
Abstract
The aim of the present study is to investigate the potential protective role of Ceratonia siliqua L. against dextran sodium sulfate (DSS)-induced oxidative damage and inflammation in liver and kidney of rats. The hepatotoxicity and nephrotoxicity were induced in rats by oral administration of synthetic DSS (5%) in the drinking water for over 7 days. However, carob pods aqueous extract (CPAE; 50 and 100 mg/kg body weight) was given by oral administration for 21 days. Myeloperoxidase (MPO) activity, malondialdehyde, H2O2 content, as well as the levels of antioxidant enzymes in organs were measured to observe the possible mechanisms. As a result, the CPAE counteracted DSS-induced increase of MPO activity, lipoperoxidation, and the activity of antioxidant enzymes, such as superoxide dismutase and catalase (CAT). DSS administration increased also in the organs hydrogen peroxide (H2O2) and free iron levels, whereas the CPAE pretreatment reversed all intracellular mediator perturbations. It was concluded that the CPAE exerted a potential protective effect against DSS-induced inflammation and oxidative stress in the rat organs. Consequently, it is essential that adequate care is taken when we use carob pods for patients with hepatotoxicity and nephrotoxicity.
Introduction
D
The carob tree (Ceratonia siliqua L.) is a typical Mediterranean plant. 3 In many Arabian countries, the fruit is used for preparing popular beverages and confectionery. In Western countries, carob powder is produced by deseeding carob pods. The dried powdered pods of the carob plant are used as a dietary supplement and in herbal medicine. Few investigations of the pharmacological properties of C. siliqua L. are described in the literature, including antiproliferative effects on certain tumor cell lines, 4 cytotoxic and antimicrobial activity against resistant strains of Staphylococcus aureus and Escherichia coli, 5 and antioxidant activity. 6 Carob pods and leaves may be also effective to protect from oxidative renal damage and the leaves are the better nephroprotective agent than pods. The protection may be mediated partially by preventing the decline of renal antioxidant statues. 7 In addition, our laboratory recently reported the antiulcerogenic activity in model of acute gastric ulcer and potent inhibitor of human neutrophils myeloperoxidase (MPO) and in vitro reactive oxygen species (ROS)-scavenging activity. 8,9 We thus investigated the possible action mechanisms involved in the protective effect of the C. siliqua L. on hepatotoxicity and nephrotoxicity induced by DSS (5%) in rat.
Materials and Methods
Chemicals
Epinephrine, bovine catalase (CAT), butylated hydroxytoluene (BHT), 2-thio-barbituric acid, 5,5-dithio bis(2-nitrobenzoic acid) (DTNB), bovine serum albumin, p-hydroxybenzoic acid (PHBA), 4-aminoantipyrine (C11H13N3O), 3, 3′, 5, 5′-tetramethylbenzidine (TMB), and trichloroacetic acid (TCA) were from Sigma Chemicals Co. (Germany). All other chemicals used were of analytical grade.
Ethics statement
These experiments were approved by our Institutional Committee on Animal Care and Use and the experimental protocol complied with Tunisian legal requirements for animal studies. 10
The mature carob pods were collected from the region of Tabarka, (North West Tunisia) during October 2014. The necessary permits for the field studies and collection of C. siliqua pods were obtained from the Ministry of Agriculture in Tunisia and identified by Mrs. Mouhiba Ben-Naceur, professor of taxonomy in the Higher Institute of Biotechnology of Beja-Tunisia. The voucher specimens (No. CS130) have been deposited with the herbarium of the Higher Institute of Biotechnology of Beja.
Animals
Adult male Wistar rats (170–200 g, 10 weeks old and housed five per cage) were purchased from the Society of Pharmaceutical Industries of Tunisia (SIPHAT), and before any experience, all animals were kept one week under the same laboratory conditions of temperature (22°C ± 2°C), relative humidity (70% ± 4%), and a 12-h light–12-h dark cycle, and received a nutritionally standard diet and tap water.
Extract preparation
The plant material was later dried in an incubator at 50°C during 72 h and powdered in an electric blender (Moulinex Ovatio 2, FR). The powdered mixture containing carob pulp (90%) and seeds (10%) was dissolved in double distilled water and filtered through a colander (0.5-mm mesh size). Finally, the carob pods aqueous extract (CPAE) was immediately used for experiments.
Study design
Animals were randomly divided into four groups (10 rats per group):
Group 1 served as a control and received 1 mL of physiological solution (NaCl, 0.9%, p.o.).
Group 2 received DSS (5%).
Group 3 and 4 received DSS (5%) associated to 50 and 100 mg/kg of body weight of the CPAE taken orally.
During 14 days C and DSS rats drank tap water, whereas groups 3 and 4 received 50 and 100 mg/kg of the CPAE. From day 14 to 21, control rats were kept on drinking tap water, whereas DSS and CPAE-DSS rats received 5% DSS in their drink.
Histopathological examinations
The freshly excised liver and kidney were washed with saline and preserved in 10% formaldehyde solution for histopathological studies. The sections of organs stained with Hematoxylin and Eosin were assessed for histopathological changes, such as congestion, edema, hemorrhage, and necrosis. The microscopic slides were photographed. 11
Protein determination
The protein concentration was assessed according to the method reported by Hartree, 12 which is a slight modification of the Lowry method. Serum albumin was used as the standard.
Measurement of lipid peroxidation
Quantification of malondialdehyde (MDA) in renal tissue was determined by using the TBARS Assay Kit (Biomaghreb) according to the manufacturer's instructions. Tissue homogenate was prepared with PBS (phosphate buffered saline) and BHT to prevent oxidation and was mixed with thiobarbituric acid reagent. Supernatant from the mixture was read at 532 nm for MDA concentration.
H2O2 determination in liver and kidney
H2O2 level in liver and kidney was performed according to Dingeon et al. 13 Briefly, the hydrogen peroxide reacts with PHBA and 4-aminoantipyrine (C11H13N3O) in the presence of peroxidase leading to the formation of quinoneimine that has a pink color detected at 505 nm.
Measurement of MPO activity in liver and kidney
The supernatant of samples was assayed for MPO activity by measuring the hydrogen peroxide (H2O2)-dependent oxidation of 3, 3′, TMB. In its oxidized form, TMB has a blue color, which was measured spectrophotometrically at 650 nm. The reaction mixture for analysis consisted of a 25 μL tissue sample, 25 μL of TMB (final concentration 0.16 mM) dissolved in dimethylsulfoxide, and 200 μL of H2O2 (final concentration 0.30 mM) dissolved in phosphate buffer (0.08 M, pH 5.4). The reaction mixture was incubated for 3 min at 37°C and the reaction was stopped by adding 1 mL of sodium acetate (0.2 M, pH 3.0), after which absorbance at 650 nm was measured. 14
Determination of antioxidant enzyme activities in liver and kidney
The activity of superoxide dismutase (SOD) was estimated by the inhibition of nicotinamide adenine dinucleotide (reduced)-phenazine methosulfate-nitroblue tetrazolium reaction systems adapted by Kakkar et al., 15 and the results have been expressed as units (U) of SOD activity/mg protein.
CAT was estimated by a method of Aebi. 16 The standard assay was performed in 3 mL of 50 mM potassium phosphate buffer at pH 7.0 (1.9 mL) containing 10 mM H2O2 (1 mL) and tissue homogenate (100 μL). Under these conditions, the amount of CAT required to decompose 1.0 μM of H2O2 per min at pH 7.0 at 25°C was defined as 1 unit of activity. Absorbance was measured at 240 nm for 2 min, and the results were expressed as mM Cat/min/mg protein.
GPx activity was measured by the procedure of Flohé and Günzler. 17 In fact, the reaction mixture consisting of 0.3 mL of phosphate buffer (0.1 M, pH 7.4), 0.2 mL of GSH (2 mM), 0.1 mL of sodium azide (10 mM), 0.1 mL of H2O2 (1 mM), and 0.3 mL of tissue homogenate (liver and kidney) was incubated at 37°C for 15 min. Reaction was stopped by the addition of 0.5 mL of TCA (5%). The mixture was centrifuged at 1500 g for 5 min, and to the supernatant, 0.7 mL of DTNB (0.4 mg/mL) and 0.2 mL of phosphate buffer (0.1 M, pH 7.4) was added. The absorbance was recorded at 420 nm and the results were expressed as nmol of GSH consumed/min/mg protein.
Biochemical analyses of plasma
The blood of rats submitted to the different treatments was collected immediately after the sacrifice and submitted to centrifugation (3000 g for 10 min). After the centrifugation, the plasma obtained was frozen at −20°C until the biochemical analyses. Plasma biochemical parameters, including urea, creatinine, creatine kinase, aspartate aminotransferase (AST) and alanine aminotransferase, and lactate dehydrogenase (LDH) were measured by a commercially available kit from Biomaghreb, Tunisia.
Free iron measurement in liver and kidney
Free iron was determined by the FerroZine method using a commercially available kit from Biomaghreb, Tunisia. Briefly, at acidic pH 4.8, all Fe3+ are released from transferrin. Fe3+ released is reduced by ascorbic acid into Fe2+, which constituted with FerroZine a colorful complex measurable at 560 nm.
Statistical analysis
Results are expressed as mean ± standard error of the mean and presence of significant differences among mean of the groups was determined using one-way StatView ANOVA for significance. Values were considered significant when P < .05.
Results
Effect of DSS and CPAE on organs' weight and histopathological evaluation
Liver and kidney weights were significantly reduced in animals exposed to DSS (5%) for 7 days (Fig. 1). CPAE pretreatment significantly and dose dependently reversed the decrease of weight of organs induced by DSS intoxication. Nevertheless, no significant histopathological changes were observed in liver and kidney of DSS-exposed rats compared with healthy animals Figure 2a, b.
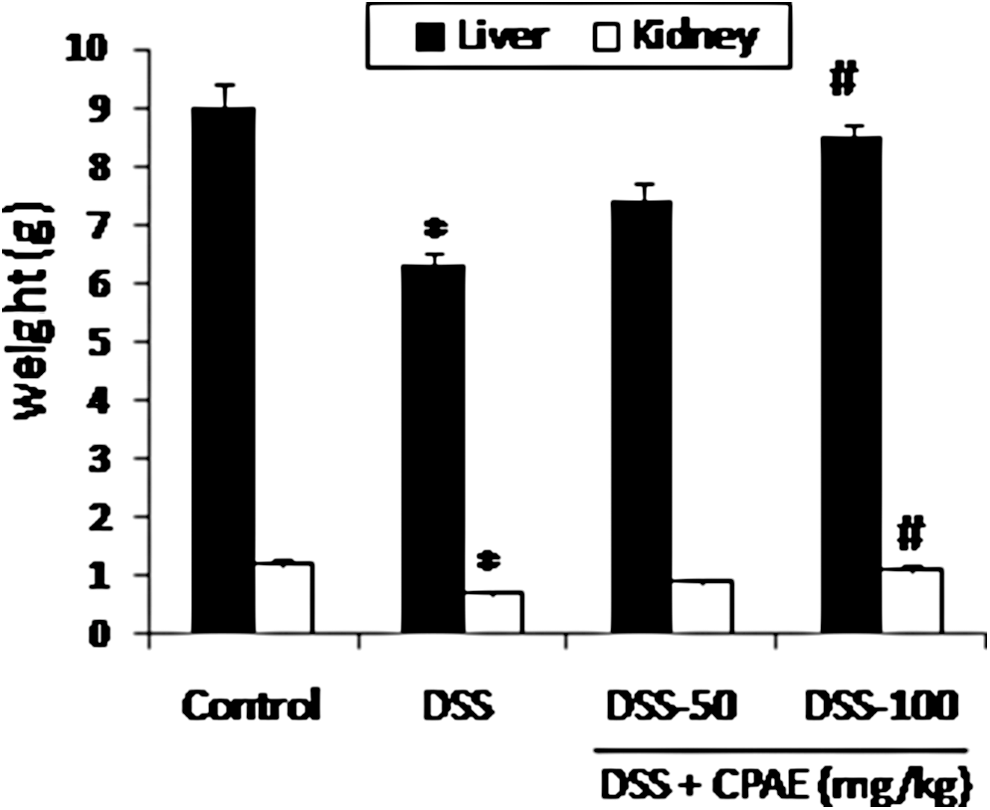
Subacute effect of CPAE on DSS-induced changes in organ weights. Animals were pretreated with various doses of CPAE (50 and 100 mg/kg, b.w., p.o.), or bidistilled water, challenged with oral administration of DSS (5%) or NaCl 9‰ for 7 days. *P < .05 compared with control group and # P < .05 compared with DSS group. Values are mean ± SEM (n = 10). CPAE, carob pods aqueous extract; DSS, dextran sodium sulfate; SEM, standard error of the mean.
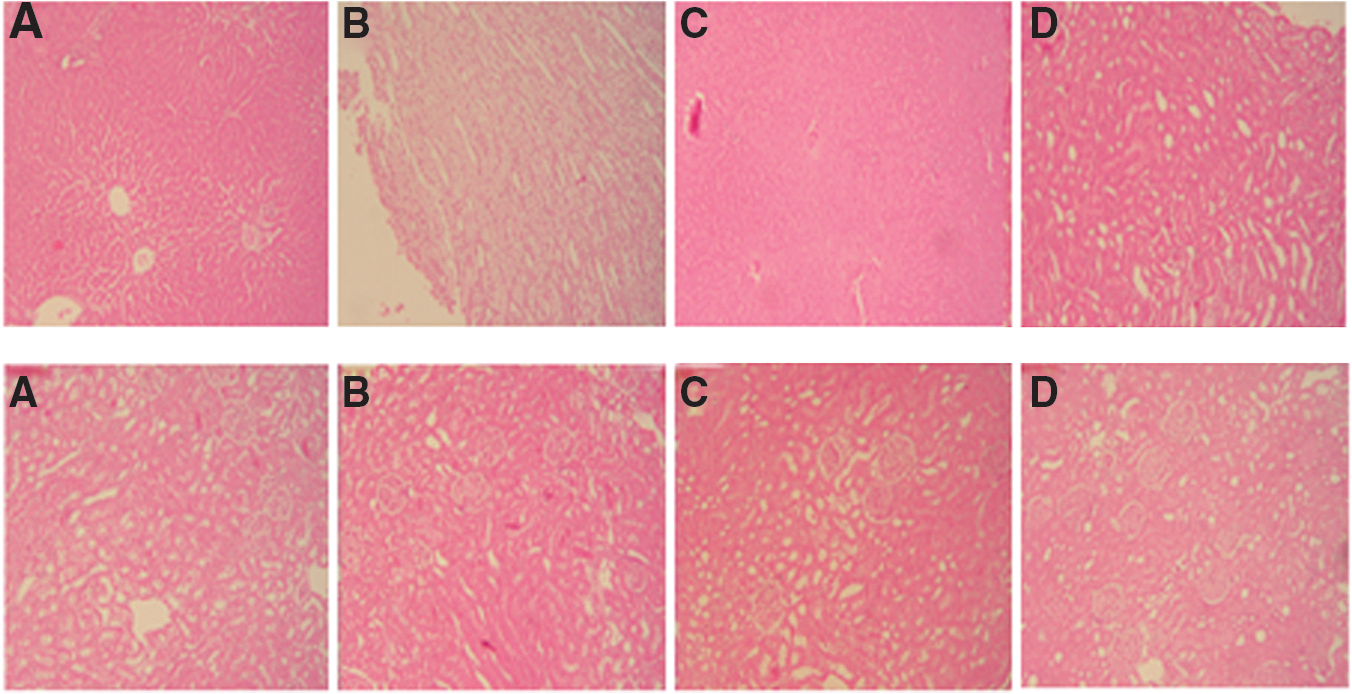
Histological analysis of liver
Effect of CPAE on DSS-induced protein oxidation, lipoperoxidation, and peroxide production in liver and kidney
The effect of DSS (5%) and CPAE treatment on the protein levels in liver and kidney was investigated and the results are presented in Figure 3a. DSS-induced ulcerative colitis was accompanied by a significant decrease in the protein content. Interestingly, in stress condition, we also studied the lipoperoxidation and production of MDA and hydrogen peroxide. In this context, DSS intoxication significantly increased the MDA (Fig. 3b) and H2O2 generation in both organs (Fig. 3c). CPAE pretreatment significantly and dose dependently reversed overproduction of MDA, protein levels, and hydrogen peroxide increase induced by DSS intoxication.
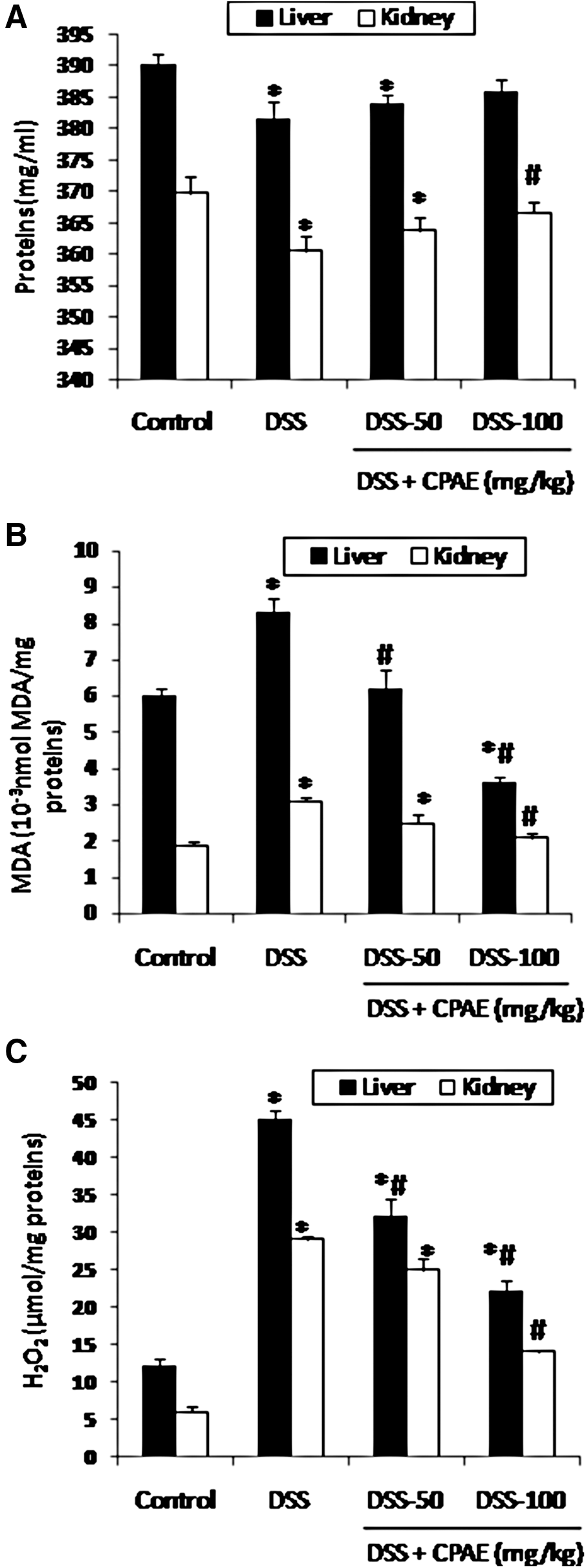
Subacute effect of CPAE on DSS-induced changes in liver and kidney on proteins
Effect of CPAE on DSS-induced MPO activity increase in liver and kidney
Bearing in mind the effect of DSS and CPAE on inflammatory condition, we studied the principal biomarker of inflammation (MPO activity) (Fig. 4). DSS (5%) intoxication drastically increased the MPO activity. CPAE pretreatment significantly and dose dependently reversed the MPO activity increase induced by DSS intoxication.
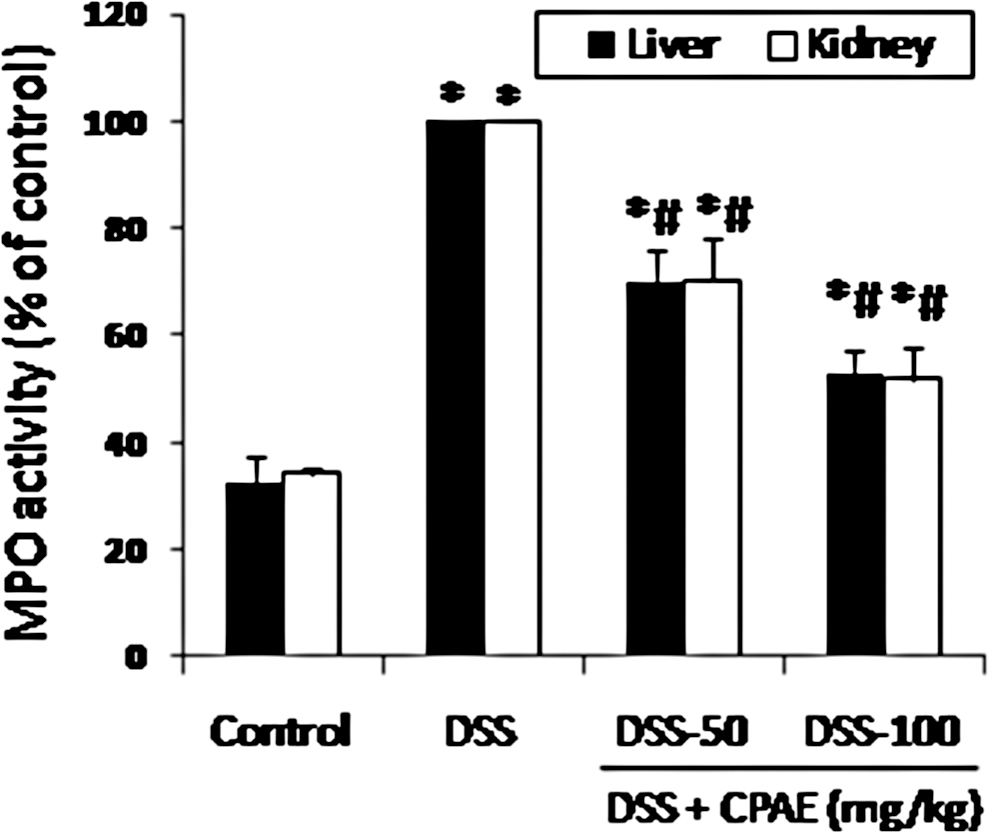
Subacute effect of CPAE on DSS-induced increase of MPO activity in liver and kidney in rats. Animals were pretreated with various doses of CPAE (50 and 100 mg/kg, b.w., p.o.), or bidistilled water, challenged with oral administration of DSS (5%) or NaCl 9‰ for 7 days. *P < .05 compared with control group and # P < .05 compared with DSS group. Values are mean ± SEM (n = 10). MPO, myeloperoxidase.
Effect of DSS and CPAE on antioxidant enzymes activities
The effect of DSS (5%) and CPAE (50 and 100 mg/kg) treatment on the antioxidant enzymes activities was also investigated and the results are presented in Figure 5. DSS-induced alteration in liver and kidney was accompanied by a significant increase in the activity of antioxidant enzymes such as SOD (A) and CAT (B). This modification was significantly and dose dependently improved by the CPAE (50 and 100 mg/kg, p.o.). However, the GPx (C) activity was not significantly modified.
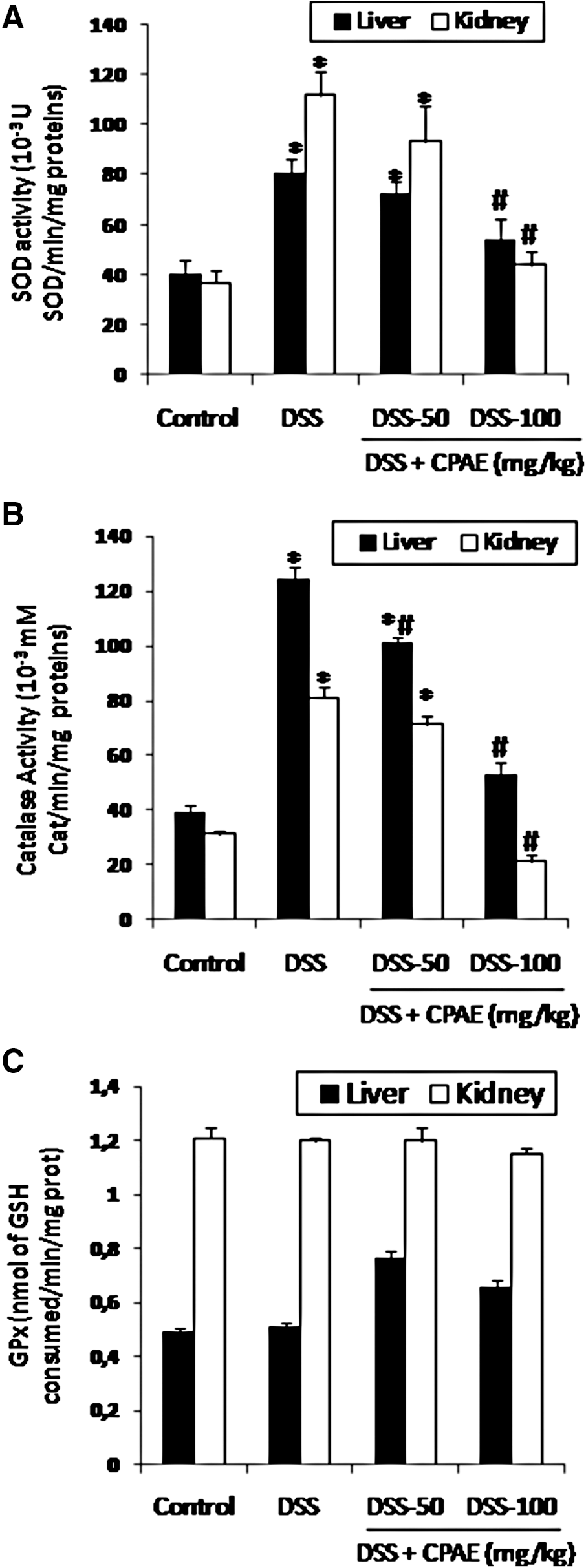
Subacute effect of CPAE on DSS-induced changes in liver and kidney of antioxidant enzyme activities in rats [SOD
Effect of DSS and CPAE on the functionality of liver and kidney
Further effects on the functionality of liver and kidney were assessed by determining the plasma levels of transaminases (ALAT and AST), LDH activity, creatinine, urea, and creatine kinase. The DSS (5%) induced significant alteration of hepatic and kidney functionalities by increase of these parameters. However, the CPAE pretreatment reversed all perturbations (Table 1).
Values are mean ± SEM (n = 10). Control group; DSS, DSS (5%) group; DSS + CPAE-50, DSS (5%) + CPAE (50 mg/kg, b. w.) group; DSS + CPAE-100, DSS (5%) + CPAE (100 mg/kg, b. w.) group. * P < .05 compared with control group and # P < .05 compared with DSS group. CPAE, carob pods aqueous extract; DSS, dextran sodium sulfate; LDH, lactate dehydrogenase; SEM, standard error of the mean.
Effect of DSS and CPAE on free iron level
We also looked at the effect of DSS and the CPAE on an intracellular mediator such as free iron. As expected, DSS administration increased the iron level. The CPAE pretreatment significantly protected against DSS-induced intracellular mediator disturbance in a dose-dependent manner (Fig. 6).
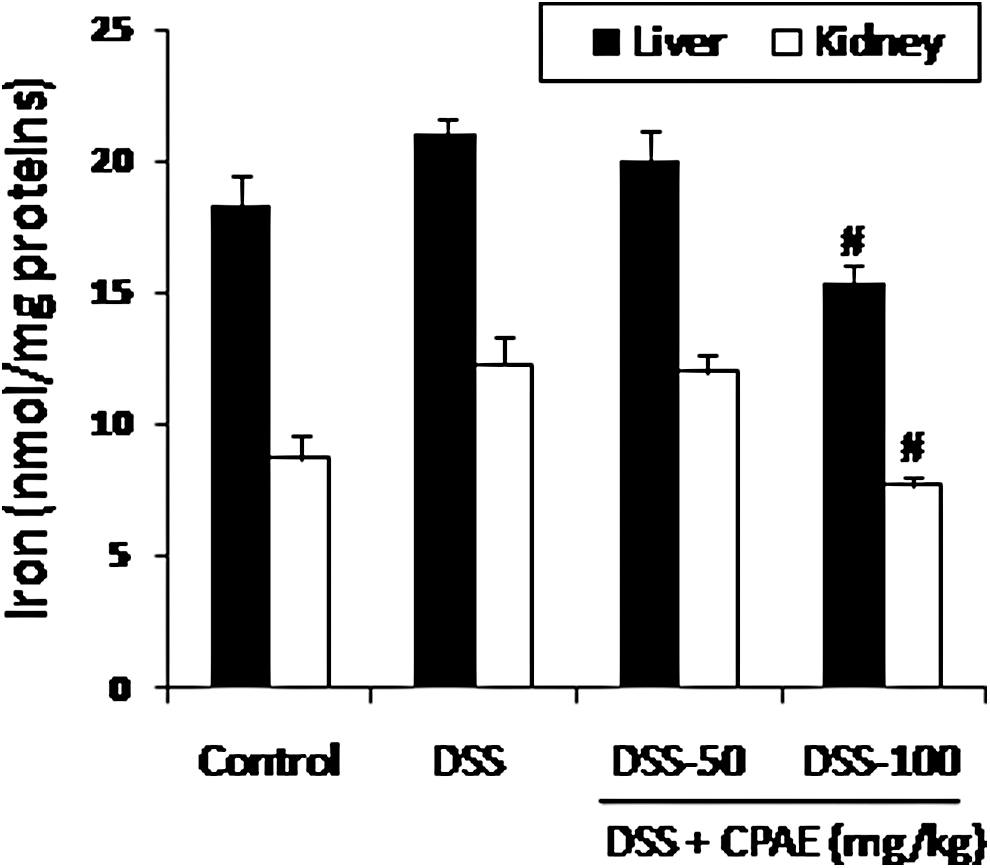
Subacute effect of CPAE on DSS-induced changes on iron free in liver and kidney of rats. Animals were pretreated with various doses of CPAE (50 and 100 mg/kg, b.w., p.o.), or bidistilled water, challenged with oral administration of DSS (5%) or NaCl 9‰ for 7 days. *P < .05 compared with control group and # P < .05 compared with DSS group. Values are mean ± SEM (n = 10).
Discussion
Numerous inflammatory diseases, including IBDs are multifactorial disorders with no therapeutic agents available to simultaneously control both inflammatory immune response and oxidative stress. 18 In the present study, we investigated the protective effect of carob pods on DSS-induced oxidative damage and inflammation in liver and kidney in rat.
First, in the acute oral toxicity study, neither abnormal behavior nor mortality was detected during the observation period. Thus, the LD50 value was greater than 20 g/kg b.w. for the CPAE. 8 Using the high performance liquid chromatography (HPLC) technique revealed the identification of phenolic compounds in carob pods with pyrogallol and catechin as main compounds in the pulp and tannic acid in the seeds. 9 In addition, using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) (ABTS) radical-scavenging assay, we found that CPAE of pulp and seeds exhibits a high scavenging capacity, although lesser than ascorbic acid, which was used as a reference molecule. 8
The oxidative stress has been increasingly recognized as a contributing factor in various forms of pathophysiology. Under physiological conditions, unstable, potentially cytotoxic molecules are produced, following the biochemical body reactions, called free radicals or ROS. 19 Biochemical lesions created by these molecules are multiple, inducing lipid peroxidation leading to membrane disruption, protein damage with fragmentation, aggregation, oxidation of sulfhydryl groups, and destruction of nucleic acids with DNA breakage and mutation. 20 In this respect, we showed in the present study DSS-induced lipid peroxidation and decrease of protein levels, and these effects were abolished by pretreatment of CPAE.
The increase in lipoperoxydation by DSS (5%) may suggest a possible mechanism of tissue injury by reactive oxygen intermediates. 21 Hydroxyl radicals thus generated oxidize important cellular constituents such as structural and functional proteins and membrane lipids, as well as deplete glutathione. Lipid peroxidation causes loss of membrane fluidity, impaired ion transport and membrane integrity, and finally loss of cellular functions. 22 The phenolic compounds are the primary source of the antioxidant ability of this plant, by scavenging free radicals as hydroxyl radical (OH•), which is the major cause of lipid peroxidation. 23 These data fully corroborate previous studies, indicating the herbal extracts, such as a Punica granatum, 24 resveratrol, 25 or purified polyphenols such as luteolin, 26 have also reported a decrease of lipoperoxidation.
On the other hand, the sequential elevation in the H2O2 level observed in liver and kidney of DSS-exposed rats indicates also a state of oxidative stress. Indeed, the deleterious chemical effects of H2O2 molecules are due to the direct activity, originating from their oxidizing properties, and the indirect activity in which they serve as a source for more deleterious species, such as hydroxyl radicals and hypochlorous acid. 27 Because of its richness in the phenol compounds, the CPAE is able to reduce the H2O2 generation in tissues following the installation of oxidative stress. In this context, we have shown that the CPAE has a scavenging activity against ROS, such as superoxide anion (O•−2) and hydrogen peroxide (H2O2) that have been generated in vitro by neutrophils stimulated by phorbol myristate acetate. 9
More importantly, we demonstrated that the CPAE treatment inhibits the MPO activity in the liver and kidney in vivo. MPO, which is released from cytoplasmic granules of neutrophils and monocytes by a degranulation process, reacts with the H2O2 formed by the respiratory burst to form a complex that can oxidize a large variety of substances. 28 Our recent results showed that CPAE caused a significant inhibition of two enzymes responsible for the production of ROS, such as MPO and NADPH oxidase by inhibition of p47-phox-ser328 in vitro. 9
The severity of oxidative stress depends on the extent of disturbances in normal redox state within the cells and tissues. 29 Endogenous antioxidant enzymes such as SOD, CAT, and GPx can limit the effects of ROS, but quickly become overwhelmed by large quantities of ROS. 30 DSS treatment resulted in the depletion of hepatic and kidney antioxidant status as evident in the significant increase in the activities of SOD and CAT. In fact, a helping effect of polyphenols on SOD and CAT activity has previously been reported in various models. Besides, a stimulating effect of polyphenols on antioxidant enzymes activity has previously been reported in various models. 31 In addition, we have shown that CPAE might have depleted ROS level, what relieved the alteration of the antioxidant enzymes and/or may have triggered their synthesis, which in turn attenuated the oxidative damage in DSS-treated rats. However, CPAE did not significantly change other antioxidative enzymes, such as GPx activity in DSS-treated rats. Thus, little is known about the effects of polyphenols on GPx activity. 26
Finally, in the present study, we showed that DSS intoxication induced an increase in liver and kidney free iron levels. The release of this metal ion from storage sites is effected for stimulating the conversion of H2O2 to OH• and catalyzing the breakdown of lipid peroxides to RO• 2 and RO•. 32 However, the CPAE protects against this homeostasis disturbance. Our data clearly demonstrate that carob pods exert protective effects against DSS-induced injury in the rat liver and kidney owing in part to their antioxidant and anti-inflammatory properties.
Footnotes
Acknowledgments
Financial support of the Tunisian Ministry of “Enseignement Supérieur et Recherche Scientifique” is appreciatively acknowledged. The study was made possible by international cooperation between INSERM U773, Faculty of Medicine Xavier Bichat, University Paris Diderot—Paris 7, France and Laboratory of Physiology and Nutrition Faculty of Sciences of Tunis, Tunisia.
Author Disclosure Statement
No competing financial interests exist.