
Abstract
We reported previously that hydroxylated polymethoxyflavones (HPMFs) effectively suppressed obesity in high-fat-induced mouse. In this study, we further investigated the molecular mechanism of action of 5-hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone (5-OH-HxMF), one of major HPMFs in orange peel. Treatment of 5-OH-HxMF effectively inhibited lipid accumulation by 55–60% in a dose-dependent manner. The 5-OH-HxMF attenuated adipogenesis through downregulating adipogenesis-related transcription factors such as peroxisome proliferator-activated receptor gamma (PPARγ) and CCAAT/enhancer-binding proteins (C/EBPs), as well as downstream target fatty acid synthase and acetyl-CoA carboxylase (ACC). 5-OH-HxMF activated adenosine monophosphate-activated protein kinase signaling and silent mating type information regulation 1 (SIRTUIN 1 or SIRT1) in 3T3-L1 adipocytes to decrease lipid accumulation. In addition, the inhibition rate of lipid accumulation was compared between 5-OH-HxMF and 3,5,6,7,8,3′,4′-heptamethoxyflavone (HpMF). 5-OH-HxMF inhibited lipid accumulation 15–20% more than HpMF did, indicating that hydroxyl group at position 5 can be a key factor in the suppression of adipogenesis.
Introduction
O
Our previous study indicated that crude extract of orange peel rich in hydroxylated polymethoxyflavones (HPMFs) could efficiently suppress the accumulation of lipid in adipocytes. 6 They can downregulate PPARγ, SREBP-1, as well as fatty acid synthase (FAS) and ACC. AMPK was activated using orange peel extract. To determine the major bioactive compounds, the composition of the orange peel was analyzed indicating that a group of HPMFs had a great potential to prevent obesity. HPMFs, which abundantly occur in the peel of sweet oranges (Citrus sinensis), have shown various biological activities, such as anti-inflammatory and antitumor activities. 7 5-hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone (5-OH-HxMF) (Fig. 1a), one of the major HPMFs, has been reported to have better activity than the rest of HPMFs to induce apoptosis in human leukemia cells. 8 Because of the concern for drug safety, more consumption of natural bioactive compounds in medicinal foods is beneficial to the management of obesity, but many issues require further investigation, including the dosage, bioavailability effectiveness, and safety in clinical studies. 9,10 All these studies should start from the single compound instead of a mixture. It is necessary to know if a single HPMF has the activity of antiadipogenesis/obesity. Therefore, the first goal of this short communication was to study the anti-adipogenic activity of a single HPMF, 5-OH-HxMF through transcription factors. Different HPMF has different bioactivity. To understand this, 3,5,6,7,8,3′,4′-heptamethoxyflavone (HpMF) was used to compare with 5-OH-HxMF.
Materials and Methods
Chemicals
Dulbecco's modified Eagle's medium (DMEM), fetal calf serum, and fetal bovine serum (FBS) were obtained from Gibco BRL (Grand Island, NY, USA). Insulin, isobutylmethylxanthine, and dexamethasone pioglitazone were purchased from Sigma Chemical Co. (St. Louis, MO, USA). The antibodies for CEBPα, PPARγ, ACC, FAS, p-AMPKα, SIRT1 (silent mating type information regulation 1), and FOXO1 were purchased from Cell Signaling Technology (Beverly, MA, USA). 5-OH-HxMF and HpMF were purified in-house using Prep-HPLC (high-performance liquid chromatography) method described previously. 8 The purity determined by an HPLC was higher than 99%.
3T3-L1 cell culture and differentiation
Mouse 3T3-L1 preadipocytes were purchased from the American Type Culture Collection (Rockville, MD, USA). They were grown and differentiated in the same conditions as in the previous study. 6 After incubation in a differentiation medium for 2 days, cells were applied with DMEM containing 10% FBS and insulin (1.7 μM) in the presence or absence of 5-OH-HxMF, which was replaced every 2 days. The cells were harvested after 8 days (at day 10) for the following test.
Cytotoxicity assay
The effect of 5-OH-HxMF on cell viability and toxicity was tested using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and Trypan Blue analysis, respectively. Details of MTT assay followed the procedure set up in the previous study. 11 After differentiation and 5-OH-HxMF treatment at day 10, the cells were harvested and determined using Trypan Blue exclusion and microscopy examination for their cytotoxicity.
Oil Red O staining
At the end of differentiation, the cells were washed by phosphate-buffered saline and applied with 0.5% Oil Red O dye. Conditions were consistent with the previous study. 6 Excess Oil Red O dye was washed with distilled water and then dried. The stained lipid droplets within cells were visualized using light microscope and photographed at 200 × magnification. Quantification of lipid accumulation was preceded at the absorbance of 520 nm using ImageJ imaging software (National Institutes of Health, Bethesda, MD, USA).
Western blot analysis
The total proteins of differentiated 3T3-L1 adipocytes were extracted with gold lysis buffer and centrifuged. The lysis buffer formula and procedure followed the published work. 11 The samples (50 μg of protein) were treated with 5 × sample buffer solution for sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis. The sample buffer solution contained 0.3 M Tris-HCl (pH 6.8), 25% 2-mercaptoethanol, 12% SDS, 25 mM ethylenediaminetetraacetic acid (EDTA), 20% glycerol, and 0.1% bromophenol blue. Proteins on the gel were electrotransferred onto an immobile membrane (polyvinylidene difluoride [PVDF]; Millipore Corp., Bedford, MA, USA) with transfer buffer composed of 25 mM Tris-HCl (pH 8.9), 192 mM glycine, and 20% methanol. The membranes were blocked with blocking solution containing 20 mM Tris-HCl and then immunoblotted with primary antibodies, including C/EBPα, PPAR γ, ACC, FAS, p-APMK, SIRT1, FOXO1, and β-actin. The transferred proteins were visualized with an Enhanced Chemiluminescence Detection Kit (Amersham Pharmacia Biotech, Buckinghamshire, United Kingdom). 11
Statistical analysis
Data are presented as mean ± standard error for the indicated number of independently performed experiments. Comparisons of statistical significance between groups were made by one-way Student's t-test or one-way analysis of variance (ANOVA). A P-value <.05 was considered statistically significant.
Results
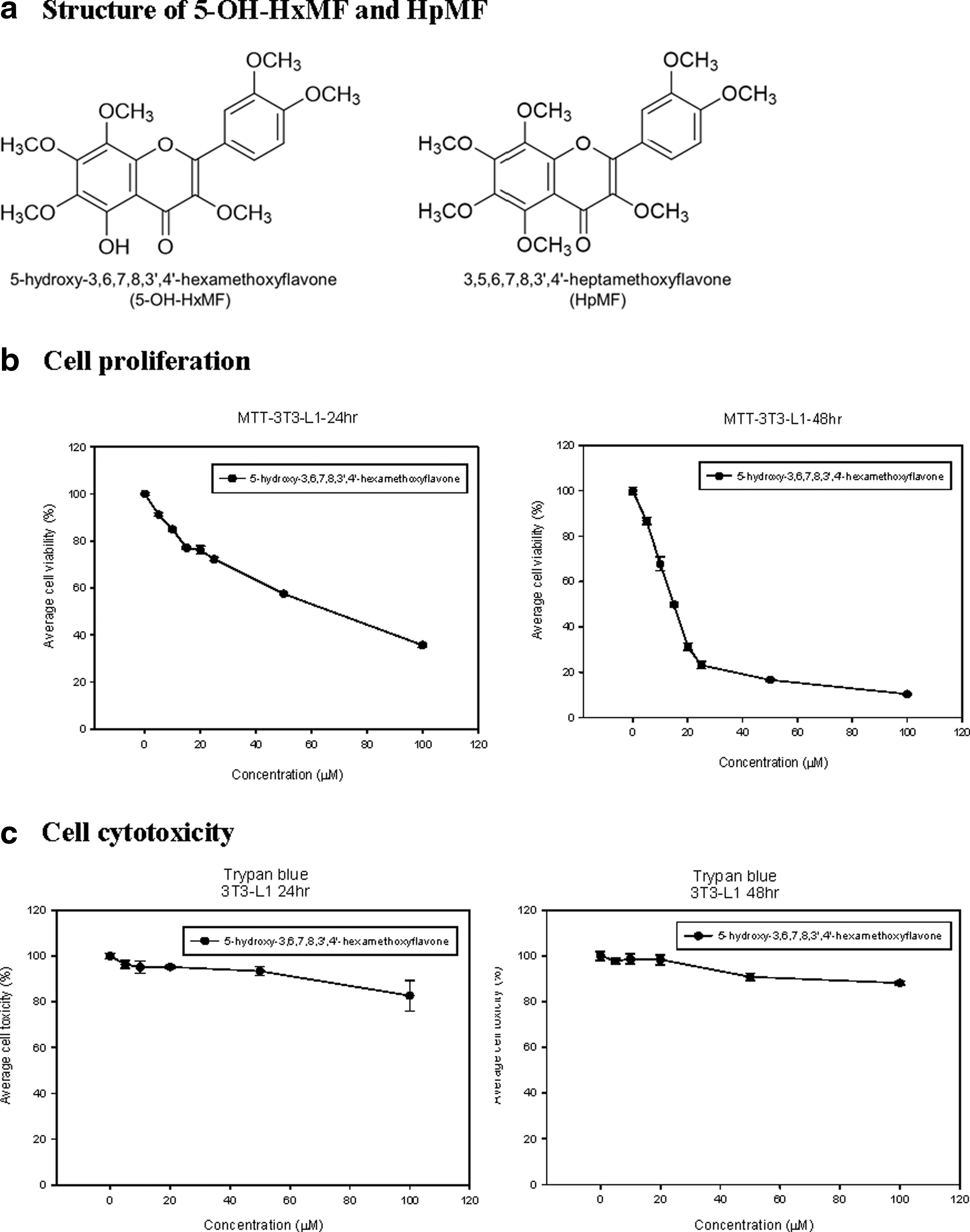
Effect of 5-OH-HxMF on 3T3-L1 cell viability and cytotoxicity
Different concentrations of 5-OH-HxMF (0, 5, 10, 15, 20, 25, 50, and 100 μM) were applied separately to 3T3-L1 cell line to investigate the cell viability using MTT tetrazolium assay. Cell viability was significantly decreased with a 48-h treatment. When cells were treated with 20 μM 5-OH-HxMF for 24 h, the cell viability was kept around 80% (Fig. 1b). Cell toxicity was also determined using Trypan Blue assay. 5-OH-HxMF at a concentration of 100 μM showed no significant cytotoxicity (<10%) to 3T3-L1 cells with a 24-h treatment (Fig. 1c). Therefore, 20 μM of 5-OH-HxMF was the maximum concentration used for the following study.
Effect of 5-OH-HxMF on intracellular lipid accumulation and structure-related activity
3T3-L1 preadipocytes were incubated with differentiation medium and insulin with 5-OH-HxMF at doses of 0, 5, 10, and 20 μM, separately for 8 days. Oil Red O staining was used to measure the lipid accumulation after 8 days of cell differentiation (Fig. 2). Without application of 5-OH-HxMF, a lot of lipid accumulation can be seen compared with control. However, when applied with 5-OH-HxMF, 3T3-L1 cells showed a decreased lipid accumulation, and 5-OH-HxMF reduced the lipid accumulation in a dose-dependence manner. For example, Figure 2a showed that under the microscope, a bit of lipid accumulation can be observed with the treatment of 5-OH-HxMF at 10 μM, but nearly no lipid accumulation was obtained at the concentration of 20 μM. Through the absorbance, 20 μM of 5-OH-HxMF could reduce the lipid accumulation by 60% (Fig. 2b). To study the structure-related activity, HpMF was used to treat 3T3-L1 cells. The microscope data showed more lipid accumulation of HpMF treatment than that of 5-OH-HxMF treatment (Fig. 2a). At a dose of 20 μM, HpMF decreased the lipid accumulation by nearly 40%, whereas 5-OH-HxMF showed a decreasing rate of 60% (Fig. 2b).
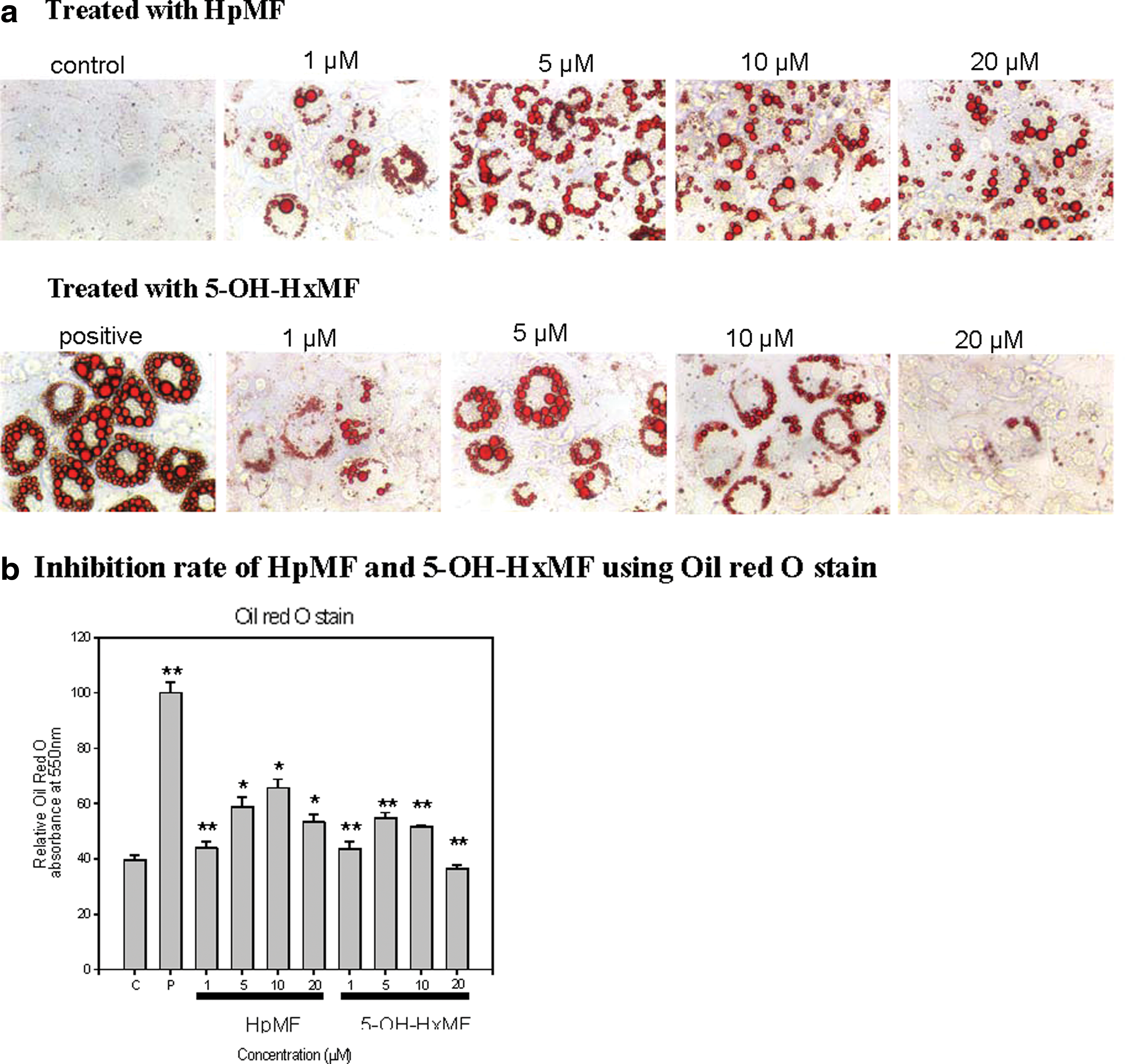
The effect of different doses of HpMF and 5-OH-HxMF on inhibition of 3T3-L1 preadipocytes differentiation.
5-OH-HxMF inhibited adipogenic transcription factors and proteins, but activated AMPK
Adipogenesis is regulated by transcription factors C/EBPα and PPARγ of which activation can lead to differentiation through inducing FAS and ACC. The control group, referred as 3T3-L1 preadipocytes, incubated without differentiation medium, whereas vice versa for positive group. When 5-OH-HxMF and differentiation medium applied to preadipocytes, the expression of C/EBPα, PPARγ1, and PPARγ2 was regulated in a dose-dependent manner. The higher the concentration of 5-OH-HxMF, the more the inhibition expression of the transcription factors (Fig. 3a). Termination of differentiation can also be controlled by the protein expression of FAS and ACC. Application of 5-OH-HxMF induced a dose-dependent downregulation of ACC and FAS, which corresponded to the transcription factors (Fig. 3b).

Effect of 5-OH-PMFs on
AMPK is a regulator to keep energy homeostasis through inhibiting lipid and glucose synthesis. Phosphorylation of AMPK was upregulated in a dose-dependent manner when 3T3-L1 adipocytes were treated with 5-OH-HxMF (Fig. 3c). Previous studies showed that activation of AMPK could inhibit the enzyme activity of ACC. 6
Regulation of SIRT1 and FoxO1 by 5-OH-HxMF
SIRT1 has been reported to regulate adipogenesis. 12 Adipocyte differentiation has been considered to relate to the upregulation of SIRT1. 13 Figure 4 indicated that protein level of SIRT1 increased with the increased level of 5-OH-HxMF suggesting the upregulation of SIRT1. Meanwhile, the expression of FoxO1 also increased in a dose-dependent manner, and upregulation of FoxO1 was related to inhibit cell differentiation. 14

Effect of 5-OH-PMFs on SIRT1, FOXO1, and β-actin protein expressions in 3T3-L1 adipocytes. Western blot analysis of SIRT1, FOXO1, and β-actin protein levels. SIRT1, silent mating type information regulation 1.
Discussion
Obesity is considered a serious health problem in the world. World Health Organization (WHO) defined obesity as a phenotypic manifestation of abnormal or excessive fat accumulation that alters health and increases mortality. Enhanced adipogenesis due to energy imbalance and storage of triacylglycerols in adipocytes can increase adipose tissue mass and induce obesity. Therefore, the adipocyte is believed to be an antiobesity target through regulating adipogenesis. Because of the concern for drug safety, numerous researches have been focused on plant or herb extracts with less detrimental effects. In our previous study, HPMFs from orange peel extract can effectively decrease the accumulation of lipid and preadipocyte differentiation through downregulation of adipogenesis-related transcription factors, such as PPARα and C/EBPs, as well as FAS and ACC. 6 However, HPMFs is a mixture of many polymethoxyflavones such as nobiletin, 3,5,6,7,8,3′,4′-heptamethoxyflavone, tangeritin, 5-hydroxy-6,7,3′,4-tetramethoxyflavone, 5-hydroxy-6,7,8,3′,4′-pentamethoxyflavone, 5-hydroxy-3,6,7,3′,4′-pentamethoxyflavone, 5-hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone, 5-hydroxy-6,7,4′-trimethoxyflavone, and 5-hydroxy-6,7,8,4′-tetramethoxyflavone. 9 Therefore, it was hard to understand the effect of one single compound and its mechanism. Understanding the molecular mechanism of one single compound is a key for the future application in obesity intervention. In addition, 5-OH-HxMF has been shown a better activity in induction of apoptosis. 8 Thus, in this short communication, 5-OH-HxMF was investigated for its antiadipogenesis.
The activity of antiadipogenesis was compared between nobiletin and a mixture of polymethoxyflavones (PMFs) from orange peel (data not shown here). Nobiletin showed less activity than PMFs indicating either number of methoxy group or hydroxyl group might be the factor. Therefore, 5-OH-HxMF and HpMF were used. HpMF has seven methoxylated groups, whereas 5-OH-HxMF has six methoxylated groups and a hydroxyl group at position 5. 5-OH-HxMF showed 15–20% more inhibition rate of lipid accumulation than HpMF did, suggesting that the hydroxyl group can affect antiadipogenesis. Considering the structures of nobiletin, 5-OH-HxMF and HpMF, we predicted that hydroxyl group at position 5 can be the reason for higher activity of antiadipogenesis.
Accordingly, the previous study on 5-OH-HxMF with a sequential concentration was applied to 3T3-L1 preadipocytes, separately. 5-OH-HxMF at the concentration of 20 μM can keep the cell viability and reduce lipid accumulation in a dose-dependent manner using Oil Red O staining. PPARγ is a key regulator in the development of adipogenesis. HPMFs were shown to downregulate PPARγ2 to suppress adipocyte differentiation, but had no apparent effect on PPARγ1. 6 However, for the pure compound, 5-OH-HxMF demonstrated the downregulation not only of PPARγ2, but also of PPARγ1. Activation of PPARγ increases the expression of C/EBPs and vice versa. 5-OH-HxMF, therefore, should also inhibit C/EBPs expression, which agreed with our results. Activation of PPARγ and C/EBP can indirectly activate FAS and ACC, which also are therapeutic targets for obesity. 5-OH-HxMF, the same as HPMFs, can downregulate the protein expression of FAS and ACC in a dose-dependent manner. In addition, AMPK, which controls cellular metabolism, is also involved in the modulation of adipose metabolism through regulating glucose uptake, lipid oxidation, and adipokine secretion. In the previous study of the HPMFs mixture, HPMFs significantly induced phosphorylation of AMPK, 6 which can be observed with the treatment of pure 5-OH-HxMF. Therefore, based on the discussion above, single compound 5-OH-HxMF can be the major compound in HPMFs to suppress 3T3-L1 adipocyte differentiation and lipid accumulation through downregulating the adipocyte-related transcription factors PPARγ and C/EBPs, as well as activating AMPK signaling (Fig. 5). Animal study will be applied for 5-OH-HxMF in the future.
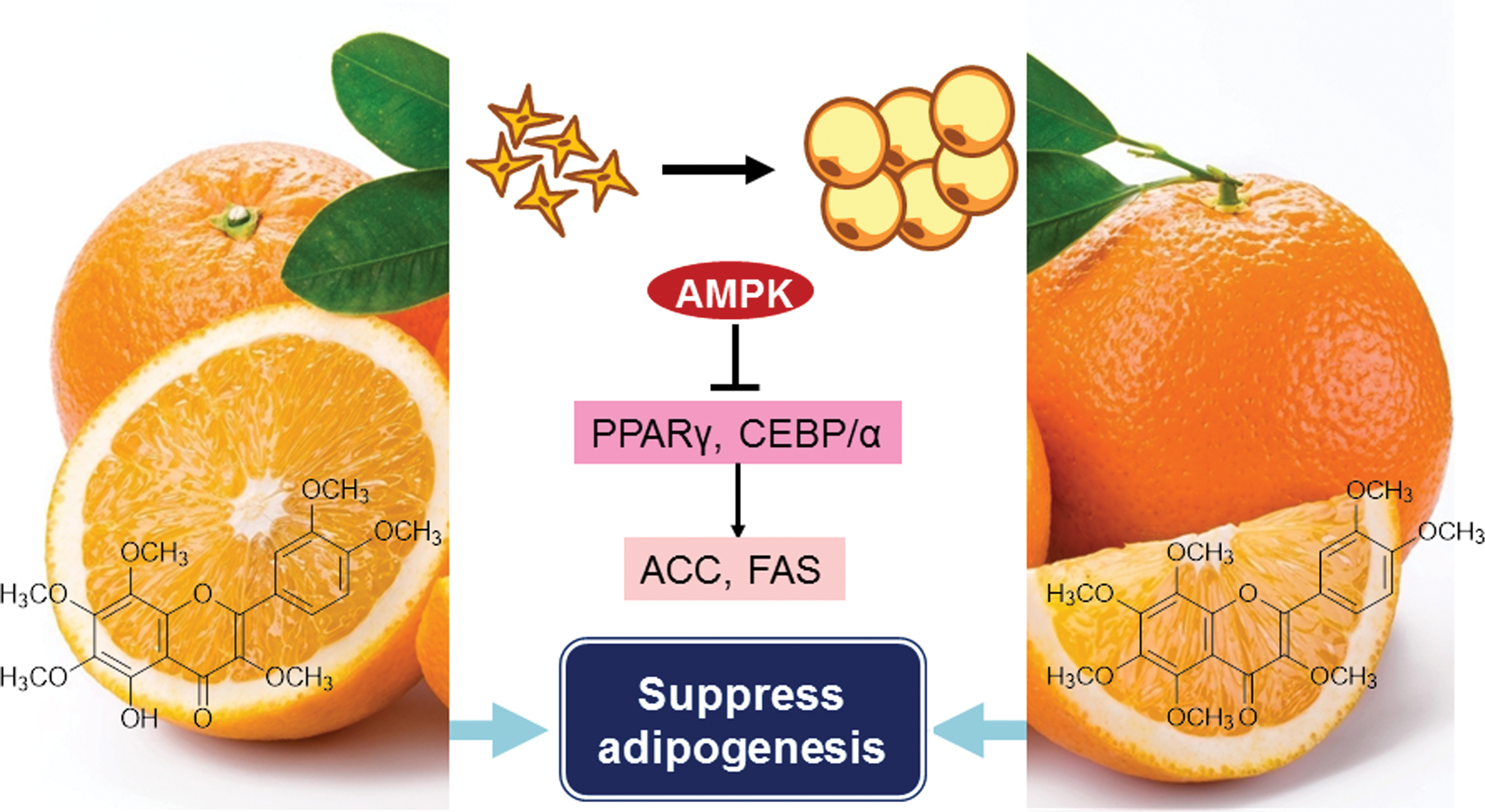
Possible mechanism of 5-OH-HxMF and HpMF on suppression of 3T3-L1 adipocytes differentiation. Color images available online at
Understanding the molecular mechanism and network implicated in the pathogenesis of obesity can provide potential opportunities for its prevention and intervention. In the previous study, the effect of HPMFs on sirtuins, particularly SIRT 1, has not been studied. However, recent increasing interest in the role of sirtuins is related to their biological functions, such as longevity, inflammation, and cellular metabolism. NAD+-dependent deacetylase SIRT1 can reprogram gene expression regulating nucleosome positioning and chromatin structure. It has an extensive regulatory role in physiological activities because of its capability to freely translocate among subcellular compartments. SIRT1 can directly deacetylate NFκB p65 and p53 to regulate inflammation and cancer. 15 Upregulation of SIRT1 can inhibit adipogenesis through suppressing PPARγ and C/EBPα. 12 Protein expression of SIRT1 increased with the treatment of 5-OH-HxMF in a dose-dependent manner. Meanwhile, downregulation of PPARγ and C/EBPα was also observed. In addition, overexpression of SIRT1 induces lipolysis by deacetylation of FoxO1 in adipocytes. Studies indicated that insulin signaling can increase adipocyte differentiation through activation of PI3K/Akt and inactivation of FoxOs. 14 In our study, application of 5-OH-HxMF caused the upregulation of FoxO1, but the detail of this mechanism still needs further study in the future.
This study is the first time to show 5-OH-HxMF increasing the expression of SIRT1 to attenuate adipogenesis. This study showed that the hydroxyl group at position 5 could be the reason for better activity of antiadipogenesis suggesting that orange peel containing higher levels of HPMFs with hydroxyl group at position 5 may have the potential to develop into the medicinal food for antiadipogenesis/obesity.
Footnotes
Acknowledgments
This study was supported by the National Taiwan University NTU-104R7777, and the Ministry of Science and Technology 101-2628-B-022-001-MY4 and 102-2628-B-002-053-MY3.
Author Disclosure Statement
No competing financial interests exist.