
Abstract
Apium graveolens has been shown to inhibit the growth of a variety of cancer tissues. In this study, we investigated the anticancer effect of A. graveolens on the human prostatic carcinoma cell line LNCaP. LNCaP cells were treated with increasing concentrations of an ethanolic extract of A. graveolens ranging from 1000 to 3000 μg/mL, and viability was determined after 24 and 48 h using the XTT cell proliferation assay. The levels of cleaved poly (ADP-ribose) polymerase (PARP), one of the best biomarkers of apoptosis, were analyzed. Finally, quantitative gene expression analysis of vascular endothelial growth factor (VEGF), a critical mediator of angiogenesis, was performed using real-time reverse transcription–polymerase chain reaction. A. graveolens extract inhibited cell viability in both a time- and dose-dependent manner. Data from cleaved PARP assays suggested that A. graveolens caused induction of apoptosis in these cells. Treatment of cells with A. graveolens also resulted in downregulation of VEGF expression. This study showed that the antiproliferative effect exerted by an ethanolic extract of A. graveolens is triggered by induction of apoptosis. We also demonstrated that VEGF expression was downregulated by treatment with A. graveolens extract.
Introduction
T
During the last decade, apoptotic induction and inhibition of tumor angiogenesis have been new targets for anticancer drug discovery. 9,10 Cytostatic drugs that inhibit tumor cell proliferation and growth with low toxicity should be of interest for the research and treatment of cancer. A. graveolens and substances derived from it have been shown to inhibit growth and to induce apoptosis in various cancer tissues. They induce apoptosis of the human colon cancer cell line HCT116, 11 Dalton's lymphoma ascites cells, 12 the human pancreatic cancer cell lines BxPC-3 and PANC-1, 13 the human stomach cancer cell line BGC-823, 14 the mouse neuroblastoma cell line neuro-2a, 15 the human cervical cancer cell line HeLa, 16 and the breast cancer cell line MCF-7. 17 However, no further work on apoptotic effect on human prostate cancer cell line LNCap has been reported.
Angiogenesis, which is initiated by particular tumor microenvironmental conditions and several angiogenic factors, including vascular endothelial growth factor (VEGF), basic fibroblast growth factor, and platelet-derived growth factor, plays a principal crucial role during tumor development and metastasis. VEGF is the most important proangiogenic factor, and the level of VEGF is an important prognostic marker of tumor angiogenesis. 18 Thus, many antitumor and antiangiogenic strategies targeting VEGF have been developed for inhibiting cancer angiogenesis, offering insights into the successful treatment of solid cancers. However, the side effects of anti-VEGF agents such as hypertension, bleeding, and gastrointestinal perforation, constrict their chronic use. 19 Considering these issues, there have recently been extensive studies done on plant-derived compounds and extracts that show potent antiangiogenic activity with the potential of overcoming the adverse effects of chemical compounds on human health.
In this study, we investigated the effects of A. graveolens extracts on cell proliferation, apoptosis, and angiogenesis in human prostate carcinoma LNCaP cells.
Materials and Methods
Cell line and culture conditions
The LNCaP cell line was obtained from the Yeditepe University, Faculty of Medicine, Department of Physiology (Istanbul, Turkey). The cell line was cultured in Roswell Park Memorial Institute (RPMI) medium 1640 supplemented with 10% fetal bovine serum, 100 U/mL penicillin G, and 100 mg/mL streptomycin and kept at 37°C in a humidified incubator with an atmosphere of 5% CO2 and 95% air.
Preparation of ethanolic extract of A. graveolens
The fresh, whole celery (root plus stalks plus leaves) used in this study were procured from the local vegetable market. Plant processing was done in the Anadolu University Medicinal Plants, Drugs and Scientific Research Center. The whole plant was chopped into small pieces and dried in the shade. The dried plant was crushed to coarse powder using a hand mill and sieved. Coarse powder was successively extracted in a soxhlet apparatus with 70% ethanol for 12 h. The ethanol extracts were concentrated by evaporating under vacuum. The yield of the ethanol extract was 44.45%. The ethanol extracts were freshly dissolved in dimethyl sulfoxide (DMSO) just before administration.
Cell viability assay of LNCaP cells using XTT assay
Cells were seeded at 1 × 104 cells per well in complete culture medium containing various concentrations of ethanolic extract of A. graveolens (0, 1000, 1500, 2000, 2500, 3000 μg/mL, and 1% DMSO vehicle control). The cells were incubated for 24 and 48 h, respectively, to determine cytotoxic and apoptotic effects. Each concentration of ethanolic extract of A. graveolens was incubated in four wells to identify the most efficient doses and incubation periods. Cytotoxic activity was measured using a colorimetric assay (the XTT Cell Proliferation Kit; Biological Industries), which measures the reduction of a tetrazolium component, XTT (sodium 3′-[1-phenyl-aminocarbonyl)-3,4-tetrazolium] bis(4-methoxy-6-nitro)benzenesulfonic acid) into a soluble formazan product by the mitochondria of viable cells. Briefly, cells were cultured in 96-well plates containing 200 μL of complete medium. One hundred microliters of XTT solution was added into the wells and plates were incubated for additional 3 h at 37°C. The controls included native cells and medium only. The optical density of 96-well plates was measured at 450 nm in a microplate enzyme-linked immunosorbent assay (ELISA) reader (Trinity Biotech PLC, Bray CO., Wicklow, Ireland). The percentage of cytotoxicity was calculated with the formula: percentage cytotoxicity (cell death) = [1 − (absorbance of experimental wells/absorbance of control wells)] × 100%.
Apoptosis assay: cleaved PARP (asp 214) sandwich ELISA
Cleaved poly (ADP-ribose) polymerase (PARP) was assayed using the PathScan Cleaved PARP (asp 214) Sandwich ELISA Kit (Cell Signaling Technology, Danvers, MA, USA). Cells were seeded at 1 × 104 cells per well in a complete culture medium containing various concentrations of ethanolic extract of A. graveolens (0, 1500, 2000, and 2500 μg/mL). The cells were incubated for 24 and 48 h, respectively, to determine apoptotic effects. Briefly, after treatment, cells were washed twice with ice-cold phosphate-buffered saline, lysed with 500 μL of ice-cold lysis buffer, scraped, and sonicated on ice. The cell lysate was then microcentrifuged for 10 min at 4°C, and the supernatant was stored at −80°C before analysis. One hundred microliters of the soluble fraction was used in each well of the ELISA plate. The ELISA was performed according to the manufacturer's instructions. Absorbance was measured at 450 nm with a microplate reader.
Reverse transcription–polymerase chain reaction to amplify VEGF mRNA
Total RNA was extracted from cultured LNCaP cells according to the manufacturer's instructions using the RNeasy Protect Mini Kit (Qiagen, Manheim, Germany). RNA concentration and purity was calculated after measuring absorbance at 260 nm on a UV spectrophotometer and then stored at −80°C. First-strand cDNA synthesis was performed with the QuantiTect Reverse Transcription cDNA Kit (Qiagen, Manheim, Germany). The synthesized cDNA was used as a template for polymerase chain reaction (PCR) amplification. Quantitative PCR was performed on a LightCycler (Roche Diagnostics GmbH, Mannheim, Germany) using TaqMan technology with amplification of human glyceraldehyde phosphate dehydrogenase (GAPDH) mRNA as a housekeeping standard. Oligonucleotide sequences of the cDNA primers were designed at Gene Research Laboratories, United Kingdom. The forward primer for VEGF was 5′-CCAGGAAAGACTGATACAGAACG-3′ and the reverse primer was 5′-GGTTTCTGGATTAAGGACTGTTC-3′. The following LightCycler conditions were used: initial denaturation at 95°C for 10 min, followed by 50 cycles with denaturation at 95°C for 20 s, annealing at 55°C for 30 s, and elongation at 72°C for 20 s. Cycle threshold values were determined by automated threshold analysis. The primer quality (lack of primer dimer amplification) was confirmed by melting curve analysis. Relative quantification of gene expression was performed using a standard curve constructed with serial dilutions of control mRNA or real-time PCR amplicons. All experiments were carried out in triplicate. VEGF levels were standardized to GAPDH (ratio VEGF:GAPDH) to account for loading differences. Gene expression levels (mRNA) were reported using the median as a point estimator and the range of values.
Statistical analysis
In the cell proliferation assay, experiments were repeated three times, measurements within an experiment were done in six duplicates, and in cell cycle analysis, experiments were repeated three times. Data from three independent experiments were expressed as mean ± standard deviation. Data were analyzed by a computerized probit analysis, which provided an estimate of the half maximal inhibitory concentration (IC50). The Shapiro–Wilk test was used as the normality test for the distribution of continuous variables. Tukey's test was used for normally distributed variables and Kruskal–Wallis test was used for non-normally distributed variables. A P-value below .05 was accepted to denote statistical significance.
Results
Effect of A. graveolens on cell viability
The effect of A. graveolens extract on the viability of LNCaP cells is shown in Figure 1. The XTT assay showed that A. graveolens extract decreased the viability of LNCaP cells. A. graveolens concentrations ranging from 1500 to 3000 μg/mL at 24 h and from 2000 to 3000 μg/mL at 48 h showed a dose–response relationship of cell survival (24 h: control vs. 1500 μg/mL P < .05, control vs. 2000 μg/mL P < .001, control vs. 2500 μg/mL P < .001, and control vs. 3000 μg/mL P < .001; 48 h: control vs. 2000 μg/mL P < .001, control vs. 2500 μg/mL P < .001, and control vs. 3000 μg/mL P < .001), whereas A. graveolens concentrations ranging from 2500 to 3000 μg/mL showed a time-dependent decrease (2500 μg/mL: 24 h vs. 48 h, P < .001; 3000 μg/mL: 24 h vs. 48 h, P < .001).

Effect of Apium graveolens treatment on the viability of prostate cancer cell lines LNCaP.
The IC50 value was 2840 μg/mL for the 48 h of treatment with extract. Based on this experiment, three concentrations (1500, 2000, and 2500 μg/mL) lower than the IC50 were chosen for later experiments examining the effects of A. graveolens on LNCaP cells.
Effect of A. graveolens on apoptosis of LNCaP cells
A. graveolens induces apoptotic cell death in LNCaP cells. The levels of cleaved PARP, one of the best biomarkers of apoptosis, were analyzed after 24 and 48 h of treatment with A. graveolens. Full-length active PARP is a 116 kDa molecule, which is cleaved to fragments of 86 and 30 kDa by the action of caspase-3 and related caspases. A marked increase in cleaved PARP was observed after 48 h of exposure to A. graveolens at concentrations of 2000 and 2500 μg/mL (P < .01, P < .01). No changes were observed after 24 h exposure to A. graveolens (Fig. 2).
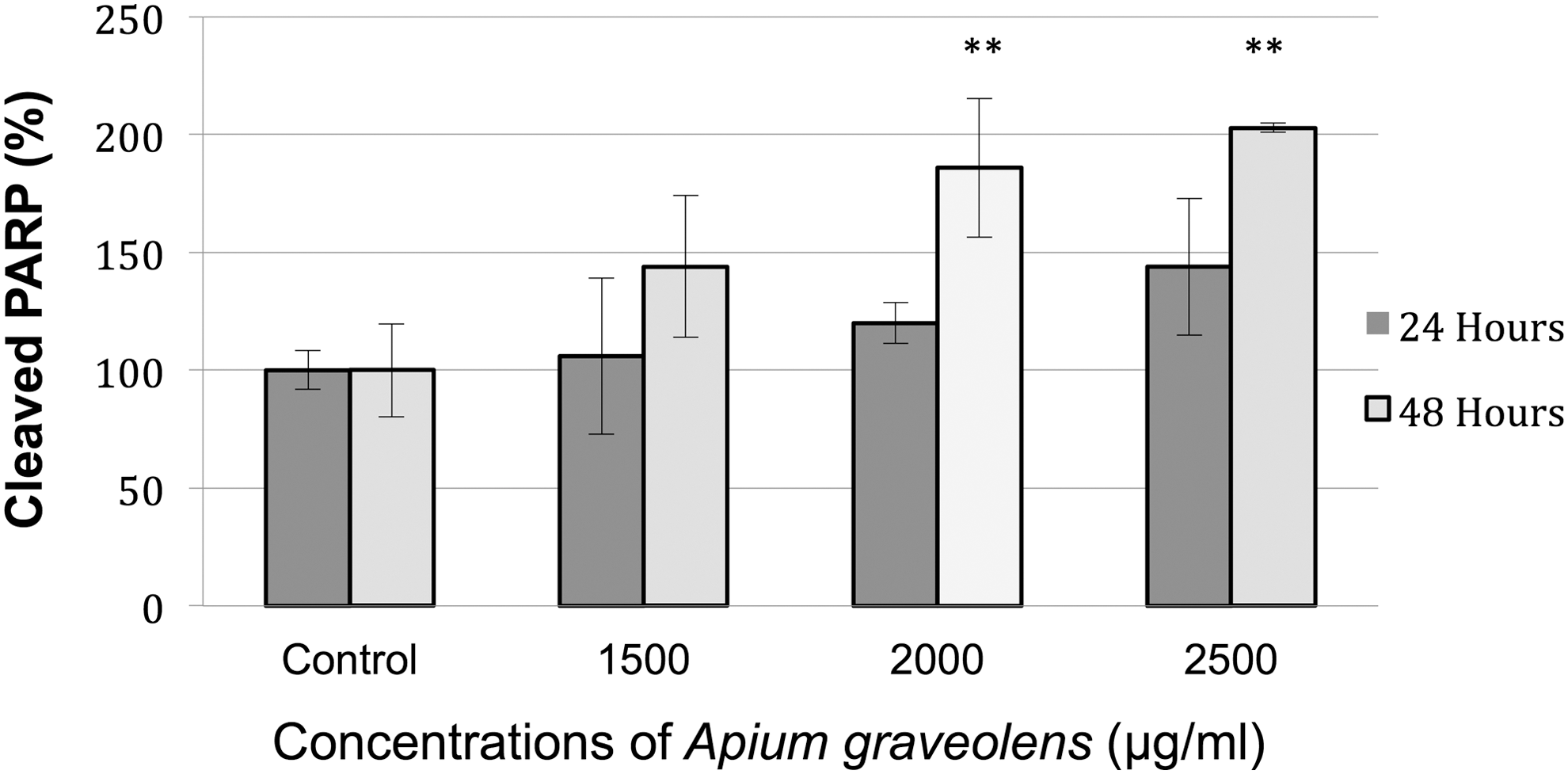
Effects of A. graveolens treatment on PARP, cleavage in prostate cancer cell lines LNCaP. The cells were treated with various concentrations (0, 1500, 2000, and 2500 μg/mL) of A. graveolens for 24 and 48 h. Values represent mean ± SD (**P < .01 compared with control group). PARP, poly (ADP-ribose) polymerase.
Effect of A. graveolens on VEGF mRNA expression
The dose-dependent effect of A. graveolens extract on LNCaP cells showed a significantly decreased VEGF mRNA expression at 1500, 2000, and 2500 μg/mL when assessed at 48 h posttreatment. No changes were observed after 24 h of exposure to A. graveolens (Fig. 3).
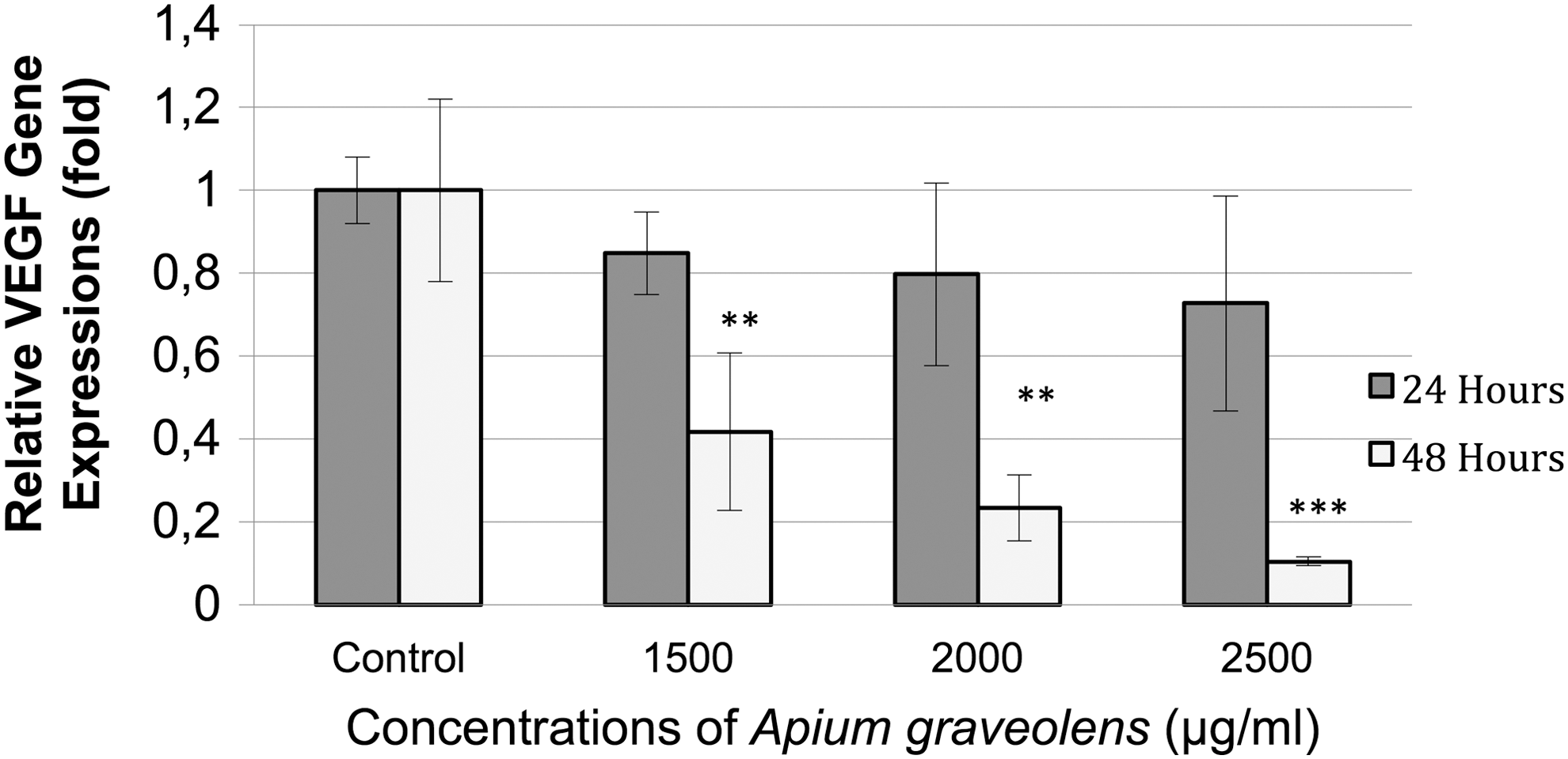
Effect of A. graveolens treatment on VEGF mRNA expression in prostate cancer cell lines LNCaP. The cells were treated with various concentrations (0, 1500, 2000, and 2500 μg/mL) of A. graveolens for 24 and 48 h. Values represent mean ± SD. (**P < .01; ***P < .001 compared with control group). VEGF, vascular endothelial growth factor.
Discussion
Several studies have demonstrated the potential effects of natural compounds as anticancer agents in vitro as well as in vivo. The aim of this study was to evaluate the anticancer effect of A. graveolens extract on human prostate carcinoma LNCaP cells. We found that A. graveolens extract inhibited the proliferation and viability of human prostate cancer LNCaP cells through induction of apoptosis. LNCaP cells showed clear apoptosis within 48 h after A. graveolens (2000 and 2500 μg/mL) treatment.
Apoptosis, a complex process that leads to cell death, is mediated by programmed signaling pathways that can be started by various extracellular or intracellular stimuli. 20 The ability to efficiently induce apoptosis is essential for the development of effective cancer therapies. 21 In the present study, we demonstrated that A. graveolens treatment of LNCaP cells resulted in both a time and dose-dependent increase of the cleavage of PARP, which is considered a biochemical hallmark of apoptosis. The enzyme PARP is localized within the cell nucleus and catalyzes DNA repair. During apoptosis, PARP is enzymatically cleaved. Detection of the cleavage products is characteristic for apoptosis.
Regarding the chemical composition of A. graveolens extract, we could hypothesize that its apoptotic effects might be dependent on the presence of phenolic acids and flavonoids. Chemical analysis has demonstrated that major phenolic acids identified in the extracts of celery are caffeic acid, p-coumaric acid, and ferulic acid, whereas the identified flavonoids are apigenin, luteolin, and kaempferol. 5
Studies have suggested several mechanisms for phenolic compounds mediating apoptosis of prostate cancer cells. For example, Mc Eleny et al. have shown that caffeic acid phenethyl ester (CAPE) causes apoptosis through reduction in the cellular levels of cellular inhibitor of apoptosis-1 (cIAP-1), cIAP-2, and X-linked inhibitor of apoptosis protein (XIAP) in human prostate cancer cells (PC-3). 22 Another study has shown that the main phenolic compounds in an ethanolic extract of propolis are caffeic acid, p-coumaric acid, and ferulic acid, which sensitize prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis through upregulation of TRAIL-R2 expression. 23
Apigenin is the dominant flavonoid in celery and possesses anti-inflammatory, 24 antiproliferative, 25 and anticarcinogenic properties. 26 Apigenin inhibits the growth of malign cells by inducing apoptosis through mechanisms that are not fully explored. Shukla et al. have suggested several mechanisms for these effects through blocking multiple signal transduction pathways, such as nuclear factor-kappa B (NF-κB), insulin-like growth factor 1 (IGF-1) axis, phosphatidylinositol 3-kinase (PI3K)-Akt, and b-catenin in various experimental models of prostate cancer. 27 –30 They have also shown that apigenin inhibits p53 activation and class I histone deacetylases (HDACs) in prostate cancer cells. 31,32 Luteolin is a common dietary flavonoid and has been found to cause apoptosis through inhibition of IGF-1R/AKT signaling, 33 downregulation of androgen receptor expression, 34 upregulation of death receptor 5 (DR5), 35 and inhibition of fatty acid synthase 36 in human prostate cancer cells. Kaempferol, a flavonol, is a polyphenol compound present in fruits and vegetables. Several reports have described that kaempferol has antiproliferative activities and/or induces apoptosis in different human cancer cell lines, including prostate cancer cells. 37
Angiogenesis plays a pivotal role in tumor growth, invasion and metastasis, and is tightly regulated by a large number of proangiogenic and antiangiogenic factors. Among these factors, VEGF is one of the most critical and specific angiogenesis factors. VEGF, through binding VEGF receptor 2 (VEGFR-2), induces a cascade of signaling pathways that result in proliferation, migration, survival, and vascular permeability. Hypoxic conditions activate the uptake of VEGF and other growth factors and induce the growth of neovasculature, allowing tumor cells to gain access to oxygen and nutrients. 38,39 Increased VEGF expression is closely associated with increased intratumoral microvessel density, invasive depth, lymphatic and venous invasion, lymph node metastasis, and a poor prognosis in cancer patients. 40,41 As a result, VEGF and its receptors have been the most commonly targeted molecules found in antiangiogenic drugs for different types of tumors. 42 –44 Therefore, we evaluated VEGF gene expression levels. Although we did not investigate the mechanism, we demonstrated that VEGF expression was downregulated by treatment with A. graveolens. Similar results indicated that apigenin inhibits the expression of VEGF in human ovarian cancer cells, 45 human prostate cancer PC3-M cells, 46 A549 lung cancer cells, 47 and in MDA-MB-231 human breast cancer cells. 48 Like apigenin, other dietary flavonoids studied for VEGF expression in cancer cells include kaempferol, which inhibits VEGF expression in ovarian cancer cells, 49 and luteolin, which inhibits VEGF expression in pancreatic carcinoma cells by inhibiting the transcription activity of NF-κB. 50
Conclusions
Our present study demonstrated: (1) A. graveolens extracts decreased the viability of human prostate carcinoma LNCaP cells in an XTT assay in a time- and dose-dependent manner; (2) Apoptosis in A. graveolens-treated LNCaP cells was shown by an increase of the cleavage of PARP; (3) VEGF expression was downregulated by treatment with A. graveolens. In conclusion, although further studies are required, the results of this study indicate that A. graveolens extract may be useful for the management of prostate cancer.
Footnotes
Acknowledgment
This work received financial support from the Afyon Kocatepe University Scientific Research Project Commission (Research Grant: 11-TIP-10), Afyonkarahisar, Turkey.
Author Disclosure Statement
No competing financial interests exist.