small leucine-rich proteoglycan and contributes both to the formation and stabilization of collagen fibers in the perimysium that support muscle fibers assembled with myogenesis. 42 Fibronectin plays a major role in synthesizing provisional granulation tissue during the early phases of wound repair. 41 Fibrillin, a type of microfibril, is also one of the key structural elements in the ECM of skeletal muscle. Being widely distributed in connective tissues, fibrillins are arranged in tissue-specific architectures. 43,44 The other ECM component, tenascin XB, determines the mechanical properties of collagen. 45 During the development of muscles, especially during myoblast fusion, myoferlin is highly expressed 46 and regulates the reutilizing of vascular endothelial growth factor receptor-2. 47 The levels of myoferlin are generally less in adult skeletal muscle and almost lacking in healthy myofibers. An increase in the myoferlin level leads to a buildup of mononuclear myoferlin-positive myoblasts that play a key role in the repair of damaged myofibers, suggesting the importance of this gene in muscle repair and regeneration. 46
Conclusions
The current study reports for the first time that oral supplementation of a natural product to overweight/class I obese human subjects resulted in skeletal muscle adaptation through upregulation of ECM-related genes that control muscle mechanotransduction properties, elasticity, repair, and regeneration.
Footnotes
Acknowledgments
Parts of this work were supported by National Institutes of Health awards, GM077185, NR015676, and NR013898, and by a research grant from Natreon, Inc., NJ, USA.
Author Disclosure Statement
The authors declare that PVS and partial research funding were provided by Natreon, Inc., NJ, USA.