
Abstract
Previous studies have shown that the precursor of olive oil secoiridoids, Oleuropein (OL) has several in vitro chemopreventive properties. OL inhibits proliferation and induces apoptosis in breast, thyroid, prostate, and colorectal cancer (CRC) cells. Much less is known about the effects of OL on animal models of carcinogenesis. In this study, we investigated the ability of OL to prevent the azoxymethane (AOM)-induced colon cancer upset and DNA damage in mice. Animals, fed with a basal diet either enriched or not with OL (125 mg/kg), were injected with AOM (10 mg/kg, once a week for 6 weeks) and sacrificed after either 7 weeks for histological analysis of colon crypt dysplasia and evaluation of DNA damage in leukocytes or 17 weeks for counting the macroscopically observable colon tumors. An OL-enriched diet prevented the AOM-induced preneoplastic lesions in different colon segments, reducing the severity of crypt dysplasia and DNA damage in peripheral leukocytes. In addition, OL significantly reduced the AOM-induced tumor incidence from 57% to 14% (P < .05, chi-square test) in the medial colon segment. This study shows that OL is able to prevent CRC and DNA damage in mice treated with the carcinogen AOM. These results stimulate further human cancer prevention studies with OL-enriched food supplements that are actually available on the market.
Introduction
C
Foods may contain factors that both increase and decrease the risk of CRC. Among the preventive components, the phenols extracted from olive oil have been demonstrated to inhibit several stages in colon carcinogenesis in vitro. 3 The major phenolic compounds in olive oil are phenolic acids, phenolic alcohols (hydroxytyrosol and tyrosol), secoiridoids (oleuropein [OL], ligstroside aglycone, and their decarboxylated dialdehyde derivatives), and lignans. 4 OL is the precursor of both hydroxytyrosol and other secoiridoids, is the most abundant compound present in the olive fruit and leaves, 5,6 and is known to possess several biological activities such as anti-microbial, 7 anti-inflammatory, 8 free radical scavenger, 9 anti-oxidant, and anti-cancer. 10
The cancer chemopreventive activity of OL has been deeply investigated in several in vitro systems. It has been shown that OL plays an important role in the prevention of DNA damage, 11 inhibits the proliferation, and induces apoptosis in breast, 12 thyroid, 13 prostate, 14 and CRC cells. 15 Recently, it was shown that OL reduces angiogenesis through inhibition of the VEGFR-2/mitogen-activated protein kinase (MAPK) signaling pathways in endothelial cells. 16
An interesting observation is that OL is not absorbed in the small intestine entering the large intestine, where it may interact with the gut microbiota and exert different healthy effects. 17 Indeed, it has been demonstrated that oral administration of OL attenuated the extent and the severity of dextran sulfate sodium-induced acute colitis in mice. 18 Although two previous studies have investigated the anti-tumor effects of OL on mice that spontaneously develop soft tissue sarcomas, 19 and on ovariectomized nude mice with MCF-7 human breast tumor xenografts, 20 no data are available on the in vivo chemopreventive potential of OL on carcinogen-induced colon cancer.
On these bases, in the present study, we have investigated the cancer preventive effect of OL on azoxymethane (AOM)-induced colon carcinogenesis in mice. It is well known that AOM induces preneoplastic lesions in different colon segments, causing increasing severity of crypt dysplasia. On the other hand, no studies have previously investigated the effect of AOM on DNA damage in peripheral blood leukocytes. In this study, we found that the OL-enriched diet prevented the AOM-induced preneoplastic lesions in different colon segments, reducing the severity of crypt dysplasia. In addition, it was observed that AOM induced clear DNA damage in peripheral leukocytes that was efficiently prevented by the OL-enriched diet.
Materials and Methods
Materials, animals, and preparation of diet with OL
AOM was purchased from Sigma-Aldrich (Irvine, United Kingdom). OL was purchased from Extrasynthese (Genay, France). Thirty-four female A/J, 6 week-old mice (Harlan Laboratories, Udine, Italy) were housed in a ventilated temperature-controlled room with a 12 h light/dark cycle and used after 7 days of acclimatization. The animal care and experimental protocols were performed according to the Italian Approved Animal Welfare Assurance. The approval protocol number was DR 33/2011-B, released by “Ministero del Lavoro, della Salute e delle Politiche Sociali” (Rome, Italy) on March 3, 2011. The basal diet (BD) was AIN-93 M (Mucedola, S. Pietro al Natisone, Milano, Italy), whereas the OL diet consisted of BD added with OL at 125 mg/kg of diet (BD-OL). The enriched diet (BD-OL) was produced by Mucedola (S. Pietro al Natisone, Milano, Italy). The concentration of OL (125 mg/kg of diet) was chosen, because it was effective at inhibiting tumor growth and metastases dissemination in ovariectomized nude mice with MCF-7 human breast tumor xenografts. 20 Mice consumed food and water ad libitum.
Experimental design
The experimental protocol of this study is shown in Figure 1A. Mice were randomly divided into three groups: the control group (n = 10), the AOM-BD group (n = 12), and the AOM-BD-OL group (n = 12). The control and AOM-BD groups were fed with the BD, whereas the AOM-BD-OL group was fed with the BD-OL. The AOM-BD and AOM-BD-OL groups were treated with AOM at a dose of 10 mg/kg body weight (prepared in 0.9% saline just before use) via an i.p. injection once a week for 6 weeks; the control group was injected with the same volume of saline not containing AOM. Five mice from each group were sacrificed (overdose of pentobarbital sodium) 7 weeks after the first AOM injection for both histological analysis of dysplasia in the colon crypts and evaluation of DNA damage in peripheral leukocytes. The remaining mice (five in the control group and seven in each AOM-BD and AOM-BD-OL groups) were sacrificed 17 weeks after the first AOM injection for counting the macroscopically observable colon tumors and measuring their dimensions. All animals were weekly monitored for body weight and rectal bleeding.
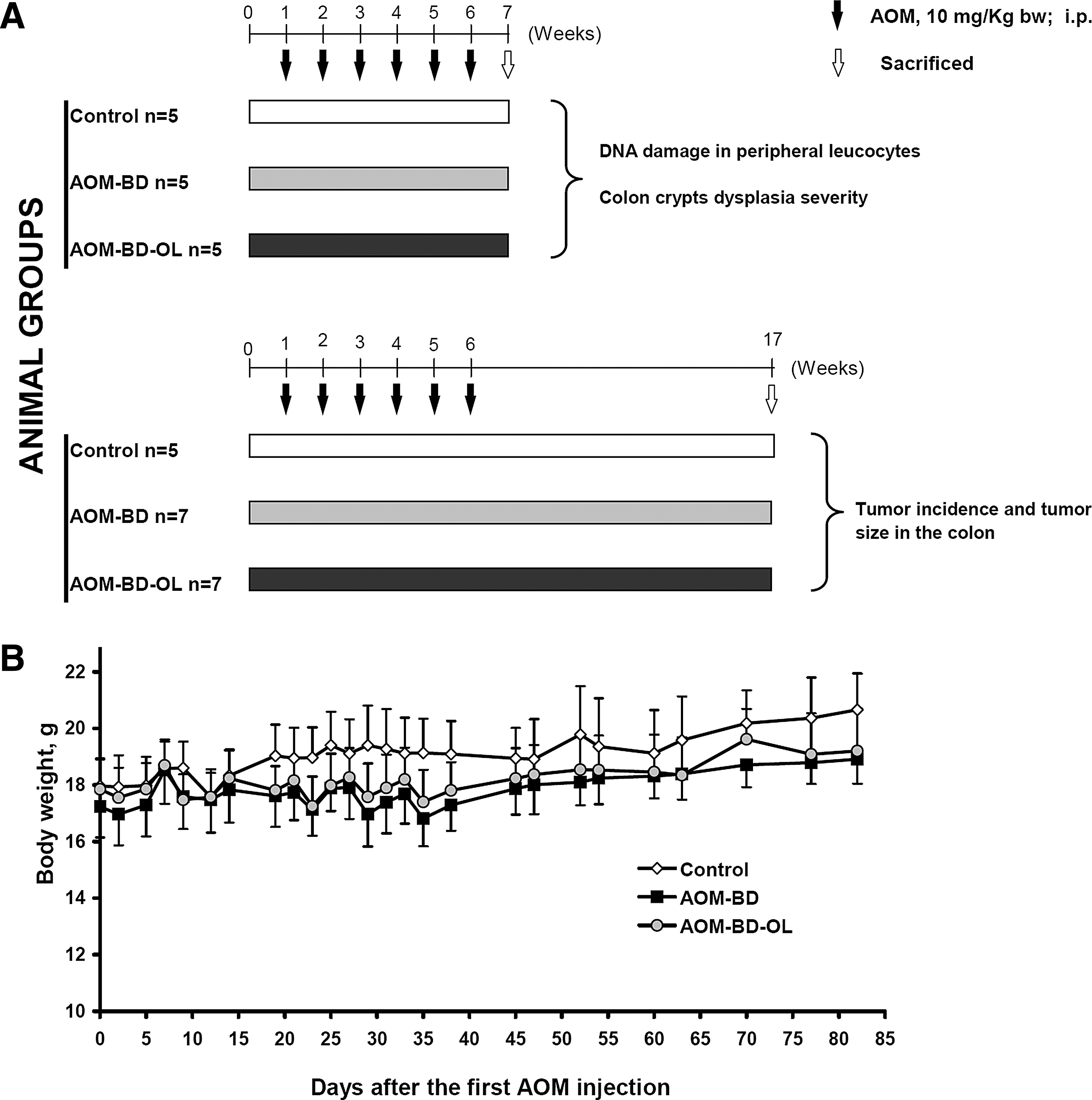
Experimental design for the AOM-induced colon tumorigenesis in A/J mice
Histological analysis of colon dysplasia and tumor measurements
For the histological analysis of colon dysplasia, immediately after sacrifice, the colons were excised starting from ileocecal junction to anal verge and individual segments (proximal, medial, and distal) were obtained. The segments were gently flushed with ice-cold phosphate-buffered saline (PBS) to remove fecal content, opened longitudinally, and fixed flat on filter paper in 10% buffered formalin. Histological examination was performed on paraffin-embedded sections after hematoxylin and eosin (H&E) staining. Colonic mucosa dysplasia (low, medium, and high grade) was classified according to the criteria previously described. 21,22 The percentage of crypt was calculated on the base of at least 100 crypts that were counted for each mouse, colon segment, and experimental condition.
For the tumor measurements, immediately after sacrifice, the entire colon was opened longitudinally along the main axis and processed as for histological analysis. Tumor number was recorded, and tumor diameters was measured with a digital calliper under a dissecting microscope. The tumor volume was calculated with the modified ellipsoidal formula: 1/2 length × width 2 .
DNA damage evaluation, comet assay
Blood samples for the comet assay were collected from intracardiac puncture immediately after sacrifice. To recover the leukocytes, the blood samples were centrifuged at 900 × g for 15 min at 4°C; the supernatant was removed; and the pellet was resuspended in RPMI 1640 medium (Gibco BRL, Life Technologies, Paisley, Scotland). The cells were counted after staining with trypan blue by a Burker chamber. The cell density was adjusted to 1 × 106 cells per mL with RPMI 1640 medium, and aliquots of 50 μL were withdrawn for the DNA damage evaluation by the comet assay (single-cell gel electrophoresis).
Comet assay was performed as previously described. 23 Briefly, the cell samples were centrifuged (200 × g for 5 min); the pellet was resuspended in 75 μL of low-melting-point (LMP) agarose (0.7% in PBS) and distributed onto conventional microscope slides (pre-coated with normal melting-point agarose and dried at 50°C). After the agarose had solidified (4°C for 10 min), a third layer of LMP agarose was applied similarly to the second. The slides were immersed in the lysis solution (2.5 M NaCl, 100 mM Na2EDTA, 10 mM Tris–HCl, pH 10, containing freshly added 1% Triton X-100 and 10% DMSO) for 1 h at 4°C and then placed into a horizontal electrophoresis apparatus that was filled with freshly made buffer (1 mM Na2EDTA, 300 mM NaOH). After 20 min of pre-incubation (unwinding of DNA), the electrophoresis was run for 20 min at a fixed voltage of 25 V (0.83 V/cm) and 300 mA. The slides were washed three times with neutralization buffer (0.4 M Tris–HCl, pH 7.5), stained with 50 μL ethidium bromide (20 μg/mL), and observed by a fluorescence microscope (Zeiss, R.G.). The extension of each comet was analyzed by a computerized image-analysis system (Comet assay II; Perceptive Instruments, Bury Saint Edmunds, United Kingdom) that, among several other parameters, gave the “tail moment” (product of the fraction of DNA in the tail and the mean distance of its migration in the tail). Calculation of the extent of DNA damage was based on the analysis of 100 randomly selected comets from each slide. The DNA damage was expressed as mean of the tail moment, as arbitrary units (A.U.), and as percentage of damaged cells. A.U. were calculated as previously reported. 23
Statistical analysis
All data are presented as mean ± SE. The statistical differences were assessed by Student t-test and chi-square test. A P-value <.05 was considered statistically significant.
Results and Discussion
General observations
AOM is a specific colon carcinogen that acts on both initiation and promotion/progression phases of carcinogenesis. A/J mice are particularly susceptible to AOM-induced colon tumor, resembling the occurrence of non-familial CRC in humans with a preponderance of distal colonic tumors, and sharing many clinical, histological, and molecular features of human CRC. 24 In the present study, we used this animal model to investigate the effect of OL-enriched diet on the early stage of colon carcinogenesis. To highlight the effect of OL on the initiation phase, a subset of animals was sacrificed early (at the end of AOM exposure, 7 weeks) to measure colon crypt dysplasia and DNA damage in peripheral leukocytes (surrogate cells). The remaining animals were followed for up to 17 weeks after the first AOM injection to measure tumor incidence and dimensions (Fig. 1A).
During the carcinogen exposure and dietary intervention, all mice were monitored to point out whether treatments caused any adverse effect. The groups of mice exposed to AOM (AOM-BD and AOM-BD-OL) showed a slight reduction of body weight over time, but this effect for each day of measurement was not statistically significant compared with the control group (Fig. 1B). Furthermore, a significant amount of blood was sporadically observed in the feces of animals treated with AOM; however, no evident differences were observed between AOM-BD and AOM-BD-OL groups (data not shown). This acute toxic effect of AOM has been previously observed and may be a consequence of the young age of animals used in our experiments. 24
Effect of OL on AOM-induced colonic dysplasia and tumors
The effect of OL on the AOM-induced preneoplastic lesions in the colon was assessed 7 weeks after the first AOM injection by H&E staining and light microscopy (Fig. 2A). No aberrant crypts were observed in the colon mucosa of mice given saline alone (control group) (Fig. 2A-left), whereas all animals (100% incidence) treated with the carcinogen (AOM-BD and AOM-BD-OL groups) showed aberrant crypts that were characterized by increased size, thicker epithelial lining, and increased pericryptal zone (Fig. 2A-middle and right, for AOM-BD and AOM-BD-OL, respectively). Aberrant crypts were classified as a function of dysplasia severity into low (hypercellularity of cells with elongated nuclei and focal nuclear stratification), medium (hypercellularity of elongated cells with abundant basophilic cytoplasm, enlarged and vesiculated nuclei, sometimes with prominent nucleoli), and high (nuclear stratification, loss of nuclear polarity, evident structural abnormality of the crypts). 25

Effect of OL on colon crypt dysplasia.
The quantitative results regarding the percentage of normal, low, medium, and high dysplastic crypts in the proximal, medial, and distal segments of the colon (in animals injected with AOM) are shown in Figure 2B–D, respectively. The percentage of crypt was calculated on the base of at least 100 crypts that were counted for each mouse, colon segment, and experimental condition. It is evident that the major effect of AOM was observed in the medial and distal colon segments, where the normal crypts were very few, whereas a significant number of medium and high dysplastic crypts were counted.
The inclusion of OL in the diet caused a preventive effect on AOM-induced preneoplastic lesions in all colon segments as follows: In the proximal segment, OL significantly reduced the low dysplastic crypts and increased the normal crypts (Fig. 2B); in the medium segment, OL reduced the number of both medium and high dysplastic crypts and increased the low dysplasia (Fig. 2C); and in the distal segment, OL caused a decrease of the number of high dysplastic crypts (Fig. 2D). As expected, the incidence of tumors in the AOM-BD group, evidenced 17 weeks after the first AOM injection, increased along the different colon segments (Table 1). In particular, no tumors were observed in the proximal region; whereas four of seven animals (57%) developed a tumor in the medial segment and six of seven (86%) developed a tumor in the distal segment of the colon (Table 1). OL was able to significantly prevent the neoplastic event only in the medial region, reducing the tumor incidence from 57% to 14% (P < .05, chi-square test); whereas no effect was observed in the distal region on both tumor number and volume (Table 1).
Animals were sacrificed 13 weeks after the first AOM injection.
Number of animals with tumor/total number of animals.
P < .05 compared with the AOM-BD.
AOM, azoxymethane; BD, basal diet; OL, oleuropein.
The reasons for the lack of effect on the distal region of the colon are not known, but it may be possible that OL and/or its metabolic degradation products do not reach the distal part of the colon. These results are of particular interest and suggest an evident chemopreventive effect of OL on the colon carcinogenesis. Although in the present study the molecular mechanisms by which OL prevents the CRC have not been investigated, some hypothesis may be suggested. OL may act on both initiation and promotion/progression phases of colon carcinogenesis. For instance, it has been shown that OL aglycone is able to inhibit the tyrosine kinase activity of the epidermal growth factor receptor (EGFR), 26 which is an important regulator of colonocyte growth and differentiation. EGFR is deregulated in up to 50% of human CRC, 27 and colon tumors induced by AOM have shown increased EGFR activity. 28 Therefore, by inhibiting this receptor, OL may reduce the promotion/progression effects of AOM on colon carcinogenesis. Alternatively, OL may inhibit the cancer initiation by preventing the AOM-induced DNA damage and mutation. AOM does not interact directly with DNA but must be activated by cytochrome P450 enzymes to form different metabolites, which, by interacting with DNA, cause gene mutations. 29 OL has been found to inhibit different P450 microsomal activities that could be involved in the AOM activation. 30 Further studies are necessary to clarify these points.
Effect of OL on AOM-induced DNA damage
To further investigate the effect of OL on the cancer initiation phase, the DNA damage in peripheral leukocytes of mice was measured by the comet assay. These surrogate cells were used, because they are easily accessible as single cells, which is a prerequisite for the comet assay. This is a sensitive method that is able to detect both single/double DNA strand breaks and (in the alkaline version) alkali-labile apurinic/apyrimidinic sites on single cells. 31 However, the induction of DNA damage in colonocytes of animals treated with AOM has been previously reported by the comet assay. 32 As shown in Table 2, we observed a significant increment of DNA damage in leukocytes of AOM-injected mice. This effect was evident when DNA damage was expressed as a mean of tail moment, as A.U., and as percentage of damaged cells. Interestingly, the OL-enriched diet almost completely prevented the AOM-induced DNA damage (Table 2).
Animals were sacrificed 7 weeks after the first AOM injection.
In comparison to control; §in comparison to AOM-BD; **,§§ P < .01; ***,§§§ P < .001.
The mechanisms by which AOM induced DNA damage in peripheral leukocytes are actually not known. AOM metabolites may interact with these cells, forming DNA adducts, which, on the action of DNA repair enzymes endonucleases and glycosylases, result in the appearance of either DNA strand breaks or apurinic/apyrimidinic sites, respectively. In this case, OL may prevent the AOM-induced DNA damage by inhibiting either the formation of AOM metabolites or their interaction with leukocytes DNA. On the other hand, it has been shown that AOM is able to induce both systemic oxidative stress and oxidative DNA damage in colonocytes. 33,34 Therefore, oxidative DNA damage may also be induced by AOM exposure in mice leukocytes. In this context, it is important to underline that OL possesses strong anti-oxidant activity that is demonstrated in both in vitro and in vivo systems, 35 and it is also able to prevent the oxidative DNA damage induced in vitro by hydrogen peroxide on human mononuclear cells. 11
In this study, we have evidenced for the first time the potential in vivo preventive activity of OL on a mouse model of CRC. The results indicate that OL was able to prevent AOM-induced carcinogenesis process in mice by reducing the formation of dysplastic crypts in the different segments of the colon and inhibiting the transition from low dysplasia to high dysplasia. OL also reduced the AOM-induced DNA damage in peripheral leukocytes, thus suggesting that it may influence the metabolism and activation of AOM. Further studies are necessary to support this hypothesis. Whatever the mechanisms involved, our data add further evidence to promote human cancer prevention studies using OL-enriched food supplements that are actually available on the market.
Footnotes
Acknowledgments
All work was completed at the University of Perugia, Italy. The authors thank their home institution for the financial support.
Author Disclosure Statement
No competing financial interests exist.