
Abstract
Natural bioactive compounds are considered an excellent alternative strategy for developing effective, safe, and cost-effective antiobesity agents. The aim of this study was to investigate if combinations of soy bean genistein (G), green tea epigallocatechin-3-gallate (E), and/or grape resveratrol (R) at low dosages synergistically inhibit preadipocyte differentiation both in 3T3-L1 cells and human primary preadipocytes (HPAs). Our results show that combinations of G, E, and/or R additively inhibited preadipocyte differentiation (39–56% of control) both in 3T3-L1 cells at 30 μM and HPAs at 15 μM, while the individual compounds have no antiadipogenic effect at the selected concentrations. We also observed similar patterns that combinations of G, E, and/or R additively reduced protein expressions of peroxisome proliferator-activated receptor gamma (PPAR-γ) and CCAAT binding proteins alpha (C/EBP-α), the two key preadipocyte differentiation regulators, both in differentiated 3T3-L1 cells and HPAs. Moreover, combined G, E, and/or R attenuated protein expressions of fatty acid binding protein 4 and perilipin, two PPAR-γ/C/EBP-α downstream molecules in fat drop development in a very similar pattern, in inhibiting differentiation in preadipocytes. This combined antiadipogenic effect of G + E + R is additive, not synergistic according to our results and the Median-Effect Principle. In addition, we found that a lower concentration (15 μM) of G, E, and/or R is required in HPAs than the concentration (30 μM) needed in 3T3-L1 cells, to exert the combined antiadipogenic effect. These data suggest that combinations of G, E, and/or R intake or soy bean, green tea, and/or grape simultaneous consumption may prevent obesity in human being.
Introduction
H
Changing lifestyle such as increasing physical activity and reducing energy intake has successfully reduced body weight; 5 however, alternative methods are required to control the body weight because increasing physical activity and/or limiting energy intake is extremely hard for many people. Natural plant-derived compounds have been recently considered an excellent alternative strategy for developing effective, safe, and cost-effective antiobesity agents because of the potential hazardous side effect and high cost of current antiobesity drugs. 5 A variety of natural products, including crude extracts and isolated compounds from plants including curcumin, capsaicin, gingerol, epigallocatechin-3-gallate (E), genistein (G), resveratrol (R), and quercetin, 6 have been studied in treating obesity.
However, the required concentrations (12.5–100 μM) of these antiobesity compounds in the in vitro studies are much higher than the circulating levels (<10 μM) of these compounds by consuming plant-derived food or pure chemical supplements. 7 –10 Our strategy to solve this problem is to combine several antiobesity compounds from different plants to prevent obesity. Actually, recent studies have shown that G enhances the circulating and cellular bioavailability of E, 11 and combinations of G, E, or quercetin synergistically inhibited cancer cell proliferation 12 and improved osteoporosis. 13,14 Moreover, combined bioactive compounds synergistically affect adipocytes. 15 –17 Therefore, in this study, we hypothesized that combinations of several bioactive compounds at low dosages may synergistically inhibit preadipocyte differentiation. The specific objectives are as follows: (1) if combinations of genistein (G), epigallocatechin-3-gallate (E), and/or resveratrol (R) at relative low concentrations synergistically inhibit cell differentiation in 3T3-L1 preadipocytes and human primary preadipocytes (HPAs); (2) the synergistic antiadipogenic effect of combinations of G, E, and/or R in preadipocytes is through regulating peroxisome proliferator-activated receptor gamma (PPAR-γ) and CCAAT binding protein alpha (C/EBP-α) pathways, the key preadipocyte differentiation regulators. 4
Materials and Methods
Reagents
3T3-L1 preadipocytes and HPAs were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). Dulbecco's modified Eagle's medium (DMEM) and penicillin-streptomycin-glutamine were obtained from Gibco (Grand Island, NY, USA), and fibroblast basal medium and fibroblast growth kit were obtained from ATCC. Trypsin/EDTA and fetal bovine serum (FBS) were obtained from LONZA (Walkersville, MD, USA) and ATLANTA Biologicals (Lawrenceville, GA, USA), respectively. TACS MTT cell proliferation assay kit was purchased from Trevigen (Gaithersburg, MD, USA). Different primary antibodies β-actin, PPAR-γ, CEBP-α, fatty acid binding protein 4 (FABP4), and perilipin were purchased from Cell Signaling Technology (Beverly, MA, USA). Secondary antibody anti-rabbit IgG was obtained from Millipore (Darmstadt, Germany). G (Glycine max, 98% pure), E (Green tea, ≥95% pure), and R (red grape) were purchased from Sigma (St. Louis, MO, USA). Oil Red O solution was from Electron Microscopy Sciences (Hatfield, PA, USA), and phosphate-buffered saline (PBS) and 2-propanol were from Fisher Scientific (Pittsburgh, PA, USA). Stock solutions of 100 mM G, E, and R in dimethyl sulfoxide (DMSO) were stored at −80°C before use.
3T3-L1 cell culture
Cells were cultured in 12-well plates with DMEM with 10% FBS and 1% P/S (culture medium); 2 days after reaching confluence (day 0), the 3T3-L1 cells were differentiated in the culture medium plus 10 μg/mL insulin, 1 μM dexamethasone, and 0.5 mM 3-isobutyl-1-methylxanthine (differentiation medium) with or without treatments as shown in Tables 1 and 2 for 72 h modified from previous study. 17 The cells were then maintained in the culture medium plus 10 μg/mL insulin (differentiation maintenance medium) with or without treatments for seven more days. Differentiation maintenance medium with or without treatments was changed after every 2 days. On day 10, cells were either stained with Oil Red O or collected for Western blot. An equivalent amount of DMSO alone (as same as the G + E + R (GER) combination) was added in the differentiated control and nondifferentiated control, complementary DMSO was added to the individual or two compounds combined treatments to make sure each well has the same amount of DMSO. The concentration, 30 μM of individual compound, was selected based on our preliminary study and previous studies 13,14,18 that an individual compound has no significant effect in inhibiting differentiation in 3T3-L1 cells.
DMSO, dimethyl sulfoxide; GER, G + E + R.
HPA cell culture
HPAs seeded in 12-well plates were maintained at 37°C in a humidified atmosphere of 5% CO2 in a fibroblast basal medium with the fibroblast growth kit, biotin 33 μM and panthothenate 17 μM (culture medium). Upon reaching 100% confluence (day 0), the media were replaced with the differentiation medium (culture medium plus 10 μg/mL insulin, 1 μM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine) with or without treatments as shown in Table 1, but at 15 μM for all compounds for 6 days, the differentiation medium with or without treatments was replaced on day 3. On day 6, the cells were maintained in a differentiation maintenance medium (culture medium plus 10 μg/mL insulin) with or without treatments for nine more days. The differentiation maintenance medium with or without treatments were replaced every 3 days. 19 On day 15, cells were either stained with Oil Red O or collected for Western blot. An equivalent amount of DMSO alone (as same as the GER combination) was added in the differentiated control and nondifferentiated control, complementary DMSO was added to the individual or two compounds combined treatments to make sure each well has the same amount of DMSO. The concentrations (15 μM) of chemicals were based on preliminary studies that an individual compound has no significant effect in inhibiting differentiation in HPAs.
Cell viability assay
3T3-L1 or HPA cells (1 × 105 cells per mL) were seeded in 12-well plates with/without different treatments as above described for 48 h. Treated cells were incubated with 100 μL MTT reagent per well for 4 h till purple dye was visible. Five hundred microliters of detergent reagent was added into each well and incubated in the dark for 4 h. The relative cell viability was determined by the absorbance at 570 nm using Synergy H1 hybrid reader (BioTek Instruments, Inc. Winooski, VT).
Oil Red O stain
On day 10 for 3T3-L1 cell and day 15 for HPAs, after removing the medium, the cells were washed with 1 × PBS twice and stained with Oil Red O modified from previous studies. 17,19 In brief, the cells were fixed with Baker's formalin (10 mL of 37% formaldehyde, 10 mL of a 10% calcium chloride solution, and 80 mL of distilled water) for 1 h at 4°C and then washed twice using distilled water. The cells were stained with Oil Red O solution (0.5 mL/well) for 15 min and then washed thrice with distilled water. Accumulated lipid was dissolved in 0.5 mL 100% isopropanol for 1 h at room temperature with continuous shaking. The lipid accumulation was measured using a Synergy H1 microplate reader (Bio-Tek Instruments, Inc., Winooski, VT) at 490 nM.
Western blot
3T3-L1 or HPA cells were differentiated and treated as described above in different experiments. On day 10 for 3T3-L1 cells and day 15 for HPAs, after removing the medium, 2 × sample buffer was added into the wells directly and collected into 1.5 mL tubes. After being centrifuged at 2400 g for 5 min, samples were sonicated for 15 sec (three times, 5 sec each time on ice after every 5-sec sonication), and were centrifuged again at 2400 g for 5 min. Samples were heated at 95°C for 5 min and kept in −20°C for storage. Proteins in samples were separated using 10% polyacrylamide gel electrophoresis at 100 mA for 2 h and then transferred to nitrocellulose membranes at 100 mA for 2 h. The membrane was blocked for 1 h with phosphate-buffered saline (TBST) containing 5% skim milk at room temperature. After washing thrice with TBST, the membrane was incubated overnight with relevant primary antibody at 4°C. The membrane was washed thrice with TBST and then incubated with secondary antibody for 1 h at room temperature. Specific bands were detected by SuperSignal West Dura chemiluminescence (ThermoFisher Scientific, USA) and visualization was performed by exposure of the membranes to X-ray films. Band intensities were quantified by ImageJ software (National health institute, USA).
Statistical analyses
All data were analyzed using one-way ANOVA. Values (means ± standard error) of three separate experiments are expressed as percentage of differentiated control. Treatment differences, if significant, were subjected to Tukey's multiple comparison tests and a value of P < .05 indicates a significant difference between groups.
Results
Combined G, E, and/or R significantly inhibited differentiation and fat accumulation in 3T3-L1 cells
After being treated with individual and various combinations of G, E, and/or R at 30 μM in differentiated 3T3-L1 cells, we found that combinations (GE, ER, GER) significantly inhibited lipid accumulation in differentiated 3T3-L1 cells, while individual compounds have no antiadipogenic effect at the selected concentrations (Fig. 1). Lipid size and area of lipid droplets in treatments GE, ER, and GER were significantly less than individual compound-treated cells, particularly, a combination of three compounds (GER) has a stronger antiadipogenic effect than combinations of two compounds (ER or GE) under microscope (Fig. 1a). From Figure 1b, while individual compounds G, E, and/or R at 30 μM did not significantly inhibit lipid accumulation (E, 96.5% of control, P > .05; R, 94.0% of control, P > .05; and G, 91.5% of control, P > .05), lipid accumulation was significantly decreased when two compounds were added (ER,73.0% of control, P < .05; and GE, 62.0% of control, P < .05). Moreover, when the three compounds (GER) were added together, lipid accumulation was extremely decreased to 39.3% of control (P < .01). We did not observe toxicity of these individual or combined compounds on the cells at the selected concentrations using the MTT cell viability assay (data not shown). To eliminate the effect of DMSO, the DMSO level was balanced and each well has the same amount of DMSO in the entire differentiation assay.
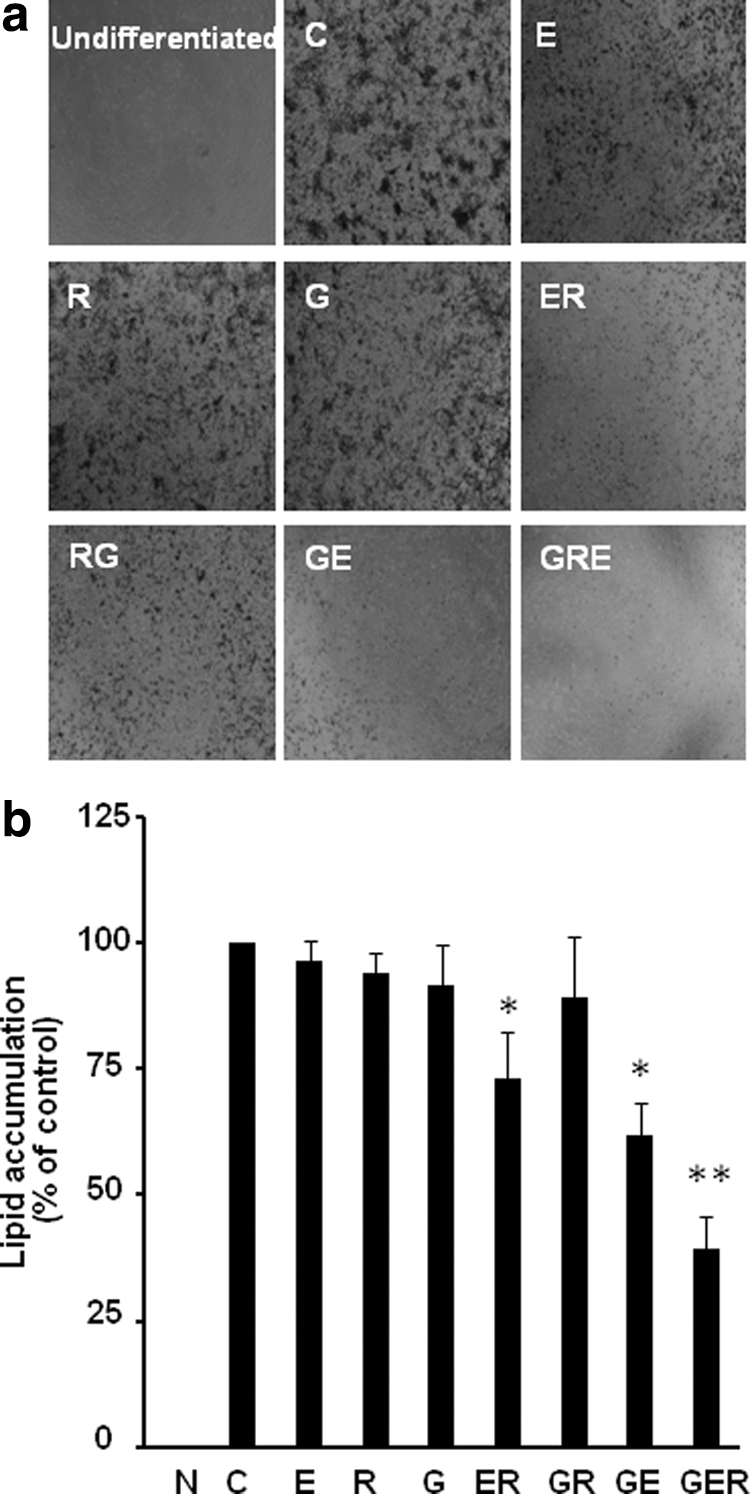
Combined genistein (G), EGCG (E), and/or resveratrol (R) inhibited differentiation in 3T3-L1cells. Representative images of lipid accumulation
Combined G, E, and/or R significantly suppressed PPAR-γ and C/EBP-α protein expressions in differentiated 3T3-L1 cells
Because of the critical roles of PPAR-γ and C/EBP-α in adipogenesis, we investigated the protein expressions of these two molecules with/without treatments in 3T3-L1 cells. We found that treatments of E slightly reduced the PPAR-γ protein expression (90.9% of control, P < .05) (Fig. 2a); however, two-compound combinations reduced PPAR-γ protein expression much more (ER, 30.3% of control, P < .01; and GE, 38.6% of control, P < .01), and three compounds together decreased most of PPAR-γ protein expression (GER, 13.9% of control, P < .01). Similarly, treatments of E slightly reduced the protein expression of C/EBP-α (83.2% of control, P < .05) (Fig. 2b); when two compounds added together, the C/EBP-α protein expression was significantly reduced (ER, 26.6% of control, P < .01; GE, 48.2% of control, P < .05), and treatment of GER reduced most of the C/EBP-α expression (GER, 18.9% of control, P < .01). Interestingly, GR did not reduce the protein expressions of PPAR-γ and C/EBP-α, a very similar pattern in lipid accumulation in 3T3-L1 cells.
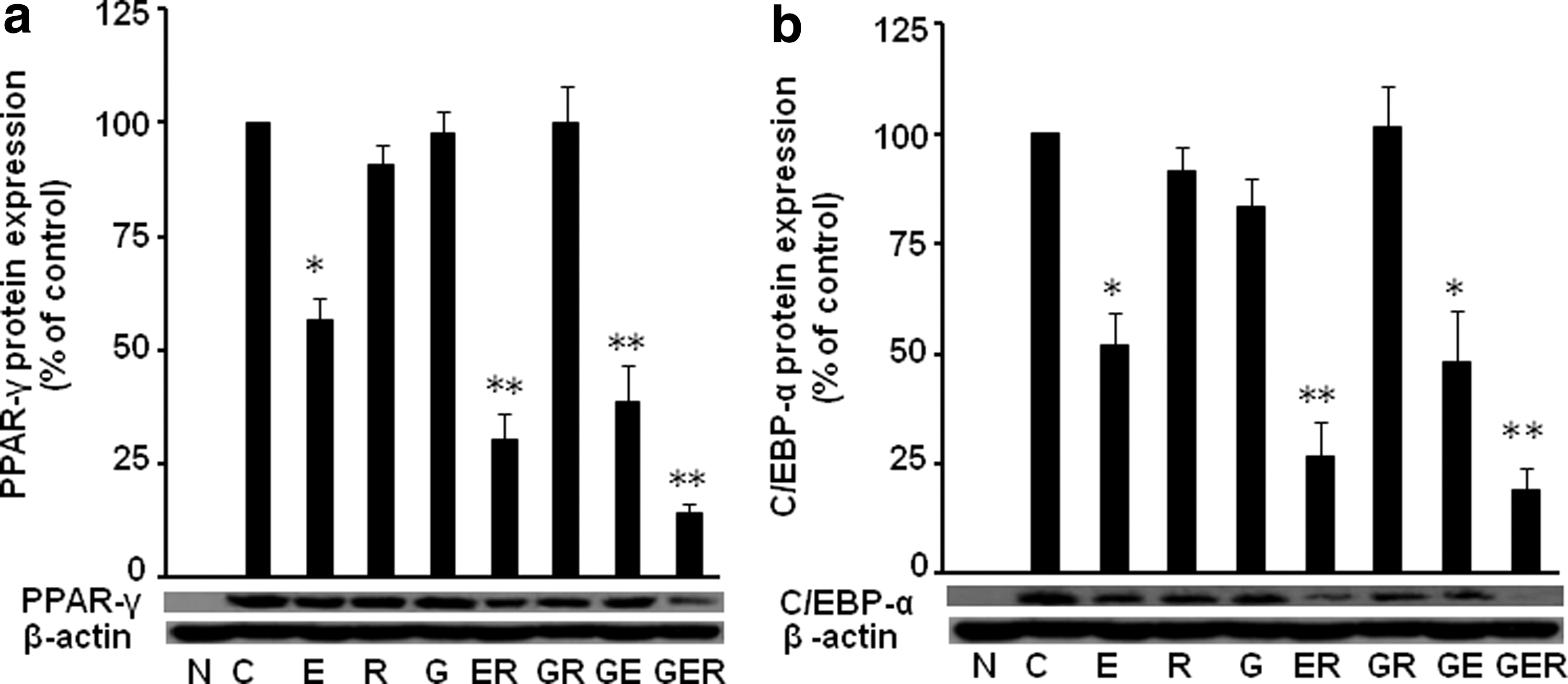
Combinations of G, E, and/or R suppressed PPAR-γ and C/EBP-α protein expressions in 3T3-L1 cells. The protein expression levels of PPAR-γ
Combined G, E, and/or R significantly attenuated downstream protein expressions in differentiated 3T3-L1 cells
We further evaluated the PPAR-γ and C/EBP-α downstream molecules FABP4 and perilipin protein expression in differentiated 3T3-L1 cells. Similarly, we found that individual treatment slightly reduced the protein expression of FABP4 (E, 74.0% of control, P > .05; R, 55.0% of control, P > .05; and G, 63.0% of control, P < .05) (Fig. 3a), but two-combined compounds GE, ER, and GR significantly reduced the FABP4 expression (ER, 23.0% of control, P < .05;GR, 15.0% of control, P > .05; GE, 72.0% of control, P > .05), and three compounds together reduced most of the FABP4 expression (GER, 12.0% of control, P < .01).
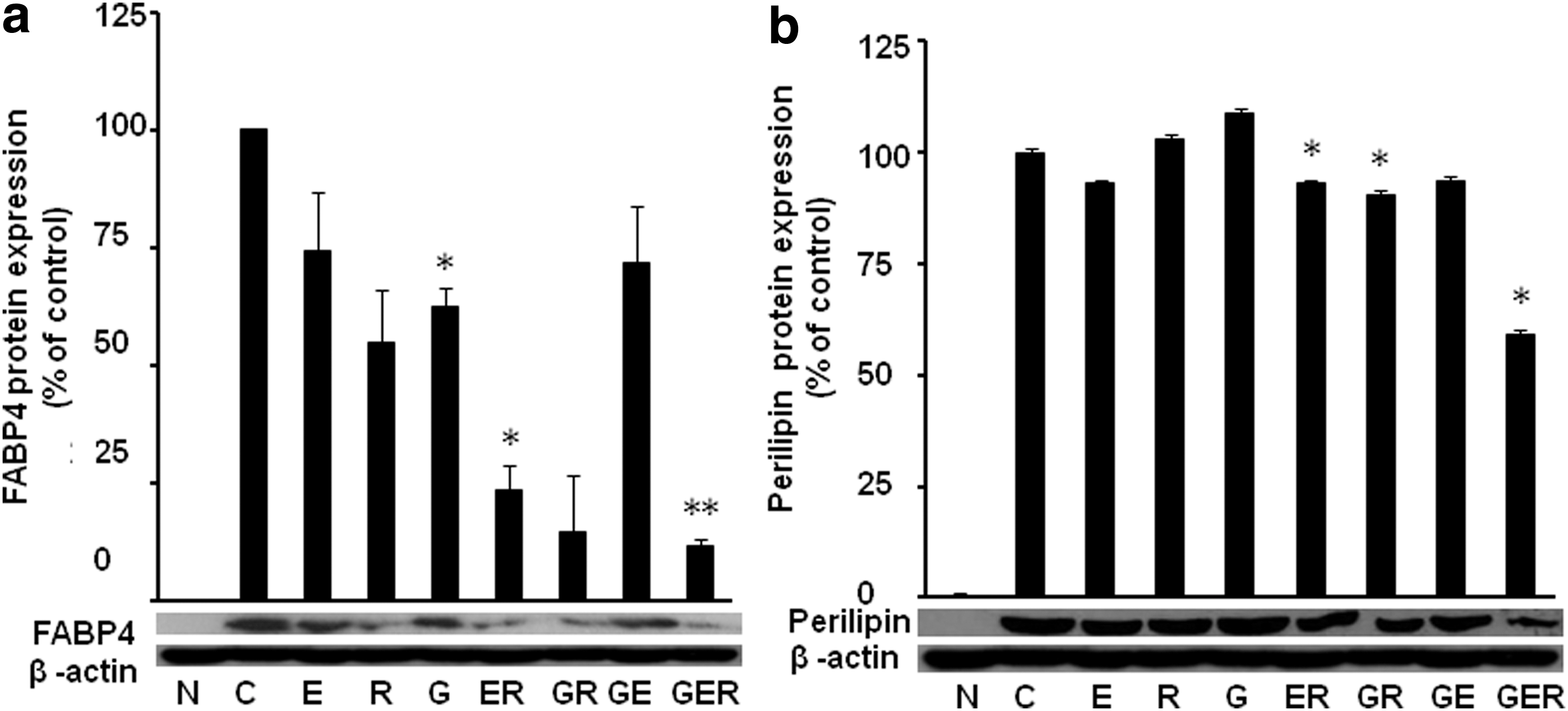
Combinations of G, E, and/or R attenuated protein expression of FABP4 and perilipin in 3T3-L1 cells. The protein expression levels of FABP4
We also found that individual treatment did not affect the protein expression of perilipin (Fig. 3b), but two-combined compounds ER (93.0% of control, P < .05) and GR (90.5% of control, P > .05) inhibited perilipin expression, and three compounds together reduced most of the protein expression (GER, 59.0% of control, P < .05).
Combined G, E, and/or R significantly restrained differentiation and relevant proteins at lower concentration in HPAs
Since 3T3-L1cells are rodent cell lines, and high concentrations of phytochemicals are required to inhibit adipogenesis, we would like to know whether HPAs are more sensitive than 3T3-L1cells in the antiadipogenic effect of these phytochemicals. Indeed, after treatment in differentiated HPAs with individual or various combinations of E, G, and/or R at 15 μM based on our preliminary experiments, we found that combinations of ER and GER significantly inhibited fat accumulation in differentiated HPA cells, while individual compounds had no antiadipogenic effect at the selected concentrations (Fig. 4). This concentration (15 μM) in HPAs is just half the concentration (30 μM) in 3T3-L1cells, confirming our hypothesis that HPAs are more sensitive than 3T3-L1cells in the antiadipogenic effect of these phytochemicals. This low concentration is much closer to the physiologically achievable blood levels (about 10 μM) after dietary intake of relevant foods or supplements. 20,21 Lipid accumulation in treatments ER and GER was significantly less than individual compound-treated cells, particularly, combinations of three compounds have more antiadipogenic effect than combinations of two compounds under microscope (Fig. 4a). We did not observe toxicity of these individual or combined compounds on the cells using the MTT cell viability assay (data not shown).
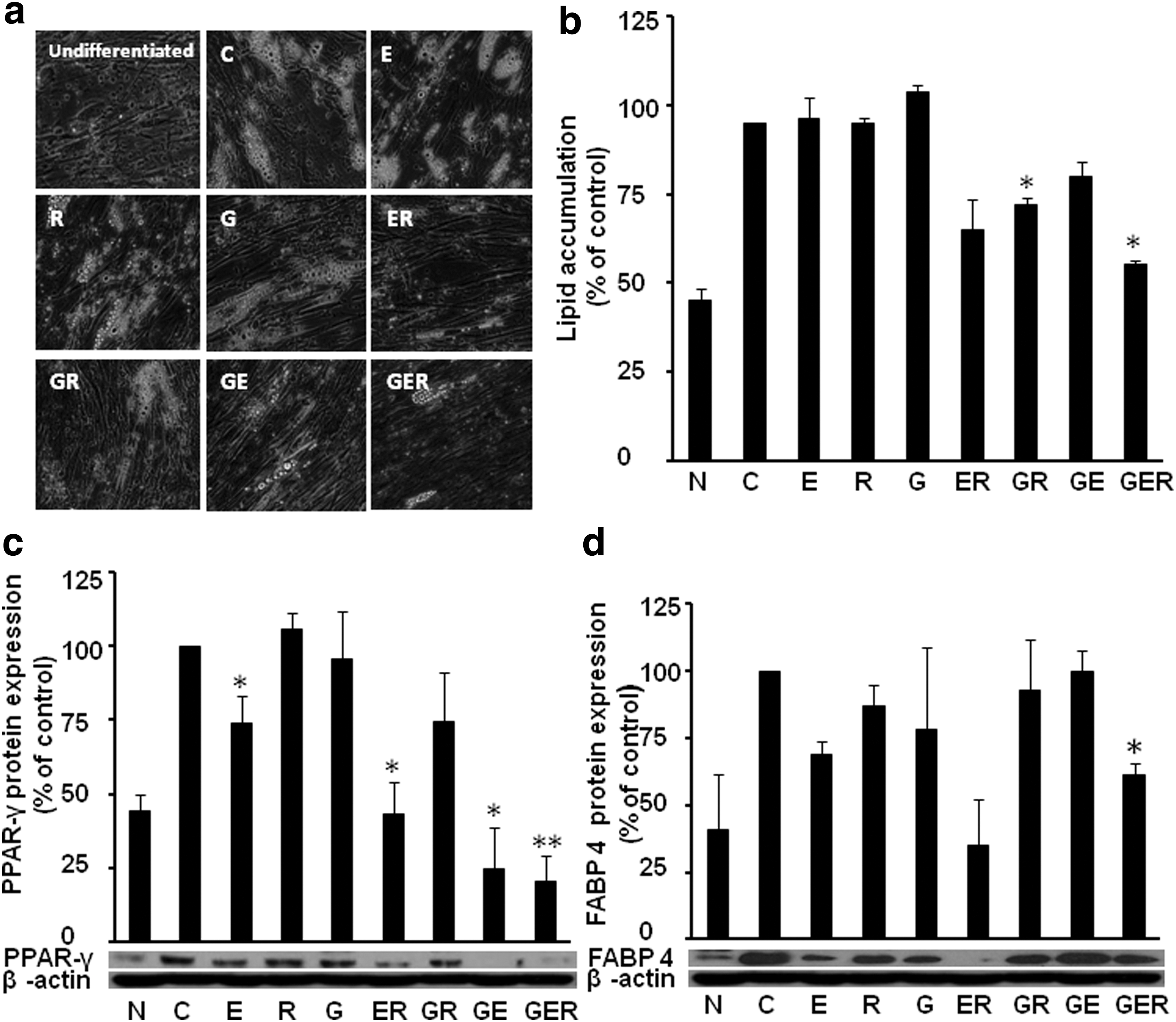
Combined G, E, and/or R restrained differentiation in human primary preadipocytes (HPAs). Representative images of lipid accumulation
We further found a very similar pattern of protein expressions of PPAR-γ (Fig. 4c) and FABP4 (Fig. 4d); GER had the best inhibitory effect than the two-compound combinations, while individual compounds had very limited or no effects in inhibiting these proteins. However, PPAR-γ protein expression was decreased by ER (43.0% of control, P < .05), GR (74.7% of control, P > .05), and GE (24.7% of control, P < .05), and three compounds together significantly decreased the PPAR-γ protein expression (GER, 20.1% of control, P < .01). In case of FABP4, we found the protein expression was not significantly reduced by individual compounds and two-combined compounds (E, 87.0% of control, P > .05; R, 69.0% of control, P > .05; G, 78.0% of control, P > .05; ER, 35.0% of control, P > .05; GR, 93.0% of control, P > .05; and GE,100.0% of control, P > .05). However, three compounds combined together significantly reduced the FABP4 protein expression (GER, 61.0% of control, P < .05).
The antiadipogenic effect of combined G, E, and/or R is additive, not synergistic in 3T3-L1 cells
Since we use 30 μM of E, G, and/or R in the above individual and combined treatments, we would like to investigate whether the combined effects are synergistic or additive. We select 90 μM individual E, R, and G to compare the effect on lipid accumulation in differentiated 3T3-L1 cells. We found that when cells were treated with G, E, and R individually at 90 μM, lipid accumulation was significantly prevented by R (3.0% of control, P < .01) and G (7.0% of control, P < .01) (Fig. 5a), and G30E30R30 (44.0% of control, P < .01), indicating that G30E30R30 is not as effective as G90 and R90 in inhibiting lipid accumulation. The protein expression of PPAR-γ (Fig. 5b), C/EBP-α (Fig. 5c), and FABP4 (Fig. 5d) are in line with the lipid accumulation. These results suggest that the combined antiadipogenic effect of GER is a kind of summation or additive, not synergistic.
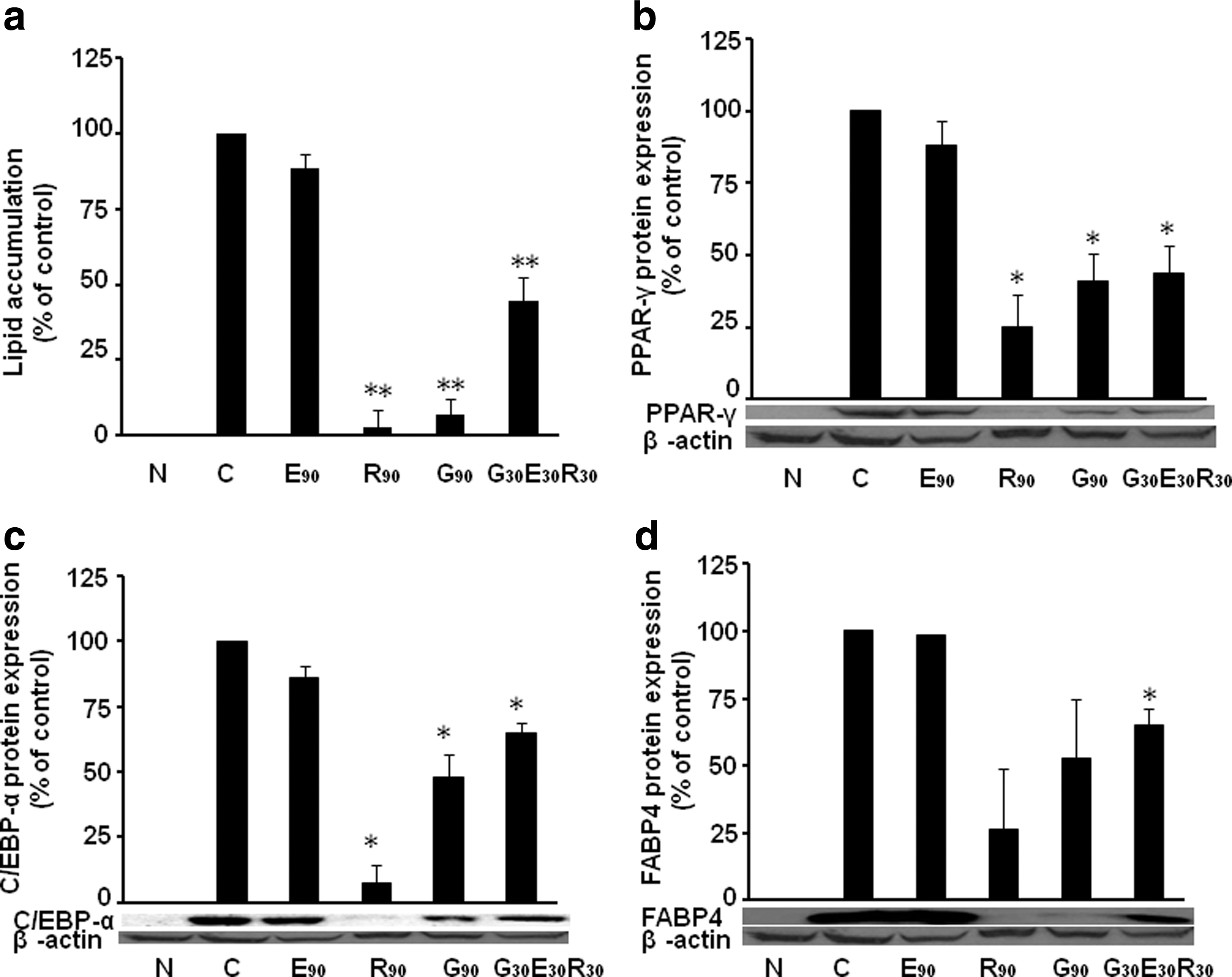
The antiadipogenic effect of combined G, El, and/or R is additive, not synergistic in 3T3-L1 cells. Lipid accumulation
Discussion
This is the first report to our knowledge that combinations of G, E, and/or R, the three most well-known food-derived chemicals majorly from soybean products, green tea and grapes/wine, additively inhibited cell differentiation at low concentration, while the individual G, E, and R did not have the antiadipogenic effect both in 3T3-L1 cells and HPAs. This antiadipogenic effect of combinations of G, E, and/or R was companied by the reduced protein expressions of PPAR-γ and C/EBP-α, the two key preadipocyte differentiation regulators, both in differentiated 3T3-L1 cells and HPAs. Moreover, combined G, E, and/or R reduced the protein expression of FABP4, a key molecule in fat drop accumulation in a very similar pattern of these compounds, in inhibiting differentiation in 3T3-L1 cells. In addition, we found that a lower concentration (15 μM) of G, E, and/or R is required in HPAs than the concentration (30 μM) needed in 3T3-L1 cells, to exert the combined antiadipogenic effect. These data suggest that combinations of G, E, and/or R intake or simultaneous consumption soy bean, green tea, and/or grape may prevent obesity in human beings.
Indeed, combining multiple natural bioactive compounds on human disease research is increasing recently. For instance, a recent study reported that vitamin D combined with G, quercetin, and R reduced body weight gain and a significant decrease in bone marrow adipocytes in rats. 22 Combinations of G, E, and/or R can synergistically inhibit cancer cell proliferation, 23 improve osteoporosis., 13 and suppress oxidative stress. 24 The major reason for using several combined compounds is that the high dosages working in cells and animals of individual compounds cannot be reached in humans by consuming relevant foods or supplements. Indeed, plasma G concentration is lower than 4 μM in human consumption of bean-based food products, and the peak plasma G concentration is about 10 μM in humans administrated G (8 mg/kg body weight), 9,25 while G only exerts antiobesity effect at 10–100 μM as shown above. 26 This is further supported by recent studies that the antiobesity effect of plant-derived compounds are still controversial in human studies, particularly, there are no clinical studies showing G 26 or R 27 decreases fat pad weights. Therefore, combining several antiobesity compounds at low level (physiological achievable) may provide a practical approach to prevent obesity by consuming a variety of foods with antiobesity bioactive compounds.
It was reported that the total maximum circulating G, E, and R levels in humans and animals consuming products of soy bean, green tea, and grape/wine or supplements are 0.74–10.0 μM, 20,21 0.7–2.4 μM, 8,28 and 4–5.8 μM, 29 respectively. Moreover, concentrations of G 30 and E 31 in tissues may be even higher than the concentrations in blood. For instance, G in rat brain reached 8.9 μM, while the highest level of G in blood is only 8.0 μM in the same study. 30 In addition, a second, equal administration to mice after a 6-h interval enhanced tissue levels of E in blood, brain, liver, pancreas, bladder, and bone 4–6 times above those after a single administration. 31 It must be noted that free genistein accounts only 5–26% of total G present in plasma in humans, 32 and G conjugates in the serum are reported to be lesser biologically active than free G, 33 while our in vitro studies use free G, E, and/or R. Therefore, the concentrations of G, E, and/or R that produced biological effects, observed in this study, in HPAs (15 μM) may be attained in the plasma and tissues in humans following dietary supplementation. Moreover, a previous study found that G inhibited HPA differentiation at 6.25 μM, 17 which is much lower than the concentration (15 μM) of G without an antiadipogenic effect used in our study. This difference may come from the different origin and characteristics of primary HPAs in these two studies.
Besides the antiadipogenic effect of G, E, and/or R by suppressing PPAR-γ/C/EBP-α pathway as aforementioned, accumulating evidence shows that G, E, and/or R also lower levels of reactive oxygen species, 34,35 inflammation, 36 and apoptosis, 21 important contributors of obesity and obesity-related diseases. Furthermore, G, 37 E, 37 and R 38 inhibit cell differentiation by activating AMP-activated kinase (AMPK), a fuel gauge regulating energy balance. Activated AMPK may regulate the PPAR-γ activity and expression. 39 While these three compounds share these common cellular and molecular approaches in regulating adipogenesis, G enhances the circulating and cellular bioavailability of E, 11 and R can directly activate sirtuin (SIRT1), which can inhibit adipogenesis as a PPAR-γ corepressor. 40 Moreover, both G and R are ligands of estrogen receptors (ERs), and activated ERs can downregulate the PPAR-γ activity and expression. 40 However, the antiadipogenic effect of E may depend on phosphoinositide 3-kinase (PI3K)/Akt and mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) kinase pathways 41 and wingless-type MMTV integration site (WNT)/β-catenin pathway. 42 Similarly, besides ERs and PPAR-γ/C/EBP-α, the antiadipogenic effect of G may also be mediated by p38 MAPK and janus-activated kinase 2 (JAK2) signaling pathways. Therefore, the antiobesity effect of combinations of G, E, and/or R may be mediated by the common or different signaling pathways, including AMPK, ERs, SIRT1, PI3K, ERK, WNT, JAK2, P38 MAPK, and PPAR-γ/C/EBP-α. More studies are ongoing now and will be published in a separate article.
3T3-L1 cells are widely used to study preadipocyte differentiation because this cell line provides a homogeneous population with virtually all cells being at the same stage. However, 3T3-L1 preadipocytes are embryonal in origin, are aneuploid, and undergo early clonal expansion during differentiation in culture. 43 HPAs, including both preadipocytes and fibroblast-like cells isolated from human adults, are diploid and differentiate in culture without a clonal expansion phase. 44 These differences may contribute to the different sensitivity of compounds in inhibiting differentiation in our study that, 30 μM of G, E, and/or R were required in 3T3-L1 cells, but only 15 μM of G, E, and/or R were needed in HPAs. This observation is in line with a recent study that combined G, quercetin, and R and synergistically inhibited preadipocyte differentiation. 17 Moreover, 3T3-L1 cells and HPAs react different in the apoptosis regulation study. 19 Thus, this lower concentration requirement in HPAs further supports our hypothesis that combination of multiple natural compounds might be an appropriate approach to prevent obesity. Moreover, the animal studies using obese mice will be reported in the following publication.
There are three forms of interactions, including additive, synergistic, and antagonistic, when two or more phytochemicals are put into the same system. There is little information of possible interactions when E, R, and/or G are added together, although these phytochemicals naturally occur as complex mixtures. To quantitatively analyze the combined effects of multiple drugs, Chou and Talalay developed the combination index (CI) based on the Median-Effect Principle three decades ago: 45,46 CI = (D)1/(DX)1+(D)2/(DX)2, where (D)1 and (D)2 are the doses of drug1 and drug2, respectively, in the combination system; (DX)1 and (DX)2 are the doses of drug 1 and drug 2 alone, respectively. For data analysis of combinations, CI <1, CI = 1, and CI >1 indicate synergistic, additive, and antagonistic effects, respectively. Although combined GER at 30 μM in 3T3-L1 cells and 15 μM in HPAs significantly inhibited cell differentiation and suppressed critical molecules expression, while individual E, R, and G does not have these effects at these levels, the CI value of GER at 50% inhibitive effect calculated using this equation is 1.3, the combined effect is antagonistic according to Chou's theory, and this CI has been tested in thousands of studies. 45 This controversy has to be studied in the future.
Footnotes
Acknowledgments
This work was supported by National Institute of Food and Agriculture of United States Department of Agriculture (2011-38821-30965 to H.S.).
Author Disclosure Statement
No competing financial interests exist.