
Abstract
Circulatory markers of low-grade inflammation such as tumor necrosis factor-alpha (TNF-α), interleukin-1 alpha (IL-1α), and interleukin-1 beta (IL-1β) positively correlate with endothelial damage, atheroma formation, cardiovascular disease, and aging. The natural vitamin K2-menaquinone-7 (MK-7) added to the cell culture of human monocyte-derived macrophages (hMDMs) at the same time as toll-like receptor (TLR) agonists did not influence the production of TNF-α. When the cells were pretreated up to 6 h with MK-7 before treatment with TLR agonists, MK-7 did not inhibit significantly the production of TNF-α after the TLR activation. However, 30 h pretreatment of hMDMs with at least 10 μM of MK-7 effectively and dose dependently inhibited the proinflammatory function of hMDMs. Pretreatment of hMDMs with 10 μM of MK-7 for 30 h resulted in 20% inhibition of TNF-α production after lipopolysaccharide (LPS) activation (P < .05) and 43% inhibition after macrophage-activating lipopeptide (MALP) activation (P < .001). Pathogen-associated molecular pattern (PMPP) activation was inhibited by 20% with MK-7 pretreatment; however, this inhibition was not statistically significant. The 30 h pretreatment of a THP-1-differentiated monocyte cell line with MK-7 resulted in a dose-dependent downregulation of TNFα, IL-1α, and IL-1β gene expression as evaluated by RNA semiquantitative reverse transcription polymerase chain reaction (RT-PCR). MK-7 is able to modulate immune and inflammatory reactions in the dose–response inhibition of TNF-α, IL-1α, and IL-1β gene expression and protein production by the healthy hMDMs in vitro.
Introduction
C
Dysregulation of tumor necrosis factor or TNFα activity has been implicated in a wide variety of inflammatory diseases, including rheumatoid arthritis, Crohn's disease, multiple sclerosis, psoriasis, scleroderma, atopic dermatitis, systemic lupus erythematosus (LE), type II diabetes, atherosclerosis, myocardial infarction, osteoporosis, and autoimmune deficiency disease. 6 TNFα upregulates NF-κB, which is critically important as a transcription factor for the expression of TNFα, and it also mediates its biological effects through activation of caspases, activator protein 1 (AP-1 is a transcription factor needed for expression of TNFα), c-jun N-terminal kinase (JNK belongs to mitogen-activated protein kinases), p38 mitogen-activated protein kinases (p38 MAPK), and p44/p42 MAPK. 4 –6 The earlier described changes in levels and activity of biomarkers may signal a chronic inflammatory process in a high-risk group of patients predisposed to or suffering from a degenerative disease and cancer. 7,8
Vitamin K has more than an 80-year-old history since its discovery. In 1929, at the Biochemical Institute of Copenhagen University, experimental studies on chicks fed a very low-fat diet led to the discovery of the existence of vitamin K, which denotes a fat-soluble vitamin that occurs in two biologically active forms, phylloquinone (vitamin K1) and menaquinone (vitamin K2). Vitamin K1 is produced by plants and algae, and vitamin K2 is predominantly of microbial origin and comprises a family of molecules distinguished from K1 by unsaturated side chains of isoprenoid units varying in length from 1 to 14 repeats (hence menaquinone-4, menaquinone-7, etc. or MK-4, MK-7).
The commonly recognized function of vitamin K is a cofactor for γ-glutamyl carboxylase, an endoplasmic enzyme involved in the posttranslational carboxylation (activation) of proteins with glutamic acid (Glu) residues into γ-carboxyglutamate (Gla). With the carboxylation of glutamic acid residues on the protein substrate, a negative chemical group is formed that attracts and binds positively charged calcium cation. Calcium cation is a central element in the physiological role played by the Gla proteins. In contrast, calcium cation cannot be properly utilized by the body without Gla proteins and vitamin K. Vitamin K2, in particular, is responsible for carboxylation and activation of osteocalcin, which is essential for physiology of bone building cells, osteoblasts. Vitamin K2 helps in carboxylation and activation of another protein besides osteocalcin, that is, matrix GLA protein (MGP), important for elasticity and prevention of calcification of blood vessels. Epidemiological findings suggest that the relative risk of mortality due to coronary heart disease is reduced with increased intake of dietary menaquinone (vitamin K2, especially MK-7) but not with intake of phylloquinone (vitamin K1). 9 The intake of menaquinone was also inversely related to all-cause mortality and aortic calcification. 9
The goal of the present study was to test a hypothetical role of vitamin K2 in modulation of the immune and inflammatory biomarkers exemplified by TNFα, IL-1α, and IL-1β in vitro. High-purity natural vitamin K2 (98.4% menaquinone-7, MK-7) was evaluated in vitro for its potential to inhibit gene expression and production of proinflammatory markers by human monocyte-derived macrophages (hMDMs) from two sources (hMDMs and THP-1).
Materials and Methods
Peripheral blood monocyte cell culture
Peripheral blood monocytes (PBMC) were isolated from human blood using a Lymphocyte Separation Medium, a density gradient yielding the fraction highly enriched in monocytes (90% CD14 positive) as described previously.
10
Cells were plated at 3 × 106 cells/well in 24-well plates (Sarstedt, Numbrecht, Germany) in the RPMI 1640 (PAA) culture medium supplemented with 2 mM
Fermentation process-derived natural vitamin K2, 98.4% pure crystalline menaquinone-7 (MK-7), in the 100% trans-form was obtained from NattoPharma ASA (MenaQ7® Crystals). Incubation and preincubation of hMDMs with MK-7 for 6 and 30 h were carried out in 10% human serum supplemented with RPMI 1640 containing MK-7 (0.1, 1, 10, or 100 μM), with medium changed every other day. The 6- and 30-h culture cells were stimulated with toll-like receptor (TLR) agonists, that is, Escherichia coli lipopolysaccharide (LPS) (10 ng/mL), macrophage-activating lipopeptide (MALP) (100 ng/mL), and pathogen-associated molecular patterns (PAMP) (10 ng/mL) in 10% HS RPMI for 6 h. After the 6-h incubation, 500 μL of cell culture supernatants was collected and stored at −80°C until analysis. The level of TNF-α was determined by using commercially available ELISA kits according to the manufacturer's instructions (BD Bioscience, St. Jose, CA).
Human monocyte cell line culture
The THP-1 human monocytes obtained from Bioresource Collection and Research Center (Hsinchu City, Taiwan) were grown in the RPMI 1640 culture medium and 10% fetal bovine serum (GIBCO BRL, Grand Island, NY), supplemented with 1.5 g/L sodium bicarbonate, 4.5 g/L glucose, 10 mM HEPES, 1.0 mM sodium pyruvate, 0.05 mM 2-mercaptoethanol, and 1% penicillin/streptomycin (10,000 units of penicillin/mL and 10 mg streptomycin/mL) in a humidified atmosphere of 95% air and 5% CO2. The experimental protocol for THP-1 monocyte differentiation into macrophages for this study is shown in Figure 1. Briefly, THP-1 monocyte cells (4 × 105 cells) were differentiated into macrophages in 60-mm2 dishes containing 5 mL of growth medium containing 5 ng/mL PMA (phorbol 12-myristate 13-acetate) for 48 h. After THP-1 differentiation and re-fresh growth medium for 12 h, the differentiated macrophages were pretreated with 50 and 100 μM MK-7 for 30 h before stimulated with 100 ng/mL LPS for 6 h.
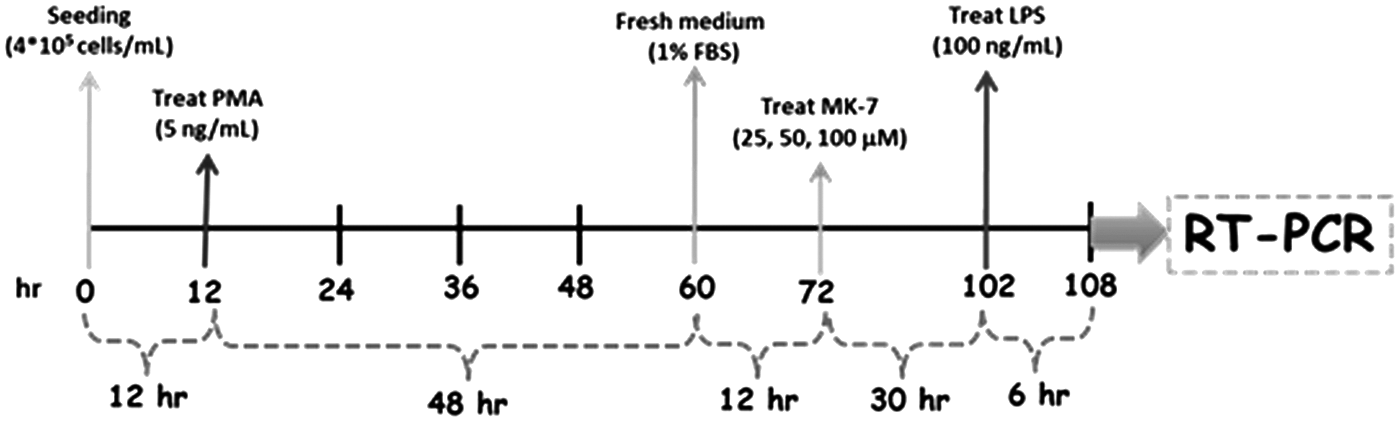
THP-1 monocyte differentiation protocol.
Semiquantitative reverse transcription polymerase chain reaction
Total RNA was isolated from THP-1 differentiated macrophages by using the TRIzol® Reagent according to the manufacturer's instructions (Invitrogen, Carlsbad, CA). A total of 2 μg RNA was transcribed into cDNA using SuperScript II reverse transcriptase (RT) (Invitrogen, Renfrewshire, United Kingdom) in a final volume of 20 μL. RT reactions were performed at 42°C for 60 min and 75°C for 15 min in PCR system 9700 (Applied Biosystems, Singapore, Singapore). The thermal cycle conditions were initiated at 95°C for 10 min and 38 cycles of amplification (94°C for 15 sec, 65°C for 10 sec, and 72°C for 10 sec), followed by extension at 72°C for 1 min. The PCR products were separated by electrophoresis on a 2% agarose gel and visualized by ethidium bromide staining. Amplification of β-actin served as a control for sample loading and integrity. PCR was performed on the cDNA using the following sense and antisense primers: β-actin, forward primer 5′-AAGAGAGGCATCCTCACCCT-3′, reverse primer 5′-TACATGGCTGGGGTGTTGAA-3′; TNF-α, forward primer 5′-ATGTGCTCCTCACCCACACC-3′, reverse primer 5′-GTCGGTCACCCTTCTCCAGCT-3′; IL-1α, forward primer 5′-ACACTATCTCAGCACCACTTGG-3′, reverse primer 5′-ATGAGGTCGGTCTCACTACCTG-3′; IL-1β, forward primer 5′-CAGCTACGAATCTCCGACCAC-3′, reverse primer 5′-GGCAGGGAACCAGCAT CTTC-3′.
Viability assays
After 72 h of incubation of MK-7 (10 and 100 μM) and/or LPS (10 ng/mL) with the hMDMs, the viability of cells was examined by a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reduction assay. The MTT reagent (Sigma) dissolved in serum-free RPMI 1640 (0.5 mg/mL) was added to the cells, followed by incubation at 37°C for 2 h. Any potential formazan crystals formed were dissolved by adding acidified isopropanol solution, and the absorbance of the solution was spectrophotometrically measured at a wavelength of 570 nm. The changes in the amount of formazan formed compared to the untreated control cells indicate the degree of cytotoxicity caused by MK-7 treatment. All assays were performed in triplicate.
Data and statistical analysis
All experiments were performed three times in duplicate. Results were analyzed for statistical significance using the nonparametric Student's t-test. Differences in experimental data were considered significant when P < .05.
Results
The MTT viability assay showed no toxic effects of MK-7 in 10 and 100 μM doses compared to vitamin MK-7-untreated controls and the LPS control. The numerical cell viability was slightly higher in MK-7-treated cells compared to MK-7- untreated and LPS controls (Fig. 2).
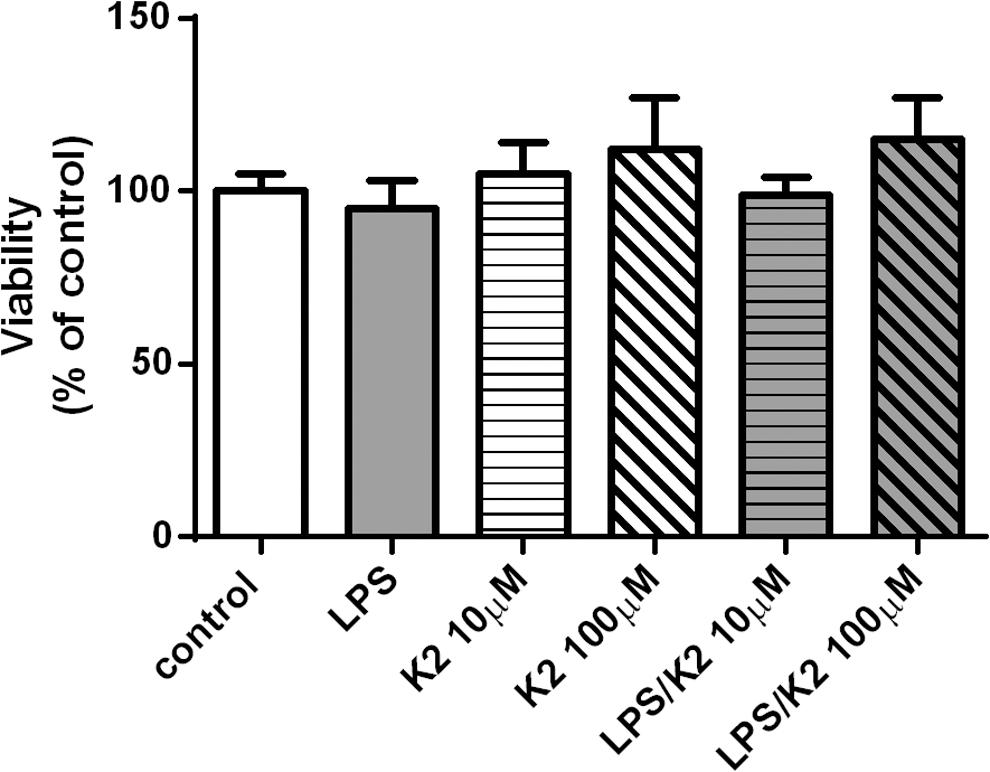
Cell viability assay of hMDMs incubated with K2 (MK-7). Toxicity of vitamin K2 (MK-7) was assessed with MTT assay. hMDMs were incubated with 10 and 100 μM of MK-7 up to 72 h. After the incubation, cells in each well were treated with MTT reagent for 2 h, and the MTT formazan was measured. The figure shows an average and the SD values of three independent cultures. hMDMs, human monocyte-derived macrophages; SD, standard deviation.
TNF-α production by hMDMs was evaluated in vitro in the absence and presence of various concentrations of MK-7 added to the cell culture at the same time as the LPS. MK-7 did not influence the production of proinflammatory mediators (Fig 3A). The next step was to evaluate the potential modulation of the immune and inflammatory biomarkers with MK-7, when the cells were pretreated with MK7 before treatment with LPS. MK-7 in concentrations of 0.1–100 μM for up to 6 h pretreatment did not inhibit significantly the production of TNF-α after the LPS activation (Fig. 3B).

The influence of K2 (MK-7) on production of TNF-α by hMDMs stimulated with LPS, when cultured concurrently or with the pretreatment up to 6 h before the activation with LPS. hMDMs were incubated with 0.1, 1, 10, and 100 μM of vitamin K2 concurrently with the LPS
The 30 h pretreatment with less than 10 μM, that is, 0.1 and 1 μM of Mk-7, was not effective in inhibiting proinflammatory functions of the stimulated hMDMs (Fig. 4A). However, the 30 h pretreatment of hMDMs with at least 10 μM of MK-7 resulted in 20% inhibition of the TNF-α production with LPS activation (P < .05) (Fig. 4B) compared to MK-7-untreated control, and 43% inhibition with MALP activation (P < .001) (Fig. 4C) compared to MK-7-untreated control. The PAMP-activated production of TNF-α was inhibited by 20% with 10 μM MK-7, however, this inhibition was not statistically significant compared to MK-7-untreated control (Fig. 4D). The inhibition of TNF-α production with a 100 μM dose of MK-7 by LPS activation was 63% (Fig. 4B) and with PAMP activation 30% (Fig. 4D) compared to MK-7-untreated controls (P < .001). The MALP activation of TNF-α production was significantly inhibited by 43% with 100 μM compared to MK-7-untreated control (P < .05) (Fig. 4C).

The influence of vitamin K2 (MK-7) on production of TNF-α by hMDMs stimulated with TLR agonists, after 30 h of treatment with MK-7 before the activation with TLR agonists. hMDMs were incubated with 0.1 and 1 μM of vitamin K2 for 30 h before LPS activation
Effects of MK-7 on LPS-induced TNFα, IL-1α, and IL-1β gene expression in THP-1 differentiation macrophage were evaluated in vitro in the absence and presence of various concentrations of menaquinone-7 (MK-7), as described in the Materials and Methods section. The RNA was isolated and analyzed by semiquantitative reverse transcription polymerase chain reaction (RT-PCR). MK-7 added to the THP-1 cell cultures at the same time as the LPS did not influence the gene expression of the three proinflammatory gene markers (data not shown). However, the 30 h pretreatment of the THP-1 with 50 μM and 100 μM MK-7 effectively and significantly (P < .05) inhibited the proinflammatory function of the THP-1 stimulated with 100 ng LPS, as assessed by IL-1α and IL-1β gene expression, and the effect was dose dependent (Fig. 5). Although the 30 h pretreatment with 50 μM of MK-7 was not effective in inhibiting TNFα gene expression in differentiated THP-1, the pretreatment with 100 μM resulted in a statistically significant inhibition of TNFα gene expression (P < .05). In conclusion, the pretreatment of THP-1 differentiated human macrophages with MK-7 resulted in a dose-dependent downregulation of TNFα, IL-1α, and IL-1β gene expression as evaluated by RNA semiquantitative RT-PCR.

Effects of vitamin K2 (MK-7) on LPS-induced TNFα, IL-1α, and IL-1β gene expression in THP-1 differentiation macrophages—RNA was isolated and analyzed by RT-PCR. A total 2 μg of complementary DNA was subjected to RT-PCR with the primers of TNFα, IL-1α, and IL-1β.The values under each lane represent change in gene expression of the bands normalized to β-actin using a densitometer. RT-PCR, reverse transcription polymerase chain reaction.
Discussion
The study showed for the first time that the MK-7 form of vitamin K2 is able to modulate the immune and inflammatory reactions as evidenced by the dose-dependent inhibition of TNF-α, IL-1α, and IL-1β gene expression and production of TNF-α, IL-1α, and IL-1β proteins by the healthy hMDMs in vitro. Highly selective inhibitors of inflammatory reactions, for example, COX-2 inhibitors, may effectively reduce inflammation in both acute and chronic inflammatory conditions, but they work in a very selective way that may be counterproductive to the purpose of treatment. Recent research reveals that selective COX-2 inhibitor drugs may induce metabolic imbalances that can result in the overproduction of toxic cytokines: TNF-α and certain interleukins that are involved in the inflammatory process. 11
The TNF-α inhibitors such as infliximab and adalimumab (Humira) are approved for treatment of rheumatoid arthritis, psoriasis, psoriatic arthritis, Crohn's disease, ankylosing spondylitis and juvenile idiopathic arthritis, and may also have potential as cardioprotective drugs in the treatment of CVDs. 6 The cardiovascular potential of TNF-α therapy was evaluated in patients treated for arthropathies, a group of patients known to develop premature arterial stiffening and arteriosclerosis. 12 One-year treatment with the TNF-α antagonist improved over time the pulse wave velocity (PWV), slowed down progression of carotid intima media thickness, and lowered levels of the circulating proinflammatory protein, calprotectin. 12 However, the use of TNF-α inhibitors, such as Humira, has been associated with serious side effects due to induction of autoimmune diseases, including systemic LE, vasculitis, and sarcoidosis. 12 Patients with psoriasis treated with Humira developed renal failure and seropositive LE markers without classic LE nephritis. 11
In view of the effect of vitamin K2 on the circulatory system by activating the MGP, which prevents calcium deposits in the arteries and is one of the most potent arterial calcification inhibitors known, the cardiovascular potential of vitamin K2 is increasingly recognized. This emerging trend in vitamin K (K2's MK-7 variety in particular) supplemental use in cardiovascular conditions has been further supported by a prospective randomized study in patients with chronic kidney disease (CKD), where the cardiovascular effects of oral administration of vitamin K2 (MK-7) plus vitamin D or vitamin D alone were evaluated. 13 In this 6-month study, the progression of coronary artery calcification index and common carotid intima media thickness detected with computerized tomography showed a slower progression of calcification in the vitamin K2/vitamin D group than in the vitamin D-alone group of patients. 13 In a rodent experimental model of CKD, K2 supplementation attenuated neointimal hyperplasia and calcification in arteriovenous fistula in healthy rats and rats with CKD, as well as significantly enhanced MGP carboxylation in control rats and rats with CKD. 14
In a double-blind randomized clinical trial evaluating the results of a 3-year regular intake of natural MK-7 in a 180 μg daily dose by a group of 244 healthy postmenopausal 55–65-year-old Dutch women, there was no report of serious objective or subjective side effects of this form of vitamin K2. 15 In addition, the study showed statistically significant benefits of MK-7 supplementation, compared to the placebo group, in preventing age-related stiffening of arteries. After 3 years of MK-7 supplementation, PWV index cfPWV and the Stiffness Index β significantly decreased in the study group, whereas distension, compliance, distensibility, Young's Modulus, and the local carotid PWV (cPWV) improved in women having a baseline Stiffness Index β above the median of 10.8. MK-7 supplementation significantly decreased circulating dephospho-uncarboxylated matrix Gla-protein (dp-ucMGP) by 50% compared to placebo, but did not influence the markers for inflammation, that is, IL-6, high-sensitive C-reactive protein (hsCRP), and TNF-α. In conclusion, the long-term use of MK-7 supplements reduces arterial stiffness in healthy postmenopausal women, especially in women having a high arterial stiffness.
The results of our study indicating lowering gene expression for IL-1α and IL-1β by MK-7 supplementation support the literature-reported role of both interleukins in the development of CVD. In humans, IL-1β was found in greater concentrations in atherosclerotic human coronary arteries compared to normal coronary arteries, and it has been suggested that IL-1β may be a novel therapeutic target for anti-inflammatory therapy in atherosclerosis, such as coronary artery disease. Currently, the Canakinumab Anti-inflammatory Thrombosis Study (a monoclonal antibody directed against IL-1β) is underway to evaluate the inflammatory hypothesis of CVD. 16
The nutritional status of MK-7 and its cardiovascular protective role may be compromised, in addition to well-recognized vitamin K2 nutritional insufficiency, 17 by interaction with statin drugs. 18 Based on a recent review article, statins may inhibit the synthesis of vitamin K2 which is, as previously discussed, the cofactor for matrix Gla-protein activation protecting arteries from calcification. In view of that finding, the authors of the report propose that current statin treatment guidelines should be critically reevaluated. 18
The epidemiological studies suggest that most reasonably healthy adults may be subclinically vitamin K2 deficient—which results in K2-dependent proteins not carboxylated and biologically inactive. 17 The high circulating concentrations of uncarboxylated MGP (ucMGP) have been shown to predict CVD mortality. 17 Thus, although the intake of vitamin K in Western countries is sufficient for maintaining homeostasis, it might be insufficient to maintain posttranslational modification of the vitamin K-dependent proteins and thus insufficient to prevent not only CVD but also other chronic degenerative conditions, including osteoporosis and cancer. 9,17
Conclusion
The MK-7 form of vitamin K2 is able to dose dependently inhibit TNF-α, IL-1α, and IL-1β gene expression and protein production by healthy hMDMs in vitro. In view of these encouraging findings on the anti-inflammatory properties of MK-7, we plan to test additional biomarkers of immune and inflammatory response in this form of vitamin K2 in vitro. This work may elucidate the anti-inflammatory mechanism of vitamin K2 and establish the potential biomarker targets in clinical testing of the role of MK-7 in the cardiovascular health as well as other chronic degenerative conditions.
Footnotes
Acknowledgment
Supported by a grant from NattoPharma ASA, Oslo, Norway.
Author Disclosure Statement
No competing financial interests exist.