
Abstract
The deposition of amyloid protein as senile plaques is the major signature of Alzheimer's disease (AD). It is produced by the sequential cleavage of the amyloid precursor protein by secretases. Moreover, peppers are noted for their antiaging and cognitive enhancing properties. Thus, in this study, the effects of polyphenol-rich extracts from bell pepper on amyloid production and aggregation in vitro were investigated. Bell pepper (ripe and unripe) was extracted with methanol-1 N HCl (1:1 v/v). Thereafter, the inhibitory potentials of the extracts on β-secretase and β-amyloid1–40 aggregation were determined. Phenolic composition of the pepper fruits was further determined by HPLC-DAD (high performance liquid chromatography–diode array detector). There was a dose-dependent inhibition of β-secretase by the pepper fruits with the ripe fruits (2.17 ± 0.17 μg/L) showing a significantly (P < .05) higher inhibitory effect than the unripe (3.44 ± 0.11 μg/L). Furthermore, Thioflavin-T and transmission electron microscopy analyses revealed that phenolic extracts from pepper fruits (1 and 10 μg/L) could counteract the initial aggregation of Aβ 1–40, as well as prevent further aggregation preformed fibrils. These inhibitory activities could be attributed to the predominant presence of phenolic constituents in the pepper fruits. It is possible to conclude that bell pepper could be a possible dietary intervention into the management of AD.
Introduction
A
Amyloid toxicity is considered critical to AD pathogenesis. 4 Changes in its generation, clearance, and degradation are believed to be a part of the triggering molecular events in the pathogenic cascade of the disease. 3 Aβ peptides (mostly 40 or 42 amino acid residues) are generated from the sequential cleavage of amyloid precursor protein (APP) by beta and gamma secretases, respectively. 5 β-secretase is at present the most attractive therapeutic target in limiting Aβ peptide production since it is considered the rate-limiting step in the proteolytic processing of APP to generate Aβ. Moreover, its activity is elevated approximately twofold in the AD brain thereby initiating/accelerating AD pathogenesis. 5 –7
Aβ peptides are able to self-associate to varying assembly states ranging from dimers to soluble oligomers to insoluble aggregates of the fibrils. 8,9 The neurotoxicity of Aβ is highly dependent on the conformation, quaternary structure, and morphology of the aggregates. 10 Eventual neuronal degeneration involving disruption of cholinergic synaptic function is attributed to the oligomeric and fibrillary Aβ deposits. 8
At present, there is no cure for AD, and so there is a continuous search for novel compounds with the ability to modulate AD pathology. This search has extended into dietary compounds among which natural polyphenols have received a lot of attention. Natural polyphenols are among the most commonly consumed phytochemicals in herbal beverages and food worldwide and are able to offer some protection against many diseases. The primary function of phenolics is in the protection of plants against reactive oxygen species produced during photosynthesis and consumption by herbivores. 11 Generally, polyphenols have received most of its attention because of their antioxidant effect with comparable minimal focus on their anti-aging and protective effect against neurodegeneration.
In this study, we have investigated the anti-amyloidogenic potentials of phenolic-rich extracts from Capsicum annuum var. grossum (bell pepper) fruits at two stages of maturity (unripe and ripe). Capsicum spp. generally referred to as pepper have been a part of human diet since 7500BC 12 and are a popular culinary spice valued for their sensory attributes of color, pungency and flavor. 13 Interestingly, Capsicum spp. are constituents of decoctions/pot soups prescribed traditionally as anti-aging and cognitive enhancing agents in southwestern Nigeria. 14 Varying sensory qualities of peppers arise from changes in phytochemical composition during growth and maturation. 15,16 Bell pepper is mild in taste and pungency making it one of the most widely distributed and consumed of the Capsicum spp. It is a rich source of polyphenols/flavonoids for which numerous medicinal properties have been ascribed. 17,18 Furthermore, the flavonoids resveratrol, curcumin, and epigallocatechin gallate have been reported to alter APP and Aβ metabolism in models of AD. 11 Therefore, this study for the first time investigates the ability of phenolic-rich extracts from unripe and ripe C. annuum var. grossum to inhibit β-secretase and Aβ aggregation.
Materials and Methods
Materials
Ripe and unripe pepper fruits of C. annuum var. grossum were collected from farm settlements around Ile Ife township and authenticated at the herbarium of the Department of Botany, Obafemi Awolowo University, Ile Ife, Nigeria. Inedible portions were removed while edible portions were washed and lyophilized after which they were pulverized into fine powder.
Amyloid β protein fragment 1–40, Thioflavin-T, β-secretase (BACE 1) Activity Detection Kit, quercetin, isoquercitrin, quercitrin, rutin, kaempferol, capsaicin, and dihydrocapsaicin were procured from Sigma-Aldrich, Inc. (St Louis, MO, USA). Methanol, acetic acid, gallic acid and chlorogenic acid were purchased from Merck (Darmstadt, Germany).
Extraction of phenolics
Powdered pepper fruits were extracted with 20 volumes of the solvent, that is, methanol:water (1:1, v/v) 1M HCl for 48 h at room temperature and, thereafter, filtered through cheesecloth. The filtrate was concentrated at 40°C using a rotary evaporator until about 90% of the solvent was evaporated. Extracted polyphenols was thereafter obtained after lyophilization of the concentrate and stored at −20°C until further analysis. 19
Quantification of compounds by HPLC-DAD
Analytical HPLC-DAD (high performance liquid chromatography–diode array detector) was performed with a Shimadzu Prominence Auto Sampler (SIL-20A) HPLC system coupled with a DAD (SPD-M20A), while data were evaluated with LC solution 1.22 SP1 software. Separation was carried out under gradient conditions on a C18 column (4.6 × 150 mm) packed with 5 μm diameter particles; the mobile phase was water containing 2% acetic acid (A) and methanol (B), and the composition gradient was 5% of B until 2 min and changed to obtain 25%, 40%, 50%, 60%, 70%, and 100% B at 10, 20, 30, 40, 50, and 60 min, respectively, as described by Amaral et al. 20 with slight modifications. The flow rate was 0.7 mL/min, and the injection volume was 40 μL. The chromatography peaks were confirmed by comparing retention time with those of reference standards and by DAD spectra (200–600 nm). All chromatography operations were carried out at ambient temperature and in triplicate.
Inhibition of β-secretase
The β-secretase Activity (BACE 1) Assay Kit (Sigma) was used. The assay was carried out according to the manufacturer's instructions. Briefly, 2 μL of enzyme solution (0.3 units/μL) was reacted with 50 μM 7-methoxycumarin-4-acetyl-(Asn670, Lue671)-amyloid β/A4 precursor protein 770 fragment 667–676-(2,4 dinitrophenyl) lys-arg-arg amide trifluoroacetate salt and pepper extracts (0.01–100 μg/L) in a fluorescence assay buffer. Baseline readings were taken immediately on a fluorometer (excitation: 320 nm; emission: 405 nm) and repeated after 2 h incubation at 37°C. Results were reported as % inhibition of control.
Inhibition of β-amyloid (Aβ 1–40) peptide aggregation
Preparation of peptides
The inhibition of Aβ 1–40 aggregation was carried as described by Gupta and Rao 4 with slight modifications. For inhibition of aggregation experiments, Aβ 1–40 was initially dissolved in Milli-Q water and then made up with phosphate-buffered saline (PBS) to a final concentration of 200 μM. Pepper extracts previously dissolved in PBS (final concentration 1–10 μg/L) was mixed with Aβ 1–40 (final concentration 10 μM) and incubated at 37°C for 4 days. For the disaggregation experiments, 10 μM Aβ 1–40 was initially incubated at 37°C to generate amyloid aggregates. After 48 h pepper extracts dissolved in PBS (final concentrations 1–10 μg/L) were introduced in the Aβ 1–40 solution and further incubated for another 48 h at 37°C.
Thioflavin-T spectroscopy
Aliquots from each incubated sample were removed at t = 0, 24, 48, 72, and 96 h and treated with 25 μM Thioflavin-T (ThT) after which it was vortexed for about 10 sec before measurements. Fluorescence signals were determined on a spectrofluorometer at 450 nm (excitation)/480 nm (emission). The assay blank consisted of ThT solution mixed with PBS (pH 6.6). Fluorescence versus time curve was integrated to obtain the area under curve values, which were further used to calculate the % inhibition of aggregation/disaggregation as follows:
Transmission electron microscopy analysis
An aliquot of the Aβ 1–40 peptide incubated with pepper extracts was removed after 96 h and spotted on a copper grid (mesh size 200) coated with carbon film. Peptides were stained with 2% uranyl acetate solution. The solution was wicked off and, thereafter, air-dried before viewing under a transmission electron microscope Philips EM 208.
Data analysis
Results are expressed as mean ± standard error of the mean (n = 3). The mean values were compared using a two-way ANOVA (analysis of variance) followed by Tukey test using the statistical package for social sciences (SPSS) 10.0 for Windows.
Results
β-Secretase (BACE 1) inhibition
The ability of phenolic-rich extracts from C. annuum var. grossum to inhibit β-secretase is presented in Figure 1. The extracts were able to inhibit β-secretase in a dose-dependent manner. Furthermore, the inhibitory effect of extracts from ripe C. annuum var. grossum (2.17 ± 0.07 μg/L) was significantly (P < .05) higher than extracts from unripe C. annuum var. grossum (3.44 ± 0.11 μg/L) as shown in Table 1.

Inhibition of β-secretase (BACE 1) activity by Capsicum annumm var. grossum.
Values represent means of duplicate; n = 3. Values with the same letter are not significantly (P > .05) different.
Inhibition of Aβ 1–40 aggregation
Figure 2 depicts the inhibitory effect of extracts from ripe and unripe C. annuum var. grossum on Aβ 1–40 aggregation. Incubation of Aβ 1–40 (10 μM) with and without extracts from ripe and unripe C. annuum var. grossum resulted in an increase in fluorescence signals with a progression in incubation period. This signified assembly of the peptide monomers into aggregated forms. Aβ 1–40 incubated alone showed a significant (P < .05) increase in peptide aggregation. However, there was a significant decrease in peptide aggregation as typified by decreased fluorescent signals from Aβ 1–40 coincubated with extracts from ripe and unripe C. annuum var. grossum. Figure 2b further revealed no significant (P > .05) difference in the % inhibitory effects of the phenolic-rich extracts of both the ripe and unripe C. annuum var. grossum at a lower concentration of 1 μg/L. However, at 10 μg/L, extracts from ripe C. annuum var. grossum revealed a significantly (P < .05) higher % inhibitory ability than the unripe. TEM imaging (Fig. 2C) revealed that Aβ 1–40 incubated for 96 h at 37°C formed fibrils with extensive networks; the width of the fibrils ranged from 7.90 to 10.80 nm (data not shown). However, the presence of fibrils was completely absent when Aβ 1–40 was coincubated with C. annuum var. grossum, rather nonfibrilar aggregates, which were amorphous in nature, were otherwise observed.
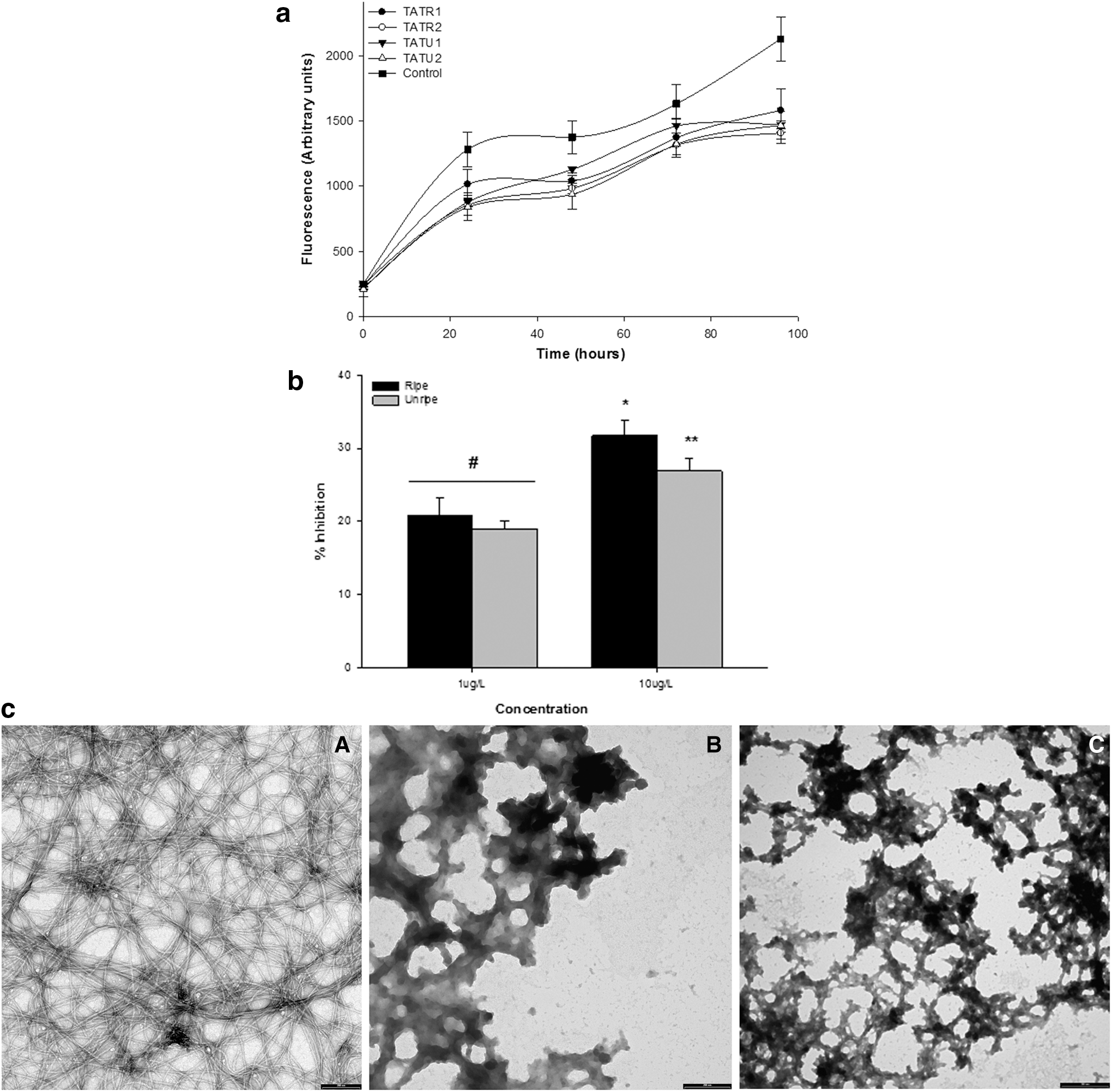
We further sought to find out if the extracts from C. annuum var. grossum could prevent further aggregation Aβ 1–40 fibrils, and the results are presented in Figure 3. Incubation of Aβ 1–40 alone for an additional 48 h resulted in continued aggregation of the peptides as demonstrated by increases in ThT fluorescent signals. However, introduction of C. annuum var. grossum extracts at the 48th hour prevented further aggregation of the peptides (Fig. 3). In addition, extract concentration at 10 μg/L resulted in significant (P < .05) decrease in ThT signals by the 96th hour compared to the 1 μg/L extract concentration where the fluorescent signals was not significantly (P > .05) different from signal observed at the 48th hour.
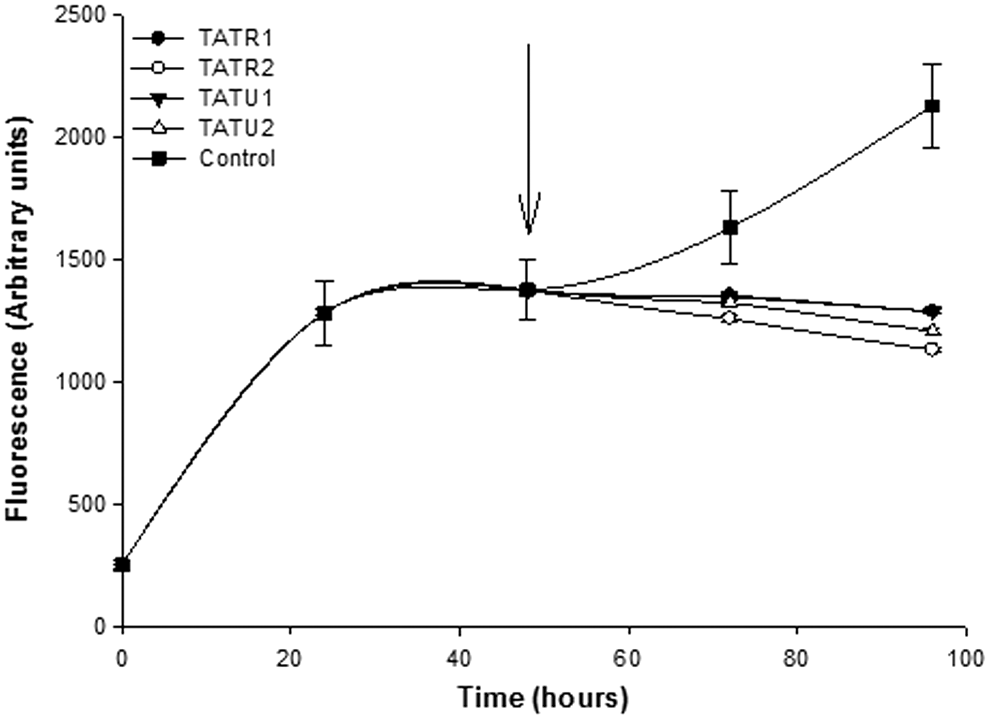
Kinetics of the disaggregation of preformed Aβ 1–40 fibrils by C. annuum var. grossum. Key: Control—Aβ 1–40 alone; TATR1—Aβ 1–40 + 1 μg/L extract of ripe C. annuum var. grossum; TATR2—Aβ 1–40 + 10 μg/L extract of ripe C. annuum var. grossum; TATU1—Aβ 1–40 + 1 μg/L extract of unripe C. annuum var. grossum; TATU2—Aβ 1–40 + 10 μg/L extract of unripe C. annuum var. grossum. Arrow indicates the time in which phenolic extracts were introduced into the reaction medium.
HPLC analysis
To predict the possible phytochemicals responsible for the observed bioactivity, HPLC-DAD profiling was carried out for the ripe and unripe C. annuum var. grossum extracts. Figure 4 and Table 2 revealed the presence of gallic acid, chlorogenic acid, rutin, isoquercitrin, quercitrin, quercetin, kaempferol, dihydrocapsaicin, and capsaicin. The presence of the flavonoids quercetin, quercitrin, isoquercitrin, and kaempferol was enhanced in the ripe fruits.
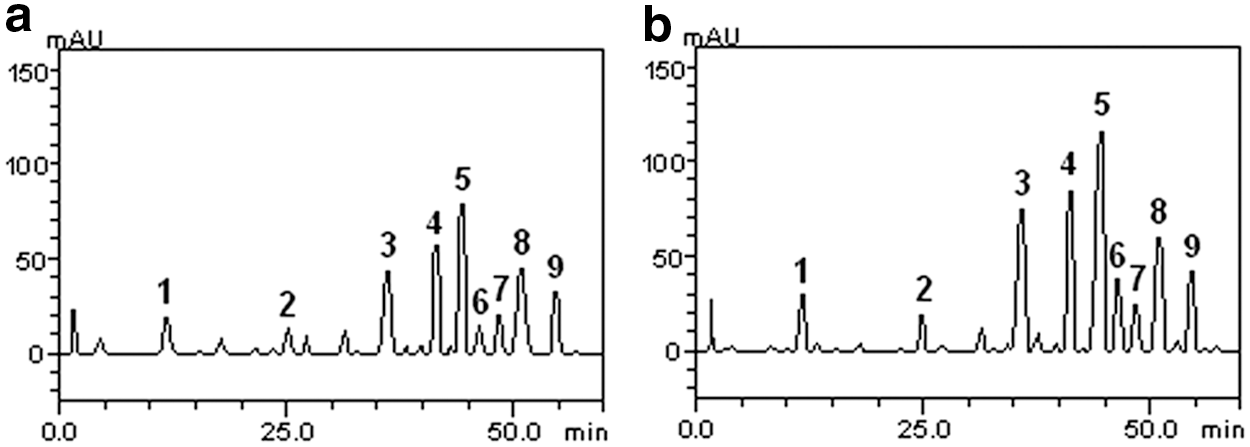
Representative high performance liquid chromatography profile of unripe
Results are expressed as mean ± standard deviation of three determinations. Means with different superscript letters are significantly different (P < .05).
Discussion
Emerging evidence suggests that dietary phytochemicals are able to exert beneficial effects in the central nervous system showing anti-aggregative and anti-neurotoxic properties in vitro. 9,21 Dietary supplementation studies in humans and animals using food extracts have highlighted their potential to influence cognition and learning presumably by protecting vulnerable neurons, enhancing existing neuronal function, or by stimulating neuronal regeneration. 21
Current trends in the therapeutic approach to AD include modulating APP processing with specific respects to β-secretase inhibition. 2,3,22 This study has been able to show that phenolic-rich extracts from the fruits of C. annuum var. grossum were able to inhibit the β-secretase activity and may be able to modulate the amyloidogenic pathway of APP processing and by this could invariably shift APP metabolism toward the non-amyloidogenic pathway. This inhibitory activity could be attributed to the presence of polyphenolic compounds present in the fruits. The inhibition of β-secretase by polyphenolic compounds from C. annuum var. grossum is consistent with previous studies by Jiminez-Aliaga et al. 23 and Choi et al. 24 where phenolic compounds such as luteolin and rosmarinic acid from Perilla frutescens, quercetin, and rutin all inhibited β-secretase. Considering that the present β-secretase inhibitors used in AD management/treatment are peptidomimetics of the β-cleavage site of APP, 25 which are usually very large molecules and as such may be unable to cross the blood–brain barrier (BBB), this present study becomes relevant in that polyphenols are small compounds, which are relatively advantageous for crossing the BBB. 23,26,27
The inhibition of Aβ aggregation and disaggregation of preexisting fibrils is equally regarded as a promising therapeutic strategy in the prevention and management of AD. Results from this study have shown that phenolics from unripe and ripe C. annuum var. grossum at both concentrations (1 and 10 μ g/L) were able to inhibit β-amyloid fibril formation at the later growth phase. This is suggestive of an interference in the structural conversion of the β-amyloid random coils to its β-structures or in subsequent addition of the peptide monomers to existing fibrillar forms. Several low molecular weight molecules, particularly scaffolds with aromatic or heteroaromatic rings, have been identified as potent in their ability to perturb amyloid fibril assembly. 28,29 Their ability to inhibit the assembly of amyloid protein in its fibrillar form is due to nonspecific interactions with hydrophobic moieties in the protein structure and also interactions of the hydrogen bonds and the aromatic rings with the peptidic carbonyl backbone of the peptide structure. 23 This could give some insight into the mechanisms involved in the inhibitory activity of the phenolic-rich extracts from C. annuum var. grossum on Aβ aggregation and its ability to destabilize aggregated forms. Furthermore, polyphenols such as curcumin have been reported to suppress Aβ aggregation through hydrogen bonding to the Aβ structure at Gln, 15 Glu, 22 and Asp 23 positions. The region between Gln 15 –Ile 32 is predicted to be an intramolecular β-sheet in Aβ assemblies. Any molecular associations however weak between the peptide and phenolic molecule within this region could result in the inhibition of amyloid assemblies and also in its disaggregation. 30
Ripe pepper fruits showed a higher inhibitory effect on β-secretase and amyloid aggregation, as well as a higher ability to disaggregate preformed fibrils than the unripe fruits. This can be attributed to the higher phenolic content (particularly quercetin and its derivatives) of the ripe fruits (Table 2) as the phenolic content of plant foods has a positive correlation to their biological activity. This increased phenolic content upon ripening of the fruits could be attributed to bioconversion of phytochemicals possibly to phenolics with progression in the maturity of the fruits. Ripening of fruits or other plant tissues involves a series of complex reactions that leads to changes in the phytochemistry of the plants. 31 Polyphenols, which are products of the shikimate and phenylpropanoid pathways, 32 are observed to increase with fruit maturity and eventual ripening. This phenomena of increase in phenolic content as ripening occurs in pepper fruits is consistent with earlier studies by Conforti et al., 16 Oboh and Rocha, 33 and Loizzo et al., 34 where increases in total phenol content were observed after the fruit had attained full maturity. However Menichini et al. 35 reported a contrary observation in which there was a loss in phenolic content of some peppers with maturation. It is also interesting to note that the most abundant phenolic constituents present were quercetin and its glycosides namely quercitrin (quercetin 3-O-α-L-rhamnoside), isoquercitrin (quercetin 3-O-glucopyranoside), and rutin (quercetin 3-O-rutinoside). This finding was consistent with an earlier study by Materska and Perucka, 36 where quercetin was also the most predominant phenolic constituent in Capsicum spp.
In conclusion, bell pepper extracts were able to inhibit β-secretase activity and aggregation of Aβ 1–40 peptides. Extracts from ripe fruits showed a higher activity than the unripe. These inhibitory properties could be ascribed to the presence of their constituent phytochemicals. This is indicative of the potential of C. annuum var grossum as a possible dietary means of managing AD. Further in vivo studies of C. annuum var grossum are recommended to further the understanding of its action on β-secretase activity and aggregation of Aβ 1–40.
Footnotes
Author Disclosure Statement
No competing financial interests exist.