
Abstract
Apigetrin is a flavonoid isolated from various herbal medicines such as Scutellaria baicalensis Georgi, Matricaria chamomilla, Stachys tibetica Vatke, and Teucrium gnaphalodes. In the present study, we investigated the inhibitory effects of apigetrin on neuroinflammation using the BV-2 microglia cell line. Our data revealed that apigetrin significantly reduced secretion and mRNA expression of inflammatory cytokines, tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), in lipopolysaccharide (LPS)-stimulated BV-2 mouse microglia. Apigetrin also significantly decreased LPS-mediated production of prostaglandin E2 (PGE2) level and nitric oxide (NO) production as well as expression of cyclooxygenase 2 (COX-2) and inducible nitric oxide synthase (iNOS) in BV-2 cells. In addition, apigetrin suppressed nuclear expression of nuclear factor kappa B (NF-κB) in LPS-stimulated BV-2 cells. Furthermore, apigetrin significantly impaired reactive oxygen species (ROS) generation and enhanced expression of antioxidant enzymes, hempxygenase 1 (HO-1) and nuclear factor-like 2 (Nrf2), in BV-2 cells. Apigetrin also increased 2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical scavenging activity, indicating antioxidative activity of apigetrin. Moreover, we found that apigetrin inhibited hydrogen peroxide (H2O2)-induced cell death in HT22 hippocampal cells. Overall, our findings indicate that apigetrin has inhibitory effects on neuroinflammation as well as antioxidation and neuroprotection, suggesting the potential prophylactic activity for neurodegenerative diseases through the inter-regulation of neuroinflammation, oxidative stress, and neuronal injury.
Introduction
N
Neuroinflammation is the inflammatory response in the central nervous system by various neuronal injuries. 5 Microglia are the major resident immune cells in the neuronal system unit and can be activated by various inflammatory stimulators. 6,7 Activated microglia release the inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6) and mediate neuronal damage. 8,9 Notably, Morales et al. reported that nonsteroidal anti-inflammatories have the ability to reduce the occurrence of Alzheimer's disease, suggesting that anti-inflammatory regulation is an important point for treating Alzheimer's disease. 10 Additionally, neuroinflammation can trigger oxidative stress by generating excessive reactive oxygen species (ROS) in microglia. 11 That is, inter-relation between neuroinflammation and sustained oxidative stress is a pivotal feature in neurodegenerative diseases such as Alzheimer's disease. 12
Scutellaria baicalensis Georgi is a traditional medicinal herb used for several infectious diseases, such as dysentery and carbuncles, and reducing fever. 13 Many recent articles reported its biological activities such as anti-cancer, 14 anti-inflammation, 15 and anti-diabetes. 16 In addition, S. baicalensis Georgi inhibited the lipopolysaccharide (LPS)-induced expression of cyclooxygenase 2 (COX-2) and inducible nitric oxide synthase (iNOS), inflammatory mediators, in BV-2 microglia, suggesting anti-neuroinflammatory activity. 17 Several major marker compounds of S. baicalensis Georgi, such as baicalein, wogonin, apigenin, and chrysin, also showed the inhibitory effects on neuroinflammation. 18 –21
Apigetrin, also called apigenin 7-glucoside or cosmosin, is a polyphenol and one of the marker compounds of S. baicalensis Georgi. 22 In addition to S. baicalensis Georgi, apigetrin is isolated from various herbal plants such as Matricaria chamomilla, 23 Stachys tibetica Vatke, 24 and Teucrium gnaphalodes. 25 Apigetrin has been reported to have several pharmacological properties such as anti-melanogenesis, 24 anti-cancer, 26,27 hepatoprotective effects, 28 and skin anti-inflammatory effects. 29 Interestingly, Patil et al. reported that apigetrin improved the cognitive performance and inhibited COX-2 and iNOS in an LPS-intoxicated mouse model. 30 Although Patil et al.'s article presented the anti-neuroinflammatory potential of apigetrin, there is no critical evidence of the inhibitory action on inflammation in microglia, a pivotal mediator of neuroinflammation. 31
Therefore, our present study was aimed to investigate the inhibitory effects of apigetrin against neuroinflammation using LPS-stimulated BV-2 microglia. Our data demonstrated that apigetrin exerts anti-neuroinflammatory effects in microglia by suppressing production of inflammatory molecules. Furthermore, apigetrin had antioxidant activity in microglia as well as neuroprotective effect in damage-induced hippocampal cells.
Materials and Methods
Cell lines and culture
BV-2 and HT22 cells were maintained in Dulbecco's modified Eagle's medium (Hyclone/Thermo, Rockford, IL, USA) supplemented with 10% fetal bovine serum (Hyclone/Thermo) and penicillin/streptomycin in 5% CO2 at 37°C. BV-2 cells were pretreated with apigetrin (KOC Biotech Co., Ltd., Daejeon, Korea) for 2 h and treated with LPS (1 μg/mL; Sigma-Aldrich, St. Louis, MO, USA) for an additional 22 h. HT22 cells were cotreated with apigetrin and hydrogen peroxide (H2O2, 500 μM; Sigma-Aldrich) for 6 h.
Cytotoxicity assay
Cytotoxicity test was performed as previously described.
32
In brief, BV-2 and HT22 cells were plated on 96-well microplates at a density of 3 × 104/well and 5 × 103/well, respectively. Cells were treated with various concentrations of apigetrin for 24 h. CCK-8 solution (Dojindo, Kumamoto, Japan) was added, and the cells were incubated for 4 h. The absorbance was read at 450 nm on an Epoch microplate spectrophotometer (BioTek Instruments, Inc., Winooski, VT, USA). The cell viability was calculated using the following equation:
Enzyme-linked immunosorbent assays for cytokine production
BV-2 cells were pretreated with various concentrations of apigetrin for 2 h and stimulated with LPS (1 μg/mL) for an additional 22 h. Production of TNF-α, IL-6, and prostaglandin E2 (PGE2) in the culture supernatants was measured using commercial enzyme-linked immunosorbent assay (ELISA) kits from R&D Systems (Minneapolis, MN, USA) or Cayman Chemical Co. (Ann Arbor, MI, USA).
Nitric oxide (NO) assay
NO synthesis was analyzed by determining the accumulation of nitrite (NO2 −) in culture supernatant using the Griess Reagent System (Promega, Madison, WI, USA). Equal volume of supernatant and sulfanilamide solution was mixed and incubated for 10 min at room temperature, and then naphthylethylenediamine dihydrochloride solution was added for additional 5 min. The absorbance was measured at 540 nm by using an Epoch microplate spectrophotometer. The nitrite concentration was determined from a standard curve generated with sodium nitrite (NaNO2).
Reverse transcription–polymerase chain reaction
Total RNA was prepared using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and reverse transcribed to cDNA using an iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA). The expression of inflammation-related genes was analyzed by reverse transcription–polymerase chain reaction (RT-PCR) using the primers listed in Table 1. RT-PCR products were separated on a 1.5% agarose gel and visualized by Azure C150 Gel Imaging Workstation (Azure Biosystems, Dublin, CA, USA).
Western blotting
Western blot analyses were carried out as previously described. 32 Whole-cell extracts were prepared by suspending cells in an extraction lysis buffer (Sigma-Aldrich) containing protease inhibitor cocktail (GenDEPOT, Barker, TX, USA). Nuclear extracts were isolated using the NE-PER nuclear and cytoplasmic extraction reagents (Thermo Scientific, Rockford, IL, USA) according to the manufacturer's protocol. The protein concentration was determined using a Bradford reagent (Sigma-Aldrich). Equal amounts of cell extract (20–30 μg) were resolved by 4–20% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene fluoride (PVDF) membrane. The membrane was incubated with blocking solution (5% skim milk in Tris-buffered saline containing Tween 20 [TBST]), followed by an overnight incubation at 4°C with the appropriate primary antibodies; heme oxygenase 1 (HO-1) (Abcam, Boston, MA, USA), nuclear factor-like 2 (Nrf2), nuclear factor kappa B (NF-κB) p65, nucleolin, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Santa Cruz Biotechnology, Dallas, TX, USA). The membranes were washed three times with TBST and then incubated with a horseradish peroxidase-conjugated secondary antibody (Jackson ImmunoResearch, West Grove, PA, USA) for 1 h at room temperature. The membranes were again washed three times with TBST and then developed using an enhanced chemiluminescence (ECL) kit (Thermo Scientific). Image capture was performed using ImageQuant LAS 4000 mini luminescent image analyzer (GE Healthcare Bio-Sciences, Piscataway, NJ, USA).
ROS detection
ROS generation was measured using the 2′,7′-dichlorofluorescin diacetate (DCFDA) Cellular ROS Detection Assay Kit (Roche Applied Science, Indianapolis, IN, USA). BV-2 cells were grown on 96-well microplates at a density of 5 × 103/well. Cells were pretreated with various concentrations of apigetrin for 1 h and treated with LPS for an additional 5 h and then stained with DCFDA for 45 min at 37°C. After removing DCFDA solution, phosphate-buffered saline (100 μL/well) was added into the cells and plates were read at the end point in the presence of apigetrin on a fluorescent plate reader (SpectraMax i3 Multi-Mode Platform; Molecular Devices Co., Sunnyvale, CA, USA) with excitation wavelength at 485 nm and emission wavelength at 535 nm.
2,2′-Azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) radical scavenging activity
The free radical scavenging activity of apigetrin on the 2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) was assessed using the method described by Re et al.
33
ABTS radical cations were produced by reacting 7 mM ABTS solution with 2.45 mM potassium persulfate in the dark at room temperature for 16 h. Absorbance of the reactant was later adjusted to 0.7 at a wavelength of 734 nm; 100 μL aliquots of apigetrin solution at various concentrations were mixed with 100 μL ABTS•+ solution. The reaction mixture was incubated for 5 min in the dark at room temperature. The absorbance of the resulting solution was measured at 734 nm with an Epoch microplate spectrophotometer. The radical scavenging capacity of the apigetrin-treated samples was calculated using the following equation:
Statistical analysis
Data are expressed as the mean ± SEM. Data were analyzed using one-way analysis of variance and Dunnett's multiple comparisons test. P < .05 was considered significant.
Results
Cytotoxic effects of apigetrin in BV-2 microglia
To assess cytotoxicity of apigetrin against BV-2, CCK assay was performed. Cells were treated with various concentrations (0, 12.5, 25, 50, or 100 μg/mL) of apigetrin for 24 h. As shown in Figure 1, apigetrin had no cytotoxicity up to 100 μg/mL in BV-2 cells. Subsequent experiments were performed under nontoxic concentration.

Cytotoxicity of apigetrin in BV-2 microglia. Cells were seeded on 96-well plates and treated with various concentrations (0, 12.5, 25, 50, or 100 μM) of apigetrin for 24 h. Cell viability was assessed using CCK-8 assay. The results are expressed as mean ± SEM of three independent experiments.
Effects of apigetrin on the production of inflammatory factors in LPS-stimulated BV-2 cells
To investigate the effects of apigetrin on the production of inflammatory cytokines, the amount of TNF-α and IL-6 was measured using culture supernatant from the LPS-stimulated BV-2 cells. LPS stimulation significantly increased levels of TNF-α and IL-6 in BV-2 microglia. In contrast, apigetrin treatment significantly reduced LPS-mediated release of TNF-α and IL-6 in BV-2 cells (Fig. 2A, B). The apigetrin effects on inflammatory mediators, PGE2 and NO, were also evaluated in LPS-stimulated BV-2 cells. Treatment with LPS mediated a significant increase in PGE2 and NO levels. In contrast, apigetrin significantly decreased LPS-mediated PGE2 and NO production (Fig. 3A, B).
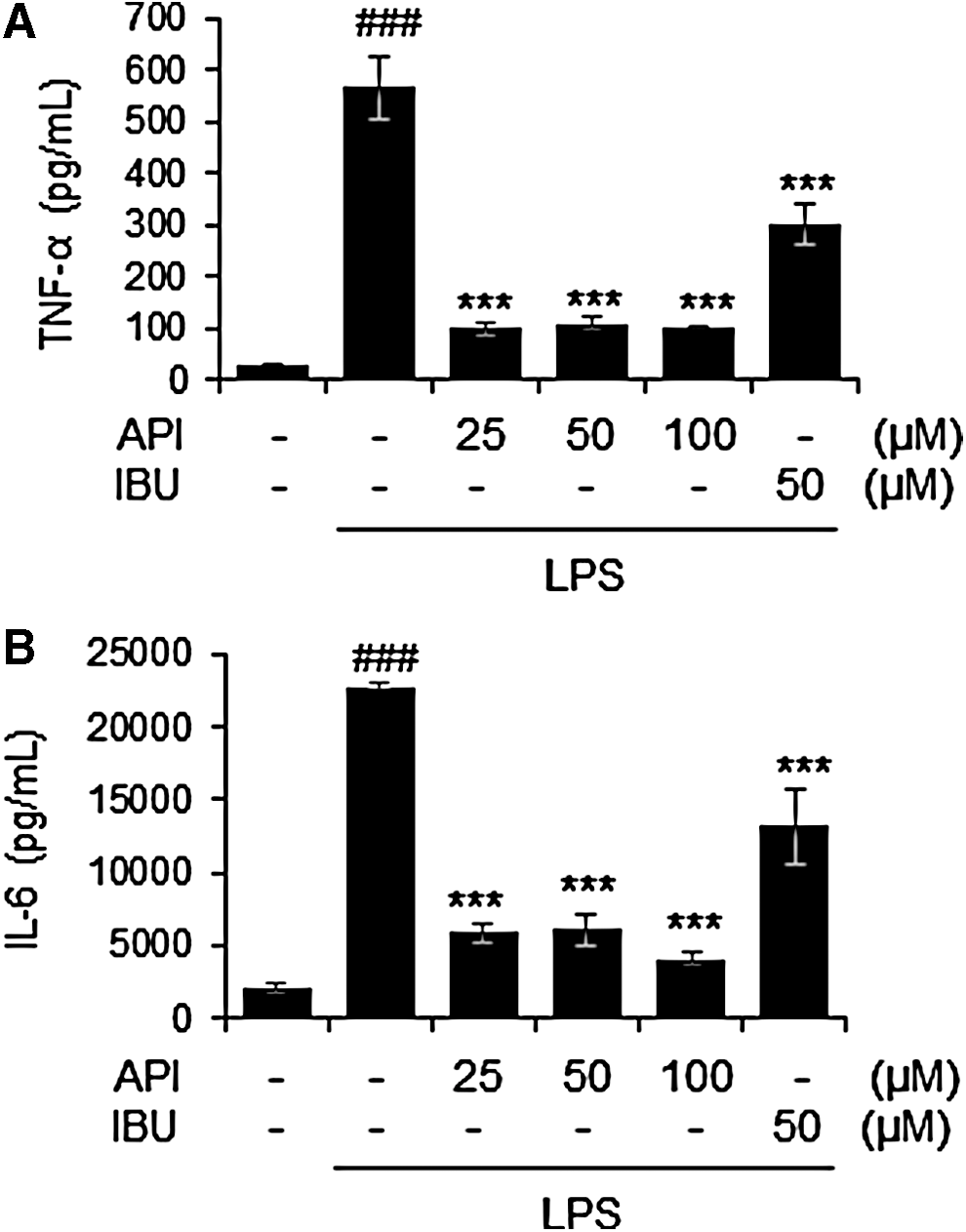
Effects of apigetrin on LPS-induced production of proinflammatory cytokines in BV-2 cells. Cells were pretreated with apigetrin (0, 25, 50, or 100 μM) for 2 h and then stimulated with LPS (1 μ/mL) for an additional 22 h. The production of TNF-α
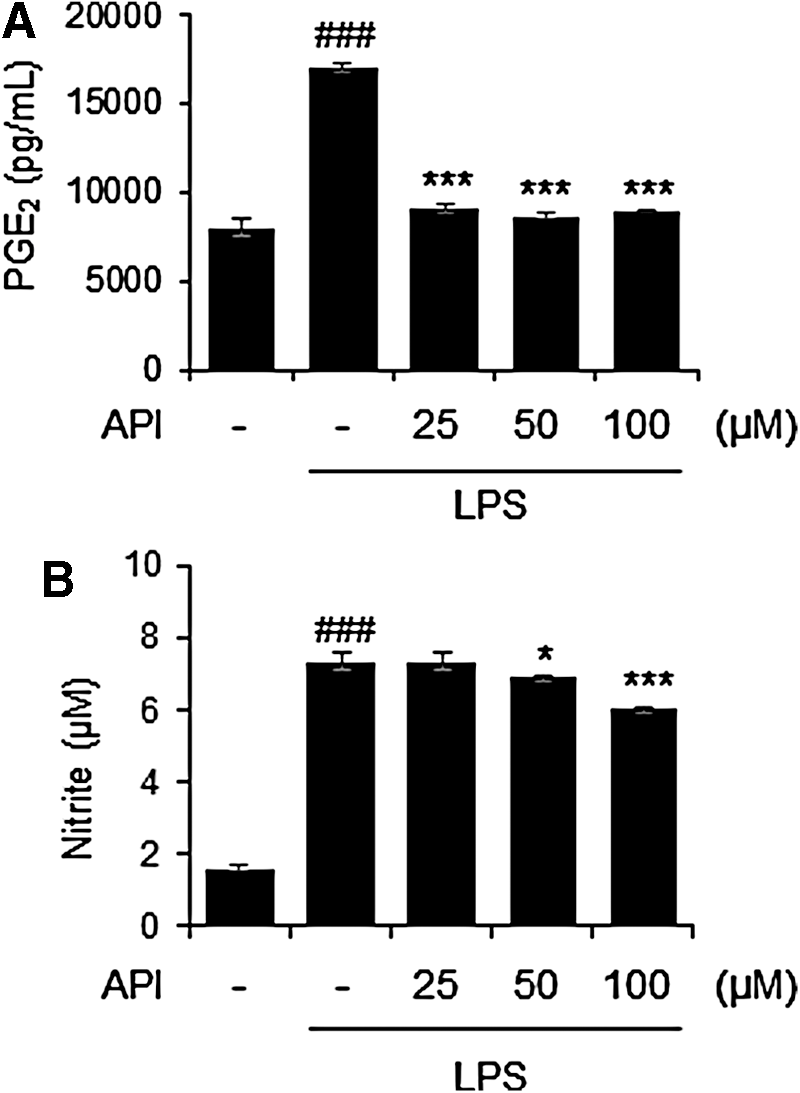
Effects of apigetrin on LPS-induced production of PGE2 and nitrite in BV-2 cells. Cells were pretreated with apigetrin (0, 25, 50, or 100 μM) for 2 h and then stimulated with LPS (1 μ/mL) for an additional 22 h. The production of PGE2
Effects of apigetrin on the expression of inflammatory molecules in LPS-stimulated BV-2 cells
To confirm the importance of inflammation-related factors in the inhibition of apigetrin on neuroinflammation, the mRNA expression of inflammatory molecules was analyzed by RT-PCR. In parallel with the results of Figures 2 and 3, apigetrin treatment suppressed the mRNA levels of TNF-α, IL-6, COX-2, and iNOS in LPS-stimulated BV-2 cells (Fig. 4A, B, respectively).
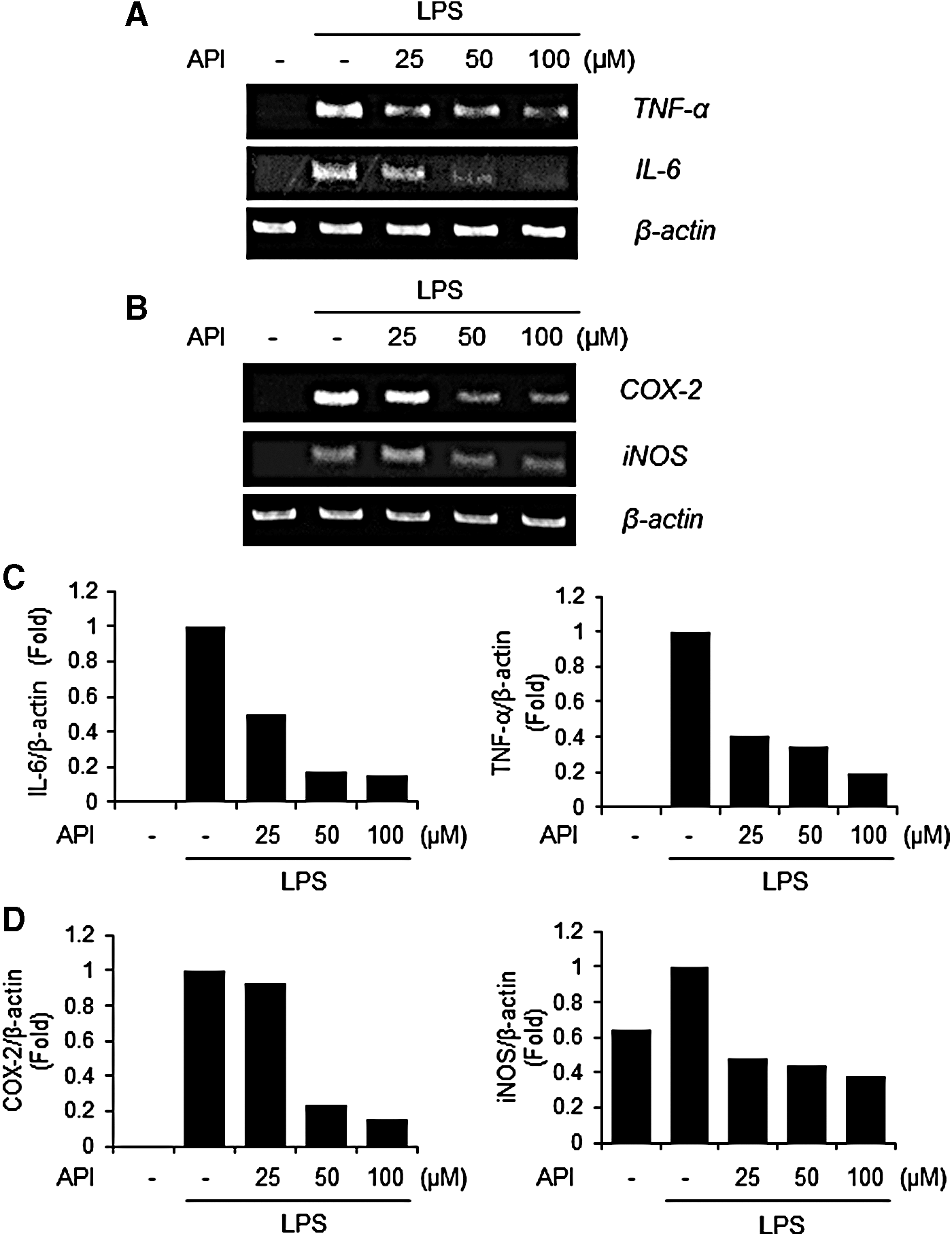
Effects of apigetrin on LPS-induced mRNA expression of inflammatory molecules in BV-2 cells. Cells were pretreated with apigetrin (0, 25, 50, or 100 μM) for 1 h and then stimulated with LPS (1 μ/mL) for an additional 5 h. Total RNA was isolated and subjected to RT-PCR to analyze TNF-α or IL-6 expression
Effects of apigetrin on NF-κB activation in LPS-stimulated BV-2 cells
Next, we investigated whether apigetrin affects activation of NF-κB, a key factor in the molecular regulation of neuroinflammation. 34 Nuclear extract was prepared from the BV-2 cells stimulated with LPS in the absence or presence of apigetrin. As shown in Figure 5, LPS stimulated dramatically increased nuclear expression of NF-κB p65. In contrast, apigetrin treatment markedly reduced the LPS-mediated increase of NF-κB p65 in the nucleus in a dose-dependent manner. Nucleolin was used as an internal control.
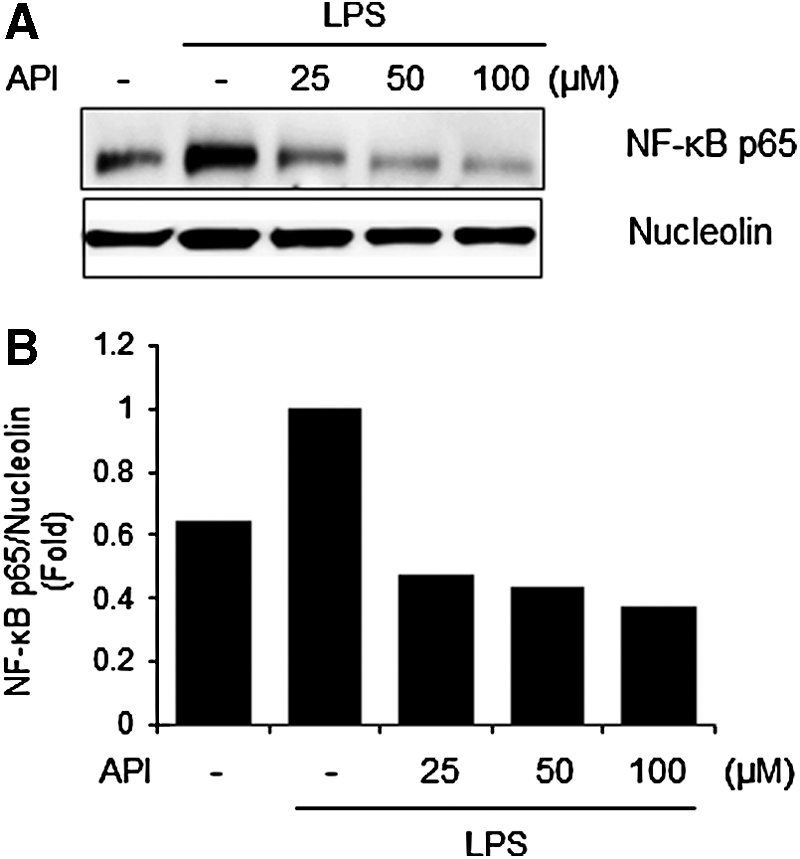
Effects of apigetrin on LPS-induced NF-κB nuclear translocation in BV-2 cells.
Inhibitory effects on ROS generation and antioxidant effects of apigetrin
Neuroinflammation is associated with oxidative stress-mediated excessive ROS production. 35 Thus, we measured the effects of apigetrin on ROS generation in LPS-stimulated BV-2 using DCFDA. ROS levels were significantly increased by LPS compared with untreated control. In contrast, apigetrin significantly reduced the LPS-stimulated ROS production in a dose-dependent manner (Fig. 6A). In addition, protein expression of antioxidant enzymes, HO-1 and Nrf2, was enhanced by apigetrin in BV-2 cells (Fig. 6B). Furthermore, apigetrin dramatically increased the ABTS radical scavenging activity in a dose-dependent manner (Fig. 6C).

Antioxidant activities of apigetrin.
Neuroprotective effects of apigetrin in H2O2-treated HT22 hippocampal cells
To assess whether apigetrin has neuroprotective activity, we utilized HT22 hippocampal cells. Cytotoxicity of apigetrin was measured against HT22 cells and it revealed a dose-dependent decrease of cell viability (Fig. 7A). Cell viability was maintained about 100% at lower levels, 25 μg/mL, and reduced to 88.65% and 66.33% at 50 and 100 μg/mL, respectively. Then, neuronal cell damage of HT22 was induced by stimulating with H2O2 and concurrently treated with various concentrations of apigetrin. Stimulation with H2O2 alone significantly decreased the cell viability compared with untreated control. In contrast, apigetrin reversed H2O2-mediated cell death at 5 μM of treatment. However, higher concentration (10 μM) of apigetrin further increased the cytotoxicity of H2O2 treatment in HT22 cells (Fig. 7B).

Neuroprotective effect of apigetrin in H2O2-damaged HT22 cells.
Discussion
Neuroinflammation and oxidative stress are the major causes of neurodegenerative diseases such as Alzheimer's disease and Parkinson's disease. 36 Various inflammatory factors, such as β-amyloid, LPS, or interferon-gamma, stimulate activation of microglia, key resident cells in the neuronal system. Activated microglia lead to generation of excessive amounts of ROS and release the inflammatory cytokines or mediators and mediate neuronal cell damage. 37 Thus, it is important to control the balance between inflammation and redox signaling for progression of neurological diseases.
In the present study, we investigated the effects of apigetrin from S. baicalensis Georgi, a traditional Korean herbal medicine, on neuroinflammation and oxidative stress in microglia. Inflammatory reaction was induced by stimulating with LPS in the BV-2 microglia cell line. As expected, LPS significantly enhanced the production of inflammatory cytokines, TNF-α and IL-6, consistent with previous reports. 38 Apigetrin administration inhibited LPS-induced production and expression of these cytokines in BV-2 cells, indicating anti-neuroinflammatory effects of apigetrin. Especially, TNF is a master factor and plays an essential role in the inflammatory regulation and also has the ability to mediate oxidative stress by generating ROS. 12 Hence, anti-TNF therapy is thought to be a valuable approach for several inflammatory diseases such as rheumatoid arthritis and bowel disease. 39 –41 Interestingly, since TNF released from activated microglia can accelerate inflammatory response as well as oxidative stress, thereby boosting neurodegeneration, 12 anti-TNF therapy may be considered as an attractive trial for neurodegenerative diseases.
In addition, we examined whether apigetrin can influence the generation of inflammatory mediators, PGE2 and NO. During chronic inflammation in the brain, inflammation stimuli increase the expression of COX-2 and iNOS for production of PGE2 and NO, respectively, in microglia. 42 In this study, apigetrin significantly inhibited LPS-stimulated production of PGE2 and NO by suppressing the mRNA expression of COX-2 and iNOS in BV-2 cells. There are accumulating evidences that COX-2 and iNOS expression is associated with ROS production mediated by inflammatory factors such as cytokines. 9,37 Apigetrin treatment significantly inhibited LPS-stimulated ROS generation in BV-2 cells. Apigetrin also showed a significant scavenging activity against ABTS radical. A number of anti-neuroinflammatory agents exert their activity through the upregulation of antioxidant enzyme(s). 43 –45 Our results present that apigetrin markedly enhanced expression of antioxidant enzymes, HO-1 and Nrf2, in BV-2 cells. Taken together, these data indicate that apigetrin could act as a suppressor of microglial activation.
It is well known that NF-κB is a key regulator in the inflammatory reaction. Targeting NF-κB transactivation can induce the suppression of inflammation-related factors, COX-2, iNOS, and cytokines, in microglia. 46 Recently, Kaur et al. reported an interesting review article that NF-κB is a critical molecule to connect between oxidative stress and neuroinflammation in the pathogenesis of Alzheimer's disease. 35 In our study, LPS stimulation dramatically augmented the nuclear expression of NF-κB p65 in BV-2 cells, indicating the nuclear translocation of activated NF-κB. In contrast, apigetrin reduced LPS-activated p65 nuclear translocation in a dose-dependent manner. These results suggest that anti-neuroinflammatory and antioxidative actions of apigetrin in microglia are associated with, at least in part, NF-κB activation.
We finally tested whether apigetrin has neuroprotective effect using HT22 hippocampal cells. HT22 cells were treated with H2O2 to induce oxidative stress and revealed a significant reduction in cell viability by H2O2. In contrast, apigetrin treatment reversed H2O2-induced cell death at 5 μM. Higher concentration of apigetrin with H2O2 treatment further increased the cytotoxicity compared with H2O2 alone, implying the protective effect of apigetrin on neuronal cell death.
In conclusion, our present data address the potent prophylactic activity of apigetrin on anti-inflammation and antioxidation in neurodegenerative diseases. This antineuroinflammatory effect is regulated by suppressing the production and expression of inflammatory cytokines and mediators through the blockade of NF-κB activation. Further studies will be necessary to confirm the neuroprotection by apigetrin and its connection with anti-inflammation and ROS generation using in vitro or in vivo models.
Footnotes
Acknowledgment
This study was supported by a research grant (K16293) from the Korea Institute of Oriental Medicine (KIOM).
Author Disclosure Statement
No competing financial interests exist.