
Abstract
Endotoxin tolerance is a phenomenon where exposure of innate immune cells to lipopolysaccharide (LPS) induces a refractory state to subsequent endotoxin exposures. The goal of this study was to investigate if Spirulina platensis organic extract (SPE) induces an endotoxin tolerance-like state. We used splenocytes and peritoneal macrophages from C57BL/6J mice fed a high-fat/high-sucrose (HF/HS) control or a HF/HS diet containing 0.25% (w/w) SPE for 16 weeks for ex vivo LPS stimulation and endotoxin-tolerant (ET) macrophages to evaluate the effects of SPE on endotoxin tolerance. Cells from SPE-fed mice displayed significantly less expression of proinflammatory genes than those from control mice. ET macrophages were produced in vitro by incubating RAW 264.7 macrophages with low-dose LPS to determine the energy phenotype of naive, SPE-treated, and ET macrophages. Compared to naive macrophages exposed to a high-dose LPS (100 ng/mL) for the first time, ET macrophages showed significantly less proinflammatory gene expression after LPS stimulation, which was also observed with SPE treatment. Consistently, nuclear translocation of p65 was markedly reduced in both ET- and SPE-treated macrophages on LPS stimulation with increase in nuclear protein levels of p50 and B cell lymphoma 3-encoded protein. In conclusion, the anti-inflammatory effect of SPE is at least partly attributable to the induction of an endotoxin tolerance-like state in macrophages, which shares common characteristics of macrophage endotoxin tolerance.
Introduction
I
Endotoxin tolerance is a phenomenon where prior exposure of innate immune cells to LPS leads to a refractory state to subsequent LPS exposure. 5 Endotoxin tolerance is characterized by the upregulation of negative regulators of TLR4 signaling, such as p50 homodimers that inhibit nuclear factor-κB (NF-κB) p65/p50 heterodimer activity. 5 –7 In addition, B cell lymphoma 3-encoded protein (BCL3) negatively regulates tumor necrosis factor α (TNFα) expression in association with p50 and also inhibits interleukin (IL)-1β independently of p50. 8 Another effector of endotoxin tolerance is IL-1 receptor-associated kinase 3, also known as IRAK-M, which is specifically expressed in monocytes and macrophages. 9 It has been demonstrated that endotoxin-tolerant (ET) macrophages share many features of alternatively activated M2 macrophages. 10,11 Furthermore, a common feature of endotoxin tolerance is the increase in nuclear p50 homodimers, which play an important role in M2 polarization. 11 Importantly, recent studies have suggested that energy metabolism in macrophages influences their phenotypic responses to stimuli. 12,13 While proinflammatory M1 macrophages largely depend on glycolysis, anti-inflammatory M2 macrophages prefer mitochondrial oxidative phosphorylation (OXPHOS) for the generation of energy. 14,15 Interestingly, inhibition of oxidative metabolism in macrophages not only blocks M2 phenotype but also drives macrophages toward an M1 phenotype. 13 Conversely, induction of OXPHOS in M1 macrophages can switch its phenotype to an M2 phenotype. 16 Therefore, macrophage phenotypes and their metabolism are functionally connected. However, it is not known if ET macrophages have an altered energy phenotype and whether energy phenotype contributes to the induction of endotoxin tolerance. Furthermore, although studies have supported that endotoxin tolerance plays a major role in the inhibition of inflammation, whether natural products and dietary bioactive components with known anti-inflammatory effects use this mode of action to exert their anti-inflammatory properties has not been investigated.
Spirulina platensis (SP), formally known as Arthrospira platensis, is a type of cyanobacterium or blue-green alga that is widely used for dietary supplements, animal feed, and pharmaceuticals. 17 The protective effects of SP against inflammatory diseases, such as colitis, arthritis, and allergic rhinitis, have been documented in several animal and human studies, 18 –20 although underlying molecular mechanisms are not well understood. We previously reported that SP and its organic extracts possess anti-inflammatory effects in vitro and in vivo. 21,22 In particular, pretreatment of RAW 264.7 and bone marrow-derived macrophages (BMDM) with SP organic extract (SPE) reduced LPS-induced cytokine secretion and mRNA expression of TNFα, IL-1β, and IL-6. In the present study, we sought to determine if endotoxin tolerance plays a role in the anti-inflammatory effect of SPE using a diet-induced obesity (DIO) mouse model, which manifests chronic inflammation, as well as in vitro macrophage models. Also, as macrophage energy metabolism is closely tied to the macrophage phenotype, in the present study, we investigated the role of SPE in the modulation of energy metabolism in comparison with that of ET macrophages using an XFe24 Analyzer.
Materials and Methods
Animal care and diet
Twenty-six male C57BL/6J mice (Jackson Laboratory, Bar Harbor, ME, USA) at 10 weeks of age were fed a modified AIN-93 high-fat/high-sucrose (HF/HS) diet (56.8% fat, 28.4% sucrose, w/w) for 4 weeks and then randomly assigned into a control (n = 13) or SPE group (n = 13). The control mice continued on the HF/HS diet, while the SPE group was fed the HF/HS diet supplemented with 0.25% SPE (w/w). SPE used in the diet was extracted as previously described. 21 Mice were housed in a controlled environment with a 12-h light/12-h dark cycle and were fed ad libitum. Body weights and food consumption were recorded weekly, and blood samples were drawn biweekly from the lateral tail vein. After 16 weeks on the experimental diets, mice were fasted for 8 h and anesthetized by ketamine/xylazine (100/10 mpk) (Henry Schein Animal Health, Dublin, OH, USA). Terminal blood samples were collected by cardiac puncture and allowed to clot for 30 min at room temperature before centrifugation at 1500 g for 10 min at 4°C to collect serum. Epididymal adipose tissues were snap frozen in liquid nitrogen and stored at −80°C. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Connecticut.
Serum chemistry
Serum concentrations of total cholesterol (TC) and total triglyceride (TG) were determined by enzymatic analysis using a cholesterol reagent from Pointe Scientific (Canton, MI, USA) and an L-Type TG M kit from Wako Chemical USA (Richmond, VA, USA), respectively, as we previously described. 23 Serum alanine transaminase (ALT) and glucose levels were determined using a Liquid ALT (SGPT) reagent set and a Liquid Glucose (Oxidase) reagent set from Pointe Scientific according to the manufacturer's protocol. Serum cytokines were measured by Luminex/xMAP magnetic bead-based multiplexing assays using MAGPIX instrumentation (EMD Millipore, Billerica, MA, USA) with a Bio-Plex® cytokine assay (Bio-Rad, Hercules, CA, USA).
Splenocyte isolation and treatment
Splenocytes were isolated from a subset of mice fed an experimental diet for 16 weeks as previously described. 21 Isolated splenocytes were then plated at a density of 2 × 106 cells/mL and then stimulated with 500 ng/mL of LPS (Sigma-Aldrich, St. Louis, MO, USA) for 20 h. Subsequently, the cells were harvested to measure inflammatory gene expression.
Flow cytometric analysis
Splenocytes used for flow cytometry were isolated as described above with the following exception: instead of RBC lysis buffer, red blood cells were removed by Histopaque-1083 (Sigma-Aldrich) density gradient centrifugation following the manufacturer's protocol. The cells were then counted and 1 × 106 cells were incubated with the following monoclonal antibodies to label monocyte subset 24 : CD115-PE, CDllb-APC, Ly6C-FITC, and Ly6G-PECy7. After staining, cells were fixed in 1% paraformaldehyde for 10 min and subsequently stored in 0.1% paraformaldehyde until cell sorting the following day. Data from samples and florescence minus one control were acquired using a BD FACSCalibur flow cytometer (BD Biosciences, San Jose, CA, USA) at the Flow Cytometry Facility at the University of Connecticut. OneComp eBeads (eBioscience, San Diego, CA, USA) were used as compensation controls. Data analysis and postacquisition compensation were conducted using FlowJo software (Tree Star, Ashland, OR, USA).
Cell culture and treatments
RAW 264.7 macrophages (RAW macrophages) and THP-1 monocytes were purchased from ATCC (Manassas, VA, USA). The cells were maintained in RPMI-1640 containing 10% fetal bovine serum, 1 × vitamin mix, 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM
Reverse transcription and quantitative real-time polymerase chain reaction
Total RNA was extracted from epididymal adipose tissue and cells, using TRIzol® RNA Isolation Reagent (Thermo Fisher Scientific, Waltham, MA), and reverse transcribed as previously described. 26 Quantitative real-time polymerase chain reaction (qPCR) was conducted using the SYBR green method with a CFX96 real-time PCR detection system (Bio-Rad). Data were analyzed by the 2−ΔΔCT method using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an internal control. Primer sequences were designed according to GenBank database using the Beacon Designer software (PREMIER Biosoft, Palo Alto, CA, USA). Primer sequences will be available on request.
Preparation of cell fractions and Western blot analysis
Cytoplasmic and nuclear fractions were prepared from cells using a Nuclear Extraction Kit (Cayman Chemicals, Ann Arbor, MI, USA). Western blot analysis was performed as previously described. 27 The following antibodies were used: TATA binding protein (TBP) and p105/p50 (Cell Signaling Technologies, Danvers, MA, USA); and BCL3, p65, and GAPDH (Santa Cruz Biotechnology, Dallas, TX, USA). The blots were developed using an Immun-Star™ WesternC™ Chemiluminescence reagent (Bio-Rad) and images were captured with a ChemiDoc XRS+ (Bio-Rad) and Image Lab software (Bio-Rad). GAPDH and TBP were used for purity of cytoplasmic and nuclear fractions, respectively, as well as loading controls.
Determination of cell energy phenotype
RAW macrophages were seeded at a concentration of 2.5 × 104 cells/well in an XFe24 cell culture plate and cultured overnight. The cells were then treated with SPE (25 μg/mL) or tolerized with LPS (10 ng/mL) for 12 h, after which they were subjected to a Seahorse Mito Stress Test to measure oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) using an XFe24 Analyzer (Seahorse Bioscience, North Billerica, MA, USA). Another set of cells was pretreated with SPE or tolerized with LPS for 12 h, and subsequently, they were activated with high dose of LPS (100 ng/mL) for an additional 12 h. Following treatments, cells were washed twice with XF media (nonbuffered RPMI-1640 containing 25 mM glucose, 2 mM
Statistical analysis
One-way analysis of variance and Tukey's post hoc analysis or unpaired t-test were performed with GraphPad Prism 6.0 (GraphPad Software, La Jolla, CA, USA) to detect differences between group means. P values less than .05 were considered significant. Values are expressed as mean ± SEM.
Results
Effect of SPE supplementation on phenotypic characteristics of DIO mice
There were no significant differences in body weights between control and SPE-fed mice throughout 16 weeks on the experimental diets (Fig. 1A). Both groups of mice consumed similar amount of diet during the study (data not shown). Serum levels of ALT, glucose, TC, TG, IL-1β, and monocyte chemoattractant protein-1 (MCP-1) were not significantly different between control and SPE-fed mice (Fig. 1B, C). The expression of inflammatory genes, including TNFα, IL-1β, IL-6, and MCP-1, as well as macrophage markers, that is, F40/80 and CD11c, was not significantly different between control and SPE-fed mice (Fig. 1D).

Body weight and metabolic phenotype of DIO mice fed a HF/HS diet supplemented with or without 0.25% SPE (w/w) through 16 weeks.
Effect of SPE on LPS reactivity and monocyte population of splenocytes
Splenocytes isolated from mice on an experimental diet for 16 weeks were challenged with LPS ex vivo. When the splenocytes were not stimulated with LPS, there were no significant differences in the expression of TNFα, IL-6, and IL-1β between control and SPE splenocytes (Fig. 2A). However, LPS challenge significantly increased mRNA levels of the proinflammatory cytokines in control splenocytes, while there was no induction of IL-6 and IL-1β in the splenocytes from SPE-fed mice.
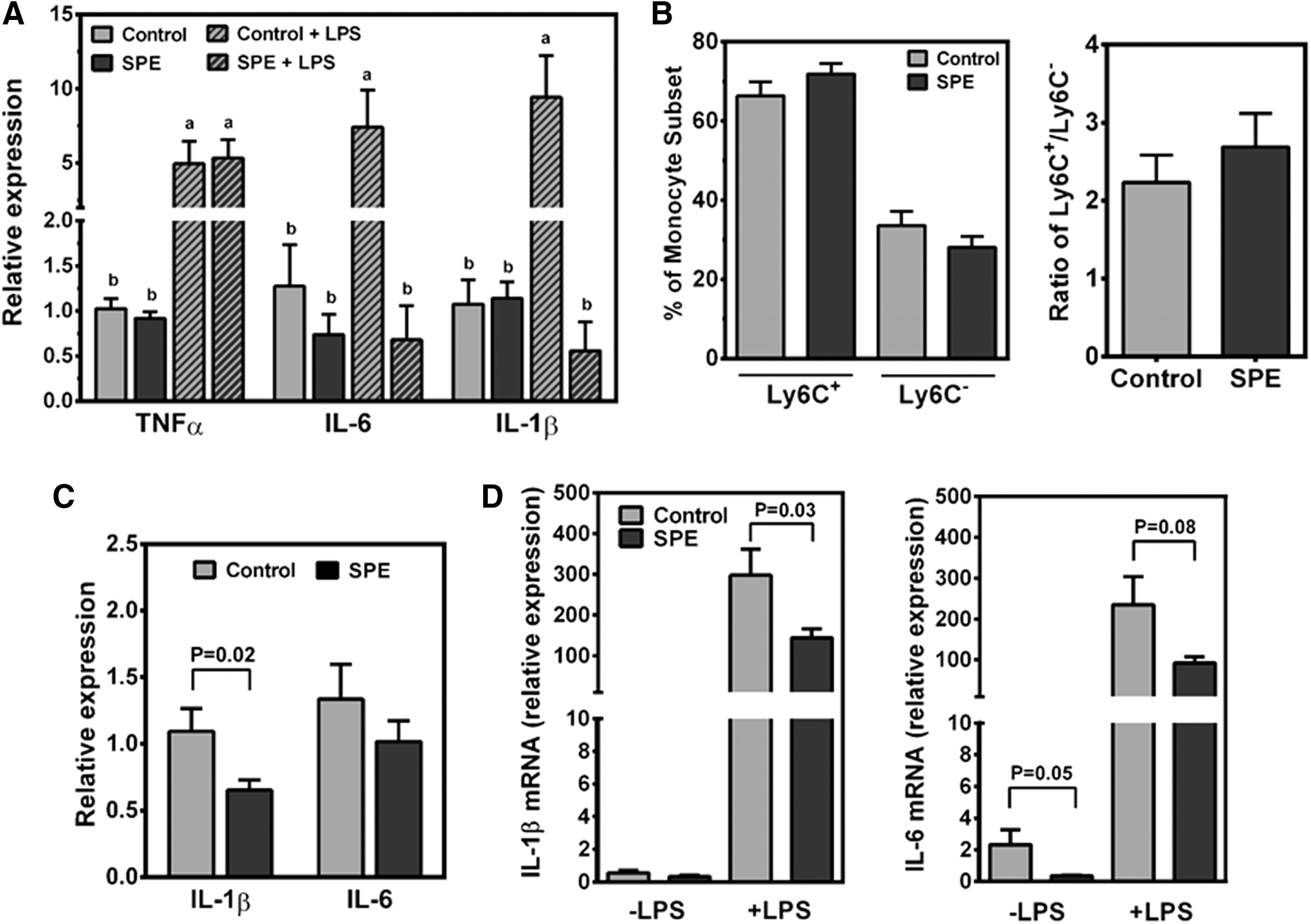
Ex vivo evaluations of splenocytes, peritoneal macrophages, and serum from DIO mice fed a HF/HS diet supplemented with or without 0.25% SPE (w/w) through 16 weeks.
As the spleen is a major reservoir for monocytes in the body, 29 it is conceivable that the decrease in LPS-induced proinflammatory gene expression in the splenocytes of SPE-fed mice may be due to changes in monocyte numbers and/or populations. To test this possibility, we conducted flow cytometric analysis to determine monocyte number and subsets within the spleen. Ly6C+ monocytes were present in greater numbers than Ly6C− monocytes, making up ∼55–80% of the monocytes in the spleen (Fig. 2B). However, there were no significant changes in the percentages of Ly6C+ or Ly6C− monocytes in the spleen of SPE-fed mice compared to those of control mice, nor was there a significant change in the ratios of Ly6C+ to Ly6C− monocytes.
Inflammatory responses of mouse serum-treated BMDM and LPS stimulation of peritoneal macrophages from mice fed experimental diets
To determine whether factors present in mouse circulation may affect inflammation, we incubated BMDM with 10% serum from SPE-fed or control mice. The expression of IL-1β was significantly lower in cells treated with SPE-fed serum than those treated with control serum (Fig. 2C). After 16 weeks of experimental diets, resident peritoneal macrophages were isolated and stimulated with LPS ex vivo. In unstimulated peritoneal macrophages, the expression of IL-1β was not significantly different between groups, but IL-6 mRNA levels were significantly lower in the SPE group (Fig. 2D). When peritoneal macrophages were stimulated with LPS, both IL-1β mRNA abundance was significantly less and IL-6 mRNA showed a trend toward decrease in the peritoneal macrophages isolated from SPE-fed mice compared to those from control mice.
Induction of an endotoxin tolerance-like state by SPE in macrophages
Endotoxin tolerance is a state of hyporesponsiveness to LPS stimulation in macrophages due to prior exposure of LPS. 5 Interestingly, as SP is a cyanobacterium, it has been suggested to contain LPS-like compounds. 30 Therefore, we determined the possibility that macrophages exposed to SPE may have features similar to endotoxin tolerance. RAW macrophages were first treated with 10–50 μg/mL of SPE or 5–50 ng/mL of LPS for 12 h to generate ET macrophages. The expression of IL-1β increased in response to all concentrations of SPE and LPS, although LPS induced a much higher expression of IL-1β than SPE (Fig. 3A). After pretreatment with SPE and low-dose LPS (i.e., endotoxin tolerization), the cells were activated with high level of LPS (100 ng/mL) for additional 12 h. Prior exposure to SPE and a low dose of LPS rendered cells to be less sensitive to high-dose LPS with marked decreases in IL-1β mRNA. To exclude the possibility that the endotoxin tolerance-like mechanism by SPE observed in RAW macrophages was cell-line specific, we treated RAW macrophages, human THP-1 macrophages, and BMDM with 25 μg/mL or 10 ng/mL of LPS for 12 h and then stimulated them with 100 ng/mL of LPS for additional 12 h. IL-1β mRNA levels in response to LPS (100 ng/mL) were significantly attenuated in all three macrophages pretreated with 25 μg/mL SPE or tolerized with 10 ng/mL LPS compared to the naive cells that had no prior exposure to SPE or LPS (Fig. 3B). The expression of IL-6, TNFα, and MCP-1 was also repressed in ET- as well as SPE-pretreated RAW macrophages (Fig. 3C).
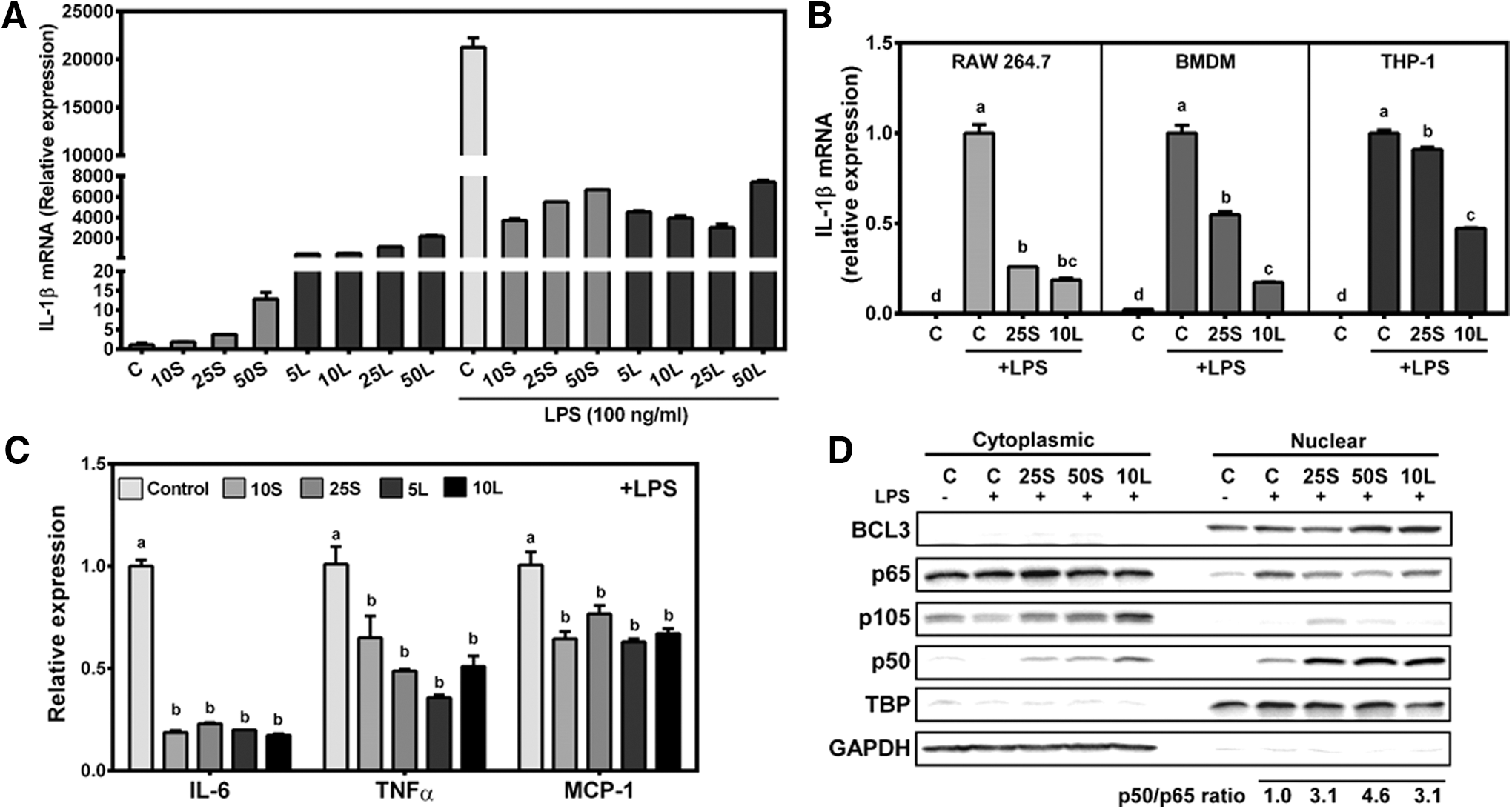
Inhibition of LPS-stimulated inflammatory gene expression by SPE similarly as ET macrophages.
As p50 homodimer translocation into the nucleus is indicative of endotoxin tolerance, 7 we determined nuclear translocation of p50. Both SPE-pretreated and ET RAW macrophages displayed a marked increase in nuclear p50 protein with concomitant decreases in the nuclear presence of p65 on LPS challenge (Fig. 3D). In contrast, on stimulation with 100 ng/mL LPS, control cells had marked increase in p65, but lower p50 levels in the nucleus than ET- and SPE-treated cells. Quantification of nuclear p50 and p65 proteins showed that in SPE-pretreated and ET RAW macrophages, nuclear p50/p65 ratios increased more than 3-fold compared to control cells in response to LPS stimulation. We also determine the level of nuclear BCL3 that can inhibit proinflammatory mediator expression in association with p50 homodimers in endotoxin tolerance. 7,8 Nuclear BCL3 protein levels were noticeably changed in ET macrophages and macrophages treated with 50 μg/mL of SPE.
Increases in negative regulators of TLR4 signaling by SPE in macrophages
To determine why macrophage stimulation with a high dose of LPS was attenuated in SPE-pretreated and ET macrophages, we investigated changes in negative regulators of TLR4 signaling in SPE-treated and ET macrophages before high-dose LPS stimulation. Treatment of RAW macrophages with 25 μg/mL of SPE, but not control or ET macrophages, increased BCL3 protein in the nucleus (Fig. 4A). Both SPE and endotoxin tolerization increased nuclear p50 protein, while control cells had no detectable p50 in the nucleus. Consistently, both SPE and LPS significantly increased the expression of BCL3 and p105/p50 (Fig. 4B). Interestingly, only LPS significantly increased IκBα and IRAK-M mRNA levels.
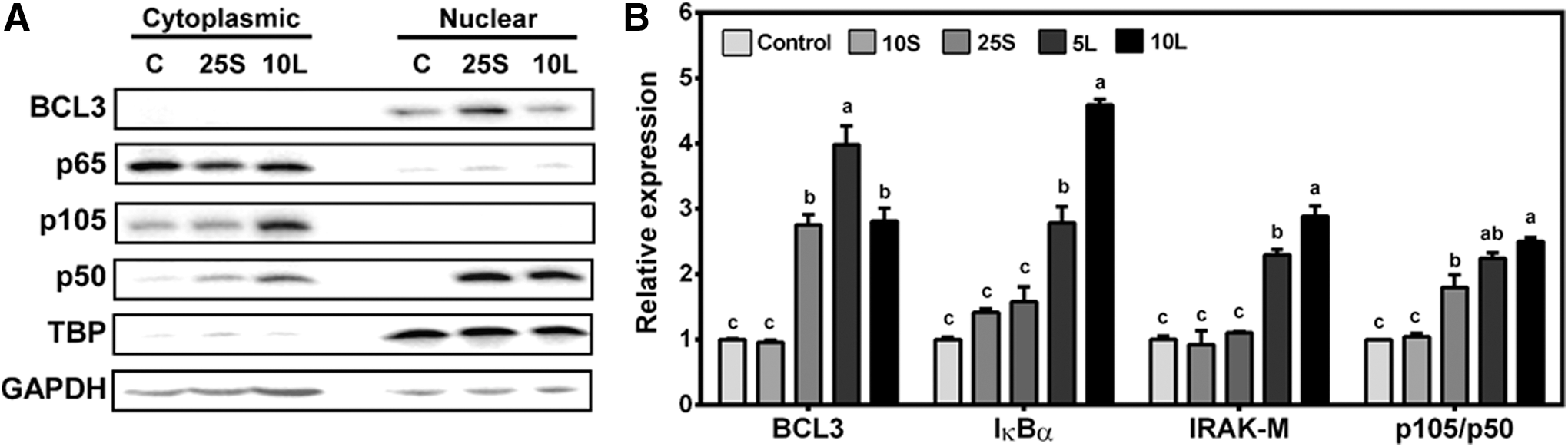
Expression of endotoxin tolerance-associated genes and proteins in response to SPE treatment and endotoxin tolerization.
ET- and SPE-treated macrophages have a similar energy phenotype
As energy metabolism in macrophages can have an impact on their inflammatory phenotypes, 13,14 we investigated changes in glycolytic and oxidative capacities of macrophages exposed to SPE and endotoxin tolerization. ET macrophages had a significant reduction in basal OCR compared to controls, while the basal OCR of macrophages treated with SPE was intermediate (Fig. 5A, B). The basal ECAR was significantly increased in SPE-treated macrophages compared to controls, while ET macrophages had an intermediate ECAR (Fig. 5C). The OCR/ECAR ratio indicates cell's preference for glycolysis or OXPHOS. 28 Both SPE-treated and ET macrophages had significantly lower OCR/ECAR ratios than controls, indicating their preference for glycolysis as an energy-generating pathway to OXPHOS (Fig. 5D). In response to tolerization with LPS, but not SPE treatment, ATP production was significantly attenuated (Fig. 5E). Interestingly, the spare respiratory capacity (SRC), an indicator of how well cells can respond to immediate energy demands, 31 was significantly elevated in macrophages treated with SPE (Fig. 5F).
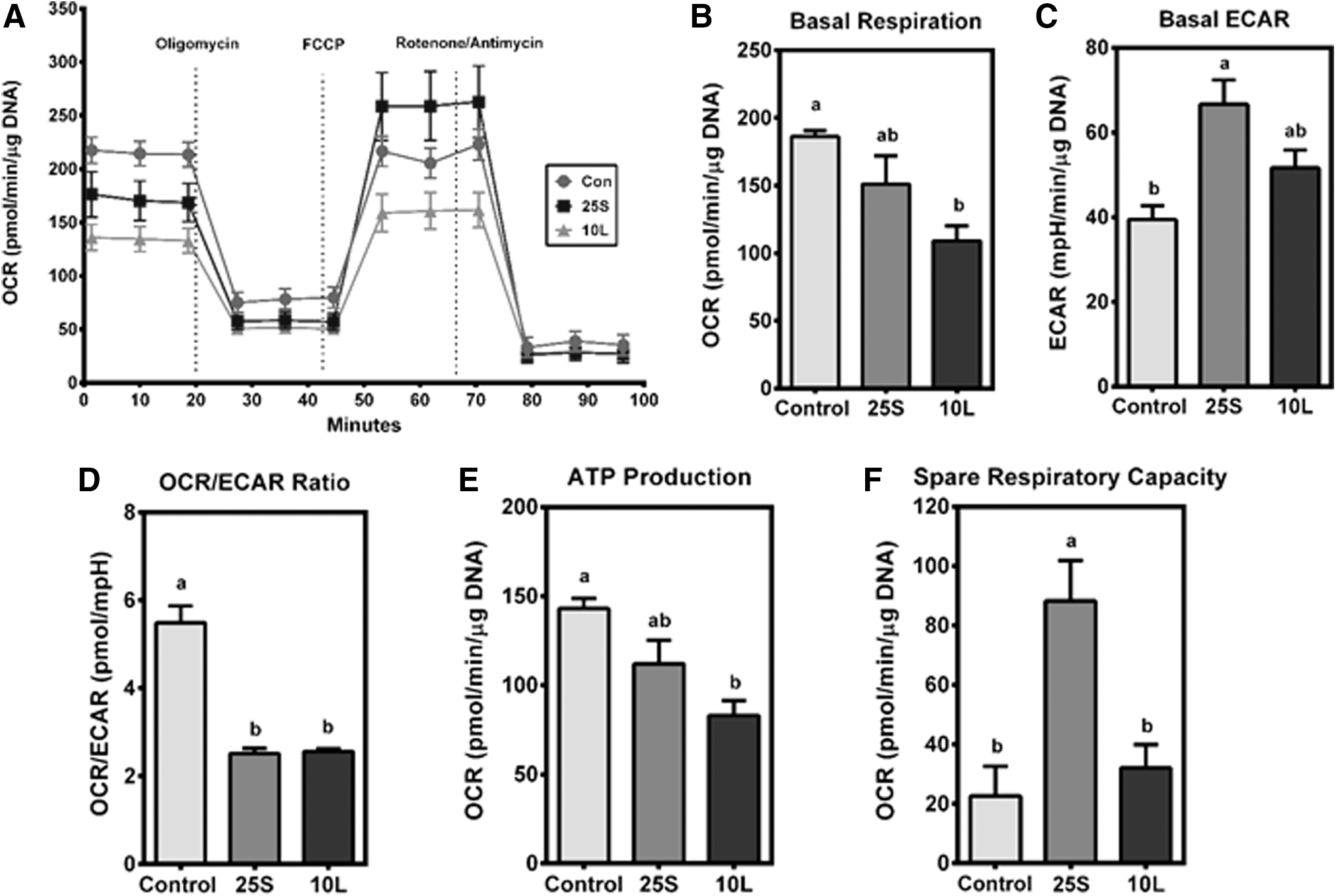
Energy phenotype of ET- and SPE-treated RAW macrophages.
Discussion
SP has been demonstrated to prevent inflammatory diseases such as colitis, arthritis, and allergic rhinitis. 18 –20 We also have shown that SPE reduces LPS-induced cytokine secretion and mRNA expression of TNFα, IL-1β, and IL-6 in RAW macrophages and BMDM. 21 As SP is a type of cyanobacteria that may possess components similar to LPS capable of stimulating immune responses 30 such as endotoxin tolerance, we rationalized that SPE may induce an endotoxin tolerance-like state in macrophages to exert its anti-inflammatory effect. It is well documented that DIO in mice leads to infiltration of immune cells into metabolic tissues, which ultimately increases cytokine production and inflammation. 32 Therefore, in the present study, we sought to determine if SPE supplementation may attenuate DIO-induced inflammation through the induction of endotoxin tolerance. We found that SPE did not alter body weights and other metabolic phenotypes of DIO mice. However, splenocytes and peritoneal macrophages isolated from SPE-fed mice showed significantly less sensitivity to LPS ex vivo than those from control mice. Our observations suggest that SPE specifically alters immune/inflammatory programs independent of metabolic changes in DIO mice. The ex vivo findings were supported by our observations that ET macrophages showed significantly less proinflammatory gene expression, which was also observed in SPE-pretreated macrophages on high-dose LPS challenge. Furthermore, SPE increased mRNA levels of endotoxin tolerance-associated genes as well as the abundance of nuclear p50 and BCL3 similarly to ET macrophages. Our data suggest that SPE may exert its anti-inflammatory effect through the induction of an endotoxin tolerance-like state in macrophages.
The spleen is a major reservoir of monocytes. 29 The Ly6C+ monocyte subset is more likely to become inflammatory M1 macrophages with high antimicrobial activity due to their potent capacity for phagocytosis and cytokine secretion, while the Ly6C− monocytes secrete IL-10, a major anti-inflammatory cytokine, and tend to differentiate into anti-inflammatory M2 macrophages. 33 In the present study, as we found reduced inflammatory gene expression in the splenocytes from SPE-fed mice compared with control splenocytes, we speculated that changes in splenic monocyte subsets may be responsible for the reduction. However, our flow cytometric analysis for the determination of splenic monocyte subsets demonstrated that the number of the Ly6C+ and Ly6C− monocytes in the spleen as well as the ratios of Ly6C+ to Ly6C− monocytes did not differ in control or SPE splenocytes. This suggests that the reduction in LPS-induced proinflammatory gene expression in the splenocytes from SPE-supplemented group is not likely due to altered monocyte subsets; rather it may be attributable to reduced reactivity of the splenocytes to LPS.
Endotoxin tolerance is a state of decreased LPS sensitivity due to prior LPS exposure in immune cells. 5,34 As a type of cyanobacterium, SP may possess pathogen-associated molecular patterns that can stimulate immune cells. 30 Because we observed reduced LPS sensitivity of the splenocytes and the peritoneal macrophages isolated from mice fed SPE, which is reminiscent of endotoxin tolerance, we explored the possibility that SPE may induce endotoxin tolerance in vitro by comparing the effects of LPS stimulation on proinflammatory gene expression in naive macrophages to those in macrophages pretreated with SPE or macrophages tolerized to LPS using three different macrophage types, that is, RAW macrophages, THP-1 macrophages, and mouse BMDM. When the macrophages were pretreated with SPE, a significant reduction in the expression of IL-1β was observed on LPS stimulation in all three macrophage types, demonstrating that the effect of SPE on inducing an endotoxin tolerance-like state is conserved between different types of macrophages.
In vitro endotoxin tolerization of macrophages involves two distinct phases. 34 The first phase is the exposure of macrophages to LPS for the first time, typically with a low LPS dose. The second phase is the stimulation of cells with re-exposure of LPS, typically at a higher dose than the first LPS exposure. We investigated the mechanisms of action by which macrophages pretreated with SPE induce an endotoxin tolerance-like state similarly to ET cells during the first phase. NF-κB p50 is generated by the processing of a precursor molecule p105. 6 p50 does not contain a transactivation domain, and therefore, DNA-bound p50 homodimers have been found to repress NF-κB-dependent proinflammatory gene expression. 6 Several studies have reported that nuclear levels of p50 homodimers become elevated in ET macrophages, supporting a central role of p50 homodimers in the suppression of NF-κB activation in endotoxin tolerance. 35 –37 BCL3 is a nuclear protein of the IκB family containing an ankyrin repeat motif, which mediates interaction with NF-κB dimers. 7 BCL3 has been demonstrated to associate with p50 and p52 homodimers, 38,39 limiting the expression of proinflammatory mediators in endotoxin tolerance. 8 Also, IRAK-M, specifically expressed in monocytes and macrophages, 9 prevents dissociation of IRAK and IRAK-4 from MyD88, consequently inhibiting TLR4 signaling. 40 Endotoxin tolerance is significantly attenuated in Irak-m knockout mice, indicating that IRAK-M mediates endotoxin tolerance. 40 Therefore, p50 homodimers, BCL3 and IRAK-M, have been shown to play important roles in the induction of endotoxin tolerance. In the present study, both SPE and endotoxin tolerization increased p50 accumulation in the nucleus, but nuclear levels of BCL3 were increased only by SPE. At the mRNA level, both SPE and low-dose LPS significantly increased the expression of BCL3 and p105, but only LPS induced IκBα and IRAK-M expression. These results suggest that SPE, like low-dose LPS, can increase effectors of endotoxin tolerance such as p50, but it may be also distinct from LPS as it did not induce all effectors of endotoxin tolerance that were induced by LPS.
When we investigated the effect of SPE on the second phase of endotoxin tolerance induction, we found that SPE-pretreated or ET RAW macrophages had increased p50 protein in the nucleus when stimulated by LPS. The increased nuclear p50 may not be indicative of endotoxin tolerance as p50 may form heterodimers with p65, which are transcriptionally active and proinflammatory. 6 However, ET and SPE-pretreated cells had more than a 3-fold increase in the p50/p65 ratios than those of naive macrophages on LPS stimulation. Therefore, increased p50/p50 homodimer formation is likely to occur in the nucleus of SPE-pretreated and ET macrophages, inhibiting the formation of proinflammatory p65/p50 heterodimers and/or competing with p65/p50 for binding to the promoters of proinflammatory genes. In the latter case, as p50/p50 homodimers do not have transcription activity, the expression of proinflammatory genes can be decreased. Importantly, our observations indicate that attenuation of LPS-induced inflammatory gene expression in response to LPS re-exposure may be attributable to the upregulation of negative regulators of TLR signaling, such as BCL3 and p50, by SPE.
Studies have shown that ET and M2 macrophages exhibit similar as well as distinct characteristics. For instance, ET macrophages have increased expression of macrophage receptor with collagenous structure and formyl-peptide receptor ligand 1, which are associated with wound healing in M2 macrophages, as well as the expression of matrix metalloproteases, genes that are associated with wound healing in M2 macrophages. 10,41 Furthermore, a common feature of endotoxin tolerance is the increase in nuclear p50 homodimers, which has been demonstrated to play a role in M2 polarization. 11 Interestingly, however, ET macrophages also have increased phagocytic ability coupled with increased reactive oxygen species (ROS) production, features of M1 macrophages. 5,42 It has not been appreciated thus far what type of energy phenotype ET macrophages have. This is a critical question to gain insight into mechanisms by which SPE-treated macrophages exert strong anti-inflammatory properties while they show an ET state. We found that during LPS tolerization and SPE treatment, there was a decrease in basal OCR but an increase in basal ECAR. As OCR is a measurement of OXPHOS and ECAR represents glycolysis, the data indicate that LPS tolerization and SPE may decrease OXPHOS while increasing glycolysis in macrophages. It has been suggested that similar changes in OXPHOS and glycolysis occurring in proinflammatory macrophages is to shift the use of mitochondria from ATP generation to ROS production for bactericidal activities while the macrophages rely on glycolysis for energy, although this way is not as efficient as OXPHOS. 43 Therefore, it is presumable that endotoxin tolerization and SPE treatment may facilitate ROS production to enhance bactericidal activity of the macrophages. Interestingly, although LPS tolerization and SPE treatment displayed similar levels of dependency on glycolysis for energy generation as evidenced by their comparable OCR/ECAR ratios, SRC values in SPE-treated cells were significantly higher than LPS-tolerized cells. The increased SRC with SPE treatment is likely independent of its ability to induce endotoxin tolerance because there was no difference in the SRC of ET macrophages and naive control macrophages.
It is apparent from the in vitro experiments that SPE induces an endotoxin tolerance-like state in macrophages. This function of SPE may explain our observation that the splenocytes and peritoneal macrophages isolated from SPE-fed mice displayed decreases in LPS-induced inflammatory gene expression. Interestingly, several studies have described that supplementation of SP or its extracts in several animal models and humans activates the immune system, characteristic of the first exposure in endotoxin tolerance. In broiler chickens fed SP, macrophage phagocytic function and nitrite production were enhanced. 44 In isolated chicken peritoneal macrophages, treatment of a water-soluble SP extract increased cell spread, vacuolization, and phagocytic activity. 45 It has been reported that Braun-type lipoprotein-enriched extracts of SP (Immunlina®) increased the mRNA expression of IL-1β and TNFα in human THP-1 monocytes. 46 Furthermore, Immunlina consumption increased adaptive immune responses by increasing CD4+ T helper cell and B cell proliferation in response to foreign antigen in healthy subjects. 47 Supplementation of healthy men with 400 mg/day of Immulina® increased the ability of natural killer cells to kill K562 tumor cells. 48 Coincidentally, ET monocyte and macrophages have been shown to have increased phagocytic function and antimicrobial activity due to elevated ROS production. 34 The similarities in the effects of SP extracts on immune cells and endotoxin tolerance suggest that the anti-inflammatory and immune-boosting effects of SP or its extract may be mediated to the induction of endotoxin tolerance.
In conclusion, we found that SPE induced an endotoxin tolerance-like state in macrophages ex vivo and in vitro, which is likely to contribute to its potent anti-inflammatory properties. The SPE-induced ET features were similar to those of endotoxin tolerization, in that it increased both the mRNA expression of p105 and BCL3 and the nuclear abundance of p50 and BCL3. Also, both SPE-treated and ET macrophages have a similar energy phenotype. To our knowledge, this study shows for the first time that through dietary means, endotoxin tolerance can be induced to exert anti-inflammatory effects. Our findings would have wide implications to human health as endotoxin tolerance may be used for certain healthy conditions to attenuate overt inflammation such as in the case of sepsis.
Footnotes
Acknowledgments
This work was supported by the U.S. Department of Agriculture Multi-State Hatch CONS 00916 to J.L. and by the U.S. Department of Agriculture AFRI NIFA Pre-Doctoral Fellowship (2014-01870) to T.X.P.
Author Disclosure Statement
No competing financial interests exist.