
Abstract
Sirtuin 3 (SIRT3) regulates mitochondrial antioxidant (AO) defense and improves mitochondrial disorders. Curcumin protects mitochondria; however, the mechanisms need investigation. We postulated that curcumin increases AO defense under hyperglycemic conditions in HepG2 cells through SIRT3-mediated mechanisms. Cell viability was determined in HepG2 cells cultured with 5 mM glucose, 19.9 mM mannitol, vehicle control, 10 mM glucose, and 30 mM glucose in the absence or presence of curcumin for 24 h. SIRT3, nuclear factor-kappa B (NF-κB), heat-shock protein 70 (Hsp70), and Lon protein expressions were determined using western blot. Transcript levels of SIRT3, peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC-1α), cAMP response element-binding protein (CREB), glutathione peroxidase 1 (GPx1), and superoxide dismutase 2 (SOD2) were measured by quantitative polymerase chain reaction. Cell viability, SIRT3 protein expression, transcript levels of SIRT3, PGC-1α, CREB, GPx1, and SOD2 and protein expressions of NF-κB, Lon, and Hsp70 were significantly increased in the curcumin-treated hyperglycemic groups. Since curcumin and SIRT3 both improve mitochondrial function and AO defense, SIRT3 may be involved in the protective effects of curcumin.
Introduction
T
Mitochondrial proteins are vulnerable to oxidative damage as they are closely situated to the electron transport chain (ETC). Oxidative damage can form adducts of proteins with carbohydrates and lipids and may create carbonyl groups and intramolecular cross-links. These may impair or inactivate enzymes and structural proteins which, if not removed, can cause significant cellular toxicity associated with diseases. 1,2
Research has placed more focus on evaluating dietary antioxidant (AO) supplementation that may improve DMII conditions. For example, purified anthocyanin supplementation was shown to reduce OS and attenuate insulin resistance and DMII in animal models. 3
Curcumin is a phytochemical compound extracted from the rhizome of Curcuma longa and possesses multiple biological properties, including AO, 4 antibacterial, 5 and anti-inflammatory properties. 6 It is a bifunctional AO that (1) reacts directly with reactive oxygen species (ROS) and (2) indirectly induces upregulation of cytoprotective and AO proteins through nuclear translocation of nuclear factor erythroid 2-related factor (Nrf2). 7
Mitochondria are protected by curcumin under oxidative conditions. 8 Curcumin treatment prevented OS and mitochondrial dysfunction in potassium dichromate-induced nephrotoxicity. 9 It also decreased malondialehyde and protein carbonyl levels and prevented the decrease in the activity of hepatic AO enzymes viz. superoxide dismutase 2 (SOD2) and glutathione peroxidase 1 (GPx1). 10
Curcumin preserved mitochondrial parameters such as mitochondrial membrane potential and ATP production following toxicity. 8 It also indirectly increased the expression of mitochondrial biogenesis genes and attenuated OS by Nrf2.
Sirtuin 3 (SIRT3), a mitochondrial stress-responsive protein deacetylase, regulates mitochondrial processes such as the tricarboxylic acid cycle and oxidative phosphorylation (OXPHOS). 11 It reduces ROS levels by modulating AO enzymes, such as SOD2, thereby protecting against oxidative damage. 12
SIRT3 also regulates AO enzymes through transcriptional coactivator peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC-1α). The transcription factor nuclear factor-kappa B (NF-κB) is also deacetylated and activated by SIRT3. Activation of NF-κB increases the expression of SOD2, thus making cells resistant to OS-mediated cell damage. 13
SIRT3 deacetylates and activates Lon, a major mitochondrial protease, which degrades oxidized and damaged proteins and is involved in mtDNA maintenance and replication. 14 Heat-shock protein 70 (Hsp70) forms part of this protein quality control network as it has the ability to prevent protein aggregation and other protein modifications. 15
Since curcumin and SIRT3 both improve mitochondrial function and AO defense, SIRT3 may be involved in the protective effects of curcumin, although this has yet to be fully elucidated.
Molecular events that occur under hyperglycemic conditions in the liver are not fully known. This is important to interrogate as the liver is the primary organ involved in glucose homeostasis. In this study, we investigated the effect of curcumin on SIRT3 expression, downstream AO defense, and protein quality control under hyperglycemic conditions in a human hepatoma cell line (HepG2).
Materials and Methods
Cell culture and treatments
HepG2 cells were cultured to confluency in 25-cm3 flasks (5% CO2, 37°C) in complete culture media (Eagles minimum essential medium, supplemented with 10% fetal calf serum, 1% L-glutamine, and 1% penstrepfungizone).
The HepG2 cells (2.5 × 104 cells/well) were cultured in the presence of control (5.5 mM glucose), osmotic control (19.9 mM mannitol), vehicle control (0.1% DMSO), and hyperglycemic (10 mM [intermediate], 30 mM glucose) conditions in the absence or presence of curcumin (5 and 10 μM) for 24 h in 30 mm3 cell culture plates.
Cell viability assay
Following treatment, the cells (three replicates) were incubated with methyl thiazol tetrazolium (MTT) salt solution (5 mg/mL in 0.1 M phosphate-buffered saline) and complete culture media (4 h, 37°C). Following incubation, the supernatants were aspirated and DMSO was added (100 μL/well) and incubated at 37°C for a further 1 h. Optical density of the formazan product was measured by a microplate reader (Bio-Tek μQuant) at 570 nm with a reference wavelength of 690 nm.
Quantitative polymerase chain reaction
Total RNA was extracted from cultured cells using the TRIzol reagent (Ambion). cDNA was synthesized by reverse transcription using the iScript cDNA Synthesis Kit (Bio-Rad).
Quantitative polymerase chain reaction (qPCR) was performed on the CFX96 Real-Time System (Bio-Rad) by using iQ SYBR Green Supermix (Bio-Rad) with primer sequences as listed in Table 1. The PCR was initiated with the following thermocycler profile: an initial denaturation for 8 min at 95°C followed by 39 cycles of 95°C denaturation for 15 sec, annealing for 1 min and extension of 72°C for 30 sec. A final extension at 60°C was performed for 31 sec. Each measurement was done in triplicate and normalized against 18S rRNA Ct values. The qPCR data are represented as a relative fold change, calculated using the method described by Livak and Schmittgen. 16
CREB, cAMP response element-binding protein; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1 alpha; GPx1, glutathione peroxidase 1; SIRT3, sirtuin 3; SOD2, superoxide dismutase 2.
Western blot analysis
Proteins were isolated using a Cytobuster™ (Novagen) supplemented with protease and phosphatase inhibitors (Roche). Proteins were quantified using the bicinchoninic acid assay (Sigma) and standardized to 1 mg/mL. Protein extracts were prepared in Laemmli buffer (dH2O, 0.5 M Tris-Cl [pH 6.8], glycerol, 10% sodium dodecyl sulfate [SDS], β-mercaptoethanol, 1% bromophenol blue) and separated by SDS-polyacrylamide gel electrophoresis (7.5% resolving gel) and electrotransferred to nitrocellulose membrane. Membranes were blocked with 5% bovine serum albumin (BSA) in Tris-buffered saline and Tween 20 (TTBS; 137 mM NaCl, 2.7 mM KCl, 24 mM Tris, 0.5% Tween 20). Membranes were probed using the following antibodies in 5% BSA (in TTBS): rabbit anti-SIRT3 (Abcam, ab86671; 1:1000), rabbit anti-LONP1 (Sigma; HPA002192), rabbit anti-NF-κB p65 (CST No. 3033), and mouse anti-HSP70 (CST No. 4872). All membranes were incubated in primary antibodies overnight at 4°C. A horseradish peroxidase-conjugated goat polyclonal antibody specific for β-actin was used for the loading control and the normalization process (1:5000 in 5% BSA in TTBS). Membranes were developed using LumiGlo® Chemiluminescent Substrate System (KPL) and images were captured on the Alliance 2.7 documentation system (UViTech). The density of the bands was quantified using UViBand Advanced Image Analysis Software (UViTech). The experiment was performed in triplicate and repeated thrice. Results are presented as relative band intensity (RBD) and normalized against β-actin.
Statistical analysis
Each experiment was performed in triplicate and repeated thrice. Results are expressed as mean ± standard error of the mean (SEM). Statistical analysis was performed using one way ANOVA followed by Bonferroni test for multiple group comparison. Differences with P < .05 were considered statistically significant.
Results
Curcumin treatment increases mitochondrial output under normo- and hyperglycemic conditions
To determine the effect of curcumin on mitochondrial output, cell viability was measured in HepG2 cells.
Significantly higher cell viability was observed in the curcumin-treated control relative to the untreated control (5 μM [P = .0086] and 10 μM [P = .0014]). Curcumin-treated hyperglycemic groups also showed significantly higher cell viability relative to the untreated control (5 μM [10 mM: P = .0112; 30 mM: P = .0092] and 10 μM [10 mM: 0.7703 ±0.0095 optical density (O.D.), P value: .0107 and 30 mM 0.7207 ± 0.0006 O.D., P value: .0069]; Fig. 1).
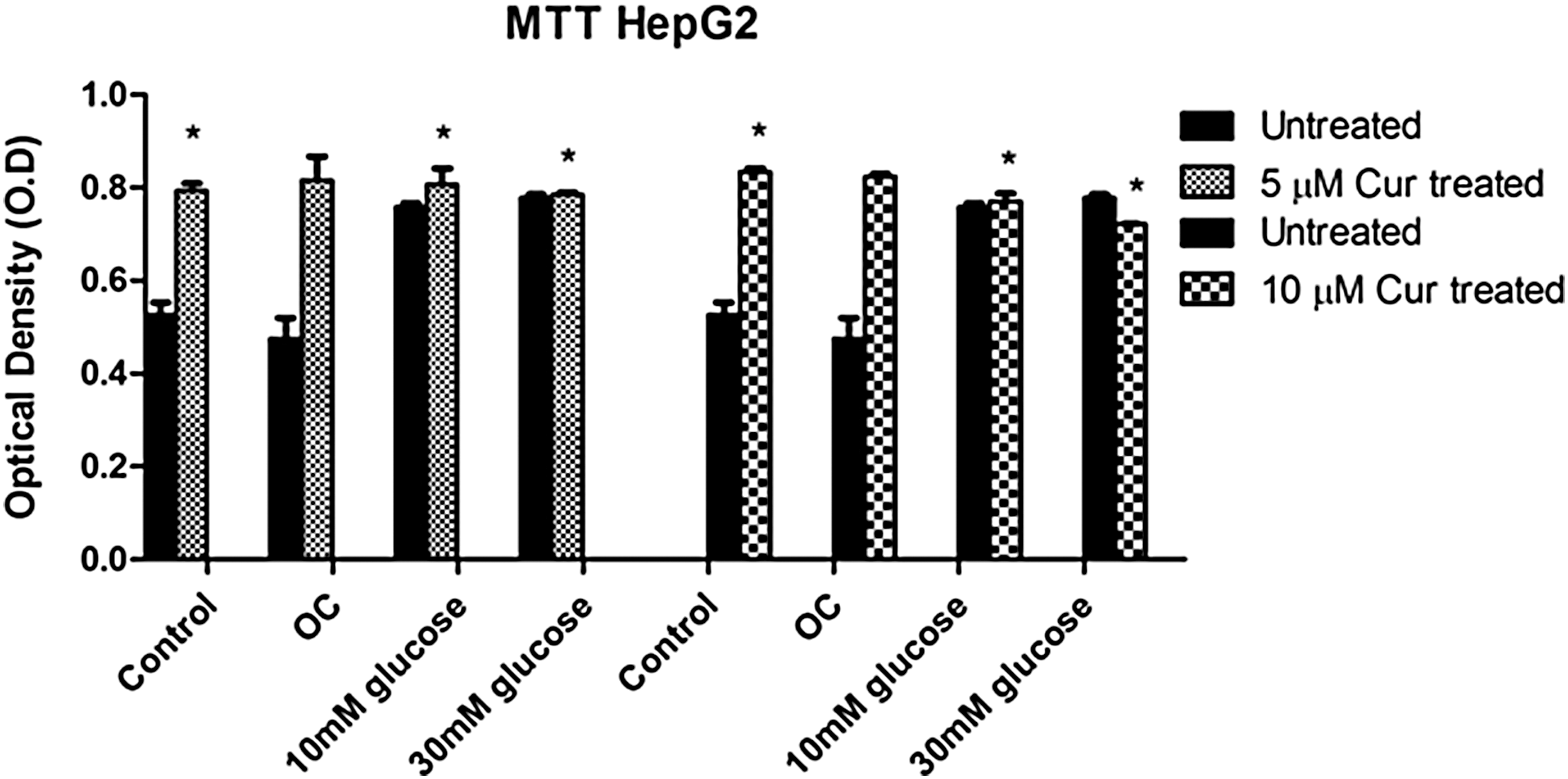
The effect of curcumin on cell viability was determined in HepG2 cells (untreated and curcumin-treated). Values are expressed as mean ± SEM. *P < .05 versus untreated control. SEM, standard error of the mean.
Curcumin increases SIRT3 expression under normo- and hyperglycemic conditions
To determine whether curcumin treatment had an effect on SIRT3 expression, western blot and qPCR were used to measure protein expression and mRNA levels of SIRT3, respectively, in HepG2 cells.
Significantly increased SIRT3 protein expression was observed in the curcumin-treated control and curcumin-treated hyperglycemic groups relative to the untreated control (5 μM [control: P = .0490; 10 mM: P = .0010] and 10 μM [control: P = .0403; 10 mM: 6.771 ± 0.5016 RBD, P = .0218; 30 mM: 5.991 ± 0.4698 RBD, P = .0293]; Fig. 2A) The treated hyperglycemic groups also showed statistically higher SIRT3 protein expression when compared with the treated control (10 mM: P = .0125; 30 mM: P = .0229; Fig. 2A).
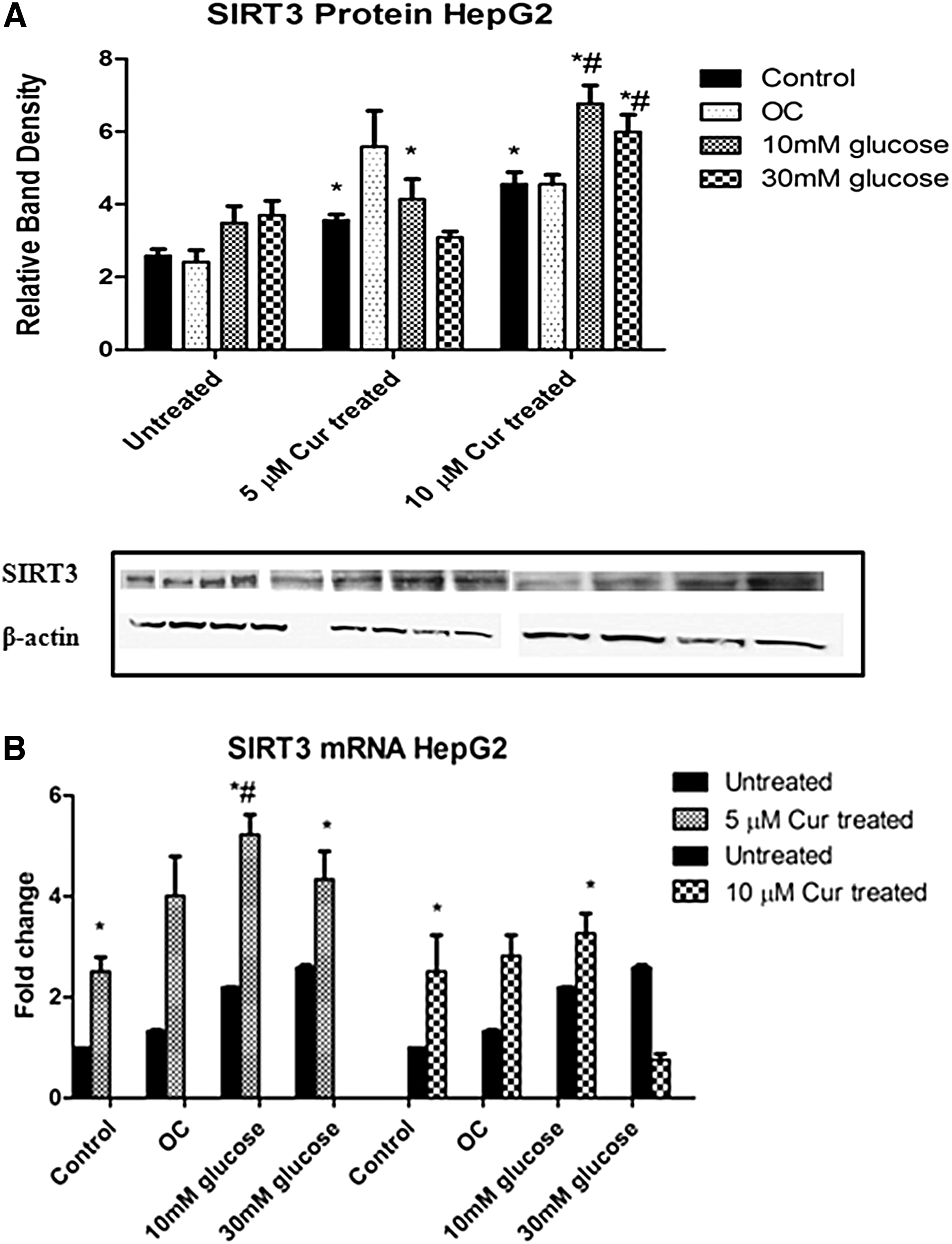
The effect of curcumin on SIRT3 expression in untreated and curcumin-treated HepG2 cells.
qPCR data showed increased SIRT3 levels in the curcumin-treated control and curcumin-treated hyperglycemic groups relative to the untreated control (5 μM [control: P = .0114, 2.5-fold; 10 mM: P = .0031, 5.1-fold; 30 mM: P = .0100, 4.2-fold] and 10 μM [control: P = .0097, 2.8-fold; 10 mM: P = .0110, 3.1-fold]; Fig. 2B).
Curcumin increases CREB expression under normo- and hyperglycemic conditions
To investigate the effect of hyperglycemia and curcumin treatment on cAMP response element-binding protein (CREB) expression, qPCR was used to measure mRNA levels of CREB in HepG2 cells
Significantly higher CREB levels were observed in the curcumin-treated control and curcumin-treated hyperglycemic groups, relative to the untreated control (5 μM [control: P = .0083; 10 mM: P = .0032; 30 mM: P = .0209] and 10 μM [10 mM: 5.9-fold, P = .0080; 3.9-fold, 30 mM: P = .0049; Fig. 3]). The intermediate hyperglycemic group showed relatively higher CREB expression when compared with the treated control, but this did not reach statistical significance.
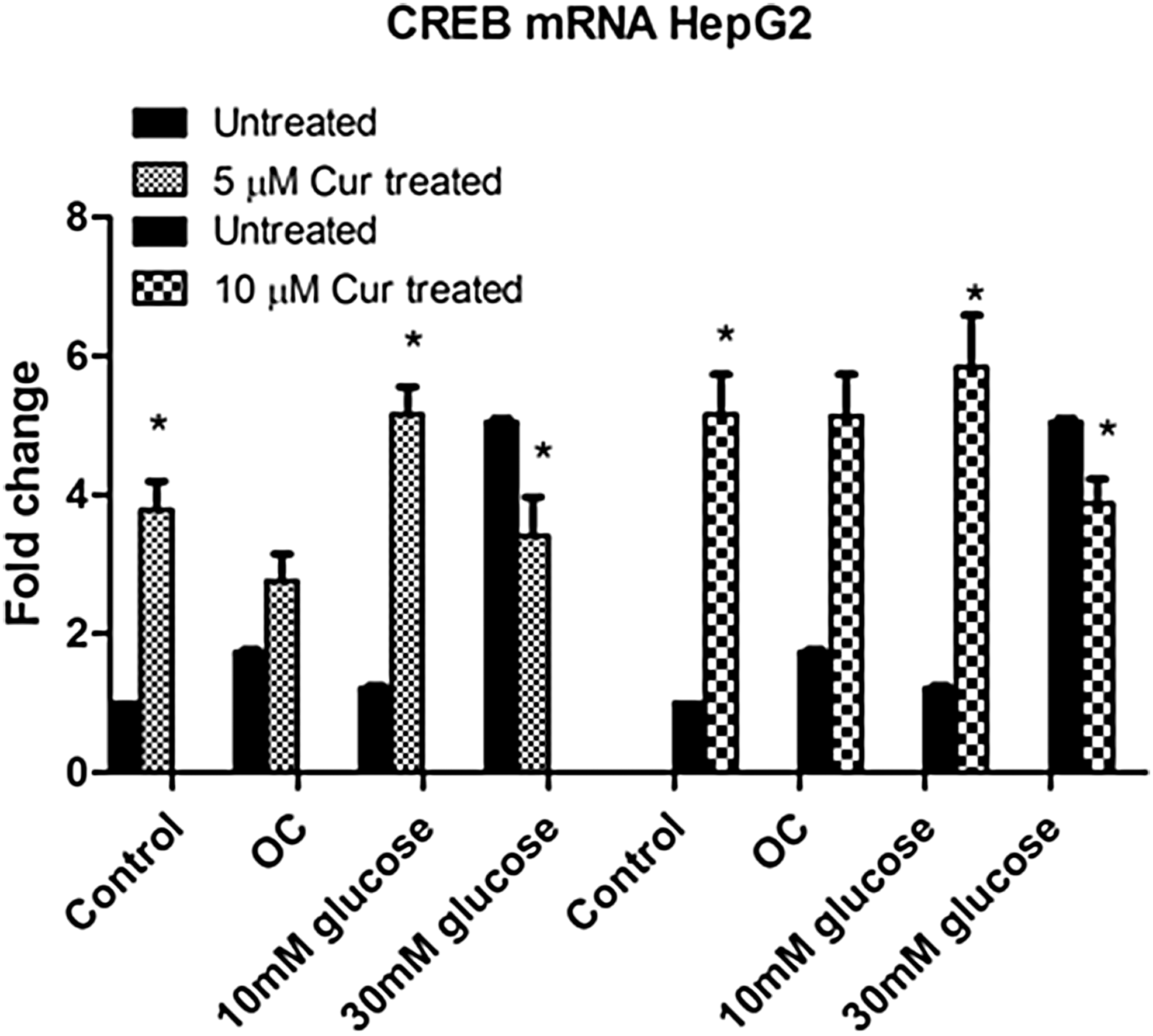
The effect of curcumin on CREB expression in untreated and curcumin-treated HepG2 cells. Gene expression for CREB was assessed. Values are expressed as fold changes relative to the control. Each bar represents the mean ± SEM of three replicates. *P < .05 versus untreated control, P < .05 versus curcumin-treated control. CREB, cAMP response element-binding protein.
Curcumin increases PGC-1α expression under normo- and hyperglycemic conditions
The effect of curcumin treatment on PGC-1α gene expression was investigated in HepG2 cells using qPCR.
Higher PGC-1α levels were observed in the curcumin-treated hyperglycemic groups relative to the untreated control (5 μM [control: P = .0033; 10 mM: P = .0128; 30 mM: P = .0164] and 10 μM [control: P = .0080; 10 mM: P = .0168, 5.1-fold]; Fig. 4).
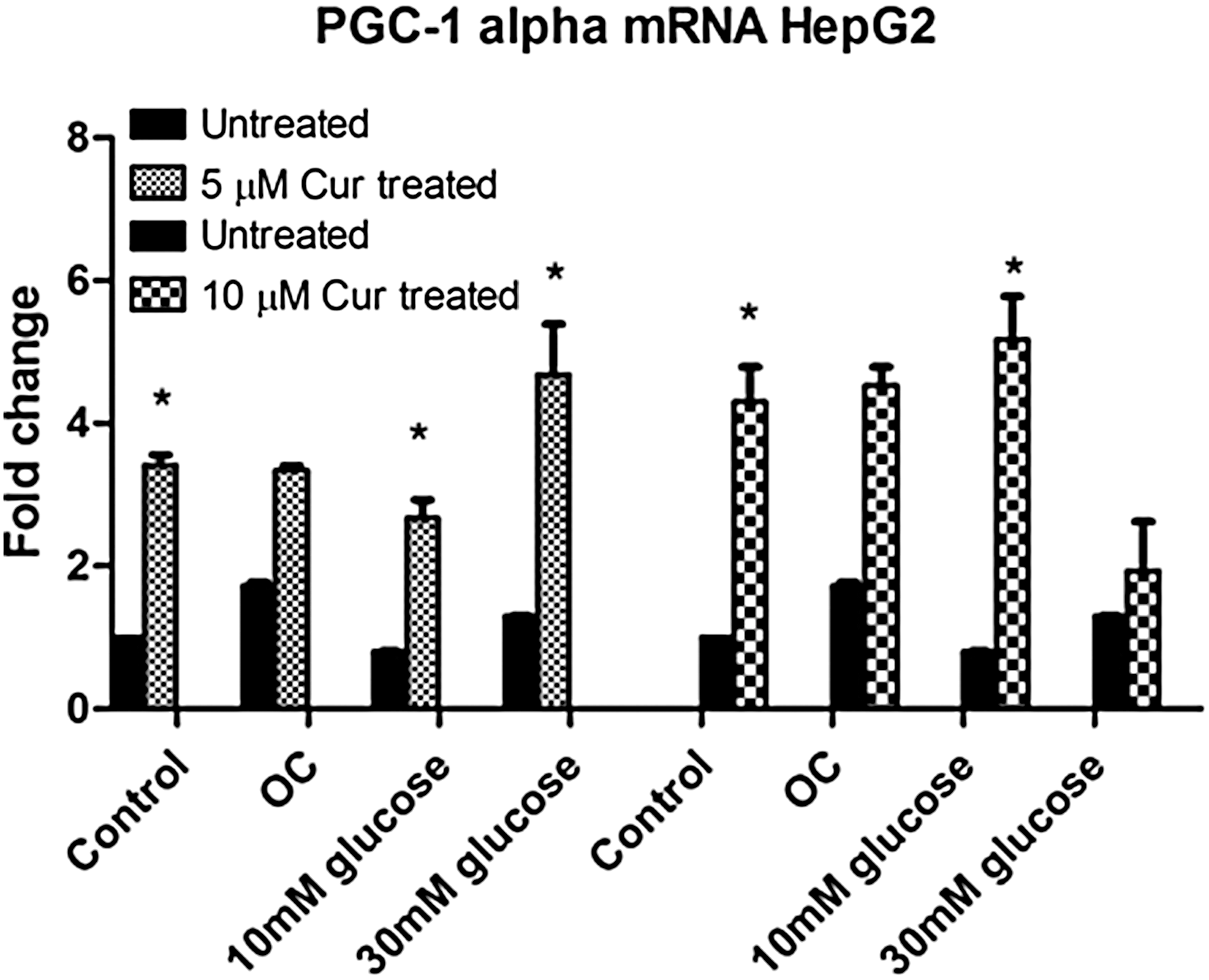
The effect of curcumin on PGC-1α expression in untreated and curcumin-treated HepG2 cells. Gene expression for PGC-1α was assessed. Values are expressed as fold changes relative to the control. Each bar represents the mean ± SEM of three replicates. *P < .05 versus untreated control, P < .05 versus curcumin-treated control. PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1 alpha.
When compared with the treated control, the intermediate hyperglycemic group also showed significantly higher PGC-1α expression (1.2-fold, P = .0077; Fig. 4).
Curcumin increases AO defense under normo- and hyperglycemic conditions
To determine the effect of curcumin treatment on AO activity in HepG2 cells, gene expressions for GPx1 and SOD2 were determined via qPCR.
Increased expressions of GPx1 and SOD2 were observed in the curcumin-treated hyperglycemic groups.
GPx1 levels were higher in the curcumin-treated control and the curcumin-treated hyperglycemic groups relative to the untreated control (5 μM [control: P = .0009; 10 mM: P = .0228; 30 mM: P = .0095] and 10 μM [control: P = .0019; 10 mM: 3.1-fold, P = .0098; 30 mM: 3.2-fold, P = .0024]; Fig. 5A). A similar pattern was observed with SOD2 mRNA expression.
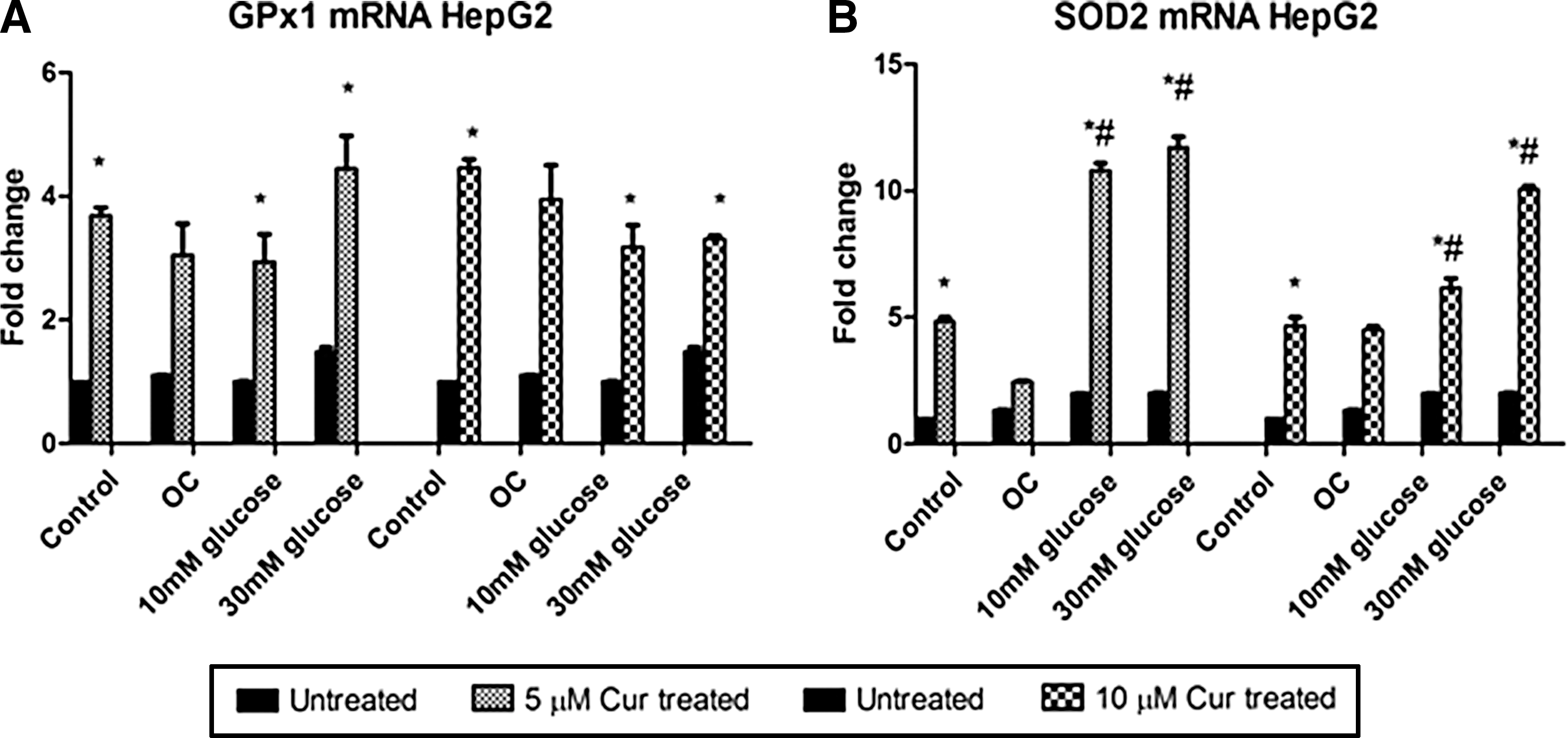
The effect of curcumin on antioxidant mRNA expression in untreated and curcumin-treated HepG2 cells. Gene expressions for
SOD2 levels were significantly higher in the treated hyperglycemic groups relative to both the untreated and treated controls in the 10 μM group (10 mM: 6.1-fold, P = .0019; 30 mM: 10-fold, P < .0001 and 10 mM: 1.4-fold, P = .0008; 30 mM: 2.2-fold, P = .0026, respectively; Fig. 5B).
Curcumin increases NF-κB, Lon protease, and Hsp70 expressions under normo- and hyperglycemic conditions
We investigated the effect of curcumin on NF-κB protein expression in HepG2 cells. To determine the effect of curcumin on protein quality control, both Lon and Hsp70 protein expressions were measured.
The NF-κB protein expressions in the 10 μM curcumin-treated hyperglycemic groups were significantly increased relative to the untreated and treated controls (10 mM: 2-fold, P = .0052; 1.7-fold, 30 mM: P = .0002 and 10 mM: 1.3-fold, P = .0468; 30 mM: 1.1-fold, P = .2509, respectively; Fig. 6A).
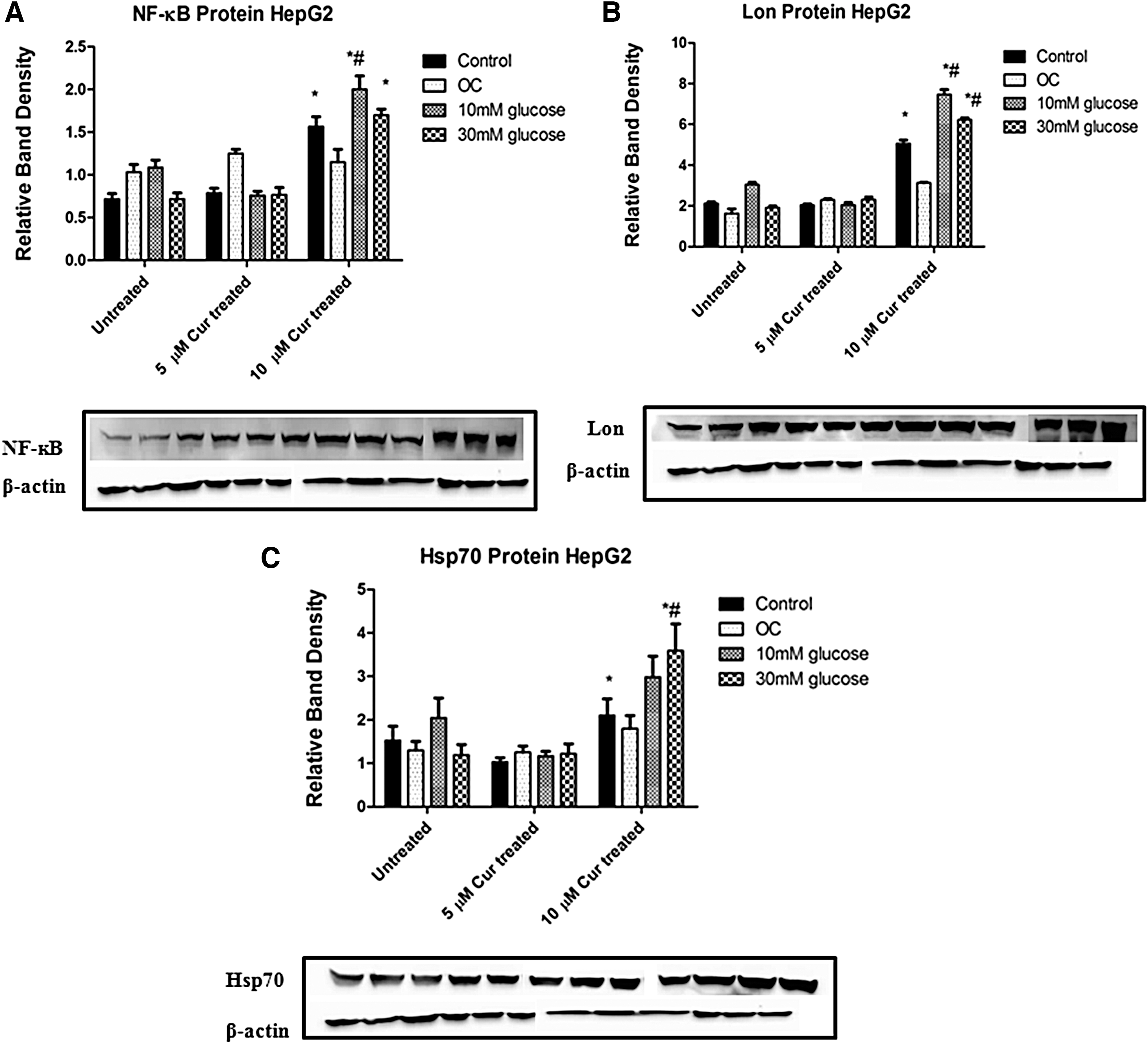
The effect of curcumin on NF-κB, Lon protease, and Hsp70 expressions in untreated and curcumin-treated HepG2 cells.
Lon protein expression in the 10 μM curcumin-treated hyperglycemic groups was higher relative to the untreated and treated controls, and these differences were statistically significant (10 mM: 75-fold, P = .0021; 30 mM: 6.2-fold, P < .0001 and 10 mM: 1.5-fold, P = .0199; 30 mM: 1.24-fold, P = .0053, respectively; Fig. 6B).
The 10 μM curcumin-treated groups also showed higher protein expression of Hsp70 relative to the untreated and treated controls but only the 30 mM groups reached significance in both (P = .0220 and P = .0233, respectively; Fig. 6C). Protein expressions of both Lon and Hsp70 in the 5 μM Curcumin-treated groups did not differ significantly from the untreated control.
Discussion
Mitochondria are the primary source of ROS formed from the ETC. Excessive ROS leads to oxidative damage of macromolecules, which play a role in DMII.
Mitochondrial DNA is vulnerable to ROS-induced damage as it is located closely to the ETC, it lacks protective histones and has limited DNA repair activity. 17 Excessive ROS impairs OXPHOS, leading to decreased mitochondrial function, decreased ATP levels, 18 and a marked reduction in AO activity. 19 These factors need to be improved to ameliorate disease conditions.
OS may be reduced through the use of exogenous compounds with AO properties that may prevent mitochondrial dysfunction. Curcumin is a phytochemical compound extracted from the rhizome of C. longa. It is a bifunctional AO as it reacts directly with ROS and RNS and also indirectly induces upregulation of cytoprotective and AO proteins such as SOD2, GPx1, and GSH. 7 The presence of phenolic and β-diketone functional groups in the structure of curcumin allows it to scavenge superoxide anions, hydrogen peroxide, hydroxyl radicals, and nitric oxide. 7,20,21
Curcumin also induces endogenous AO defense by modulating Nrf2. 22 Curcumin modifies the inhibitor of Nrf2, allowing for the translocation of Nrf2 to the nucleus where it binds to the AO response element in DNA to initiate transcription of AO genes. Curcumin has been shown to significantly ameliorate experimental diabetes and restore pancreatic AO defense. 23
Mitochondria are targets for protection by curcumin under oxidative conditions. 8 In a kidney model, curcumin maintained mitochondrial function by reducing ROS production and lipid peroxidation. 10 Curcumin also increased mitochondrial biogenesis in obese mice with liver steatosis 24 and reduced oxidative-induced mitochondrial damage in mice neuronal cells. 25
Curcumin has been shown to increase the levels of SIRT3, which forms part of an intrinsic network of regulated stress-response mechanisms. 26 We previously showed increased expressions of SIRT3 and AO defense enzymes under long- and short-term hyperglycemic stimulation. We established that SIRT3 may be the central component that modulates AO defense and confers resistance to OS-induced damage under hyperglycemic conditions in the HepG2 and HEK 293 cell lines. 27
Since both curcumin and SIRT3 target mitochondrial function, we postulated that SIRT3 may have an effect on the protective effects of curcumin under short-term hyperglycemic stimulation in HepG2 cells.
As a marker of increased mitochondrial output, we investigated cell viability following curcumin treatment using the MTT assay (measures the reduction of tetrazolium salt by succinate dehydrogenase in metabolically active cells). The level of activity is a measure of cell viability and mitochondrial function. We observed increased cell viability in all curcumin-treated groups with even higher levels in the curcumin-treated hyperglycemic groups. This suggests that curcumin maintains mitochondrial function under oxidative conditions. There was a decrease in cell viability in the 10 μM curcumin-treated hyperglycemic (30 mM) group. Curcumin has been shown to induce ROS generation in various cell lines 28,29 and this, together with the high glucose concentration, may have resulted in decreased cell proliferation.
SIRT3 induces AO genes by PGC-1α, a transcriptional coactivator that is activated in response to OS. 30 It regulates mitochondrial function and AO defense enzymes such as SOD2 (detoxifies superoxide radicals formed as a byproduct of OXPHOS) and GPx1 (enzyme that detoxifies H2O2 to H2O). 31 –33
In an oxidative environment, PGC-1α induces SIRT3 expression through activation of estrogen-related receptor-α, and in turn, SIRT3 stimulates PGC-1α expression via phosphorylated CREB, thereby forming a positive feedback cycle and increasing AO defense. 34,35
Curcumin increased expressions of SIRT3, CREB, PGC-1α, and AO enzymes in the hyperglycemic treatment groups following 5 and 10 μM curcumin administration. Although the expression levels were differentially regulated by both concentrations of curcumin under hyperglycemic conditions, the more pronounced effect was observed in the 10 μM curcumin group.
Interestingly, we found a distinct increase in NF-κB expression in the 10 μM curcumin-treated hyperglycemic groups. Studies have shown that curcumin inhibits NF-κB activity, as it is a key transcription factor involved in the inflammatory response. 36,37 Curcumin, however, has also been shown to induce ROS generation in various cell lines, 28,29 and this may modulate the NF-κB response, which is mainly involved in cell survival.
The main role of NF-κB is considered to be protective and to initiate a highly coordinated response in multiple cell types to effectively counteract OS. 38 A study showed that OS induced by reperfusion insult, in cultured neurons, was associated with NF-κB, inducing a protective signal rather than death signals. 39 Other studies have also shown that activation of NF-κB led to neuroprotection, and NF-κB inhibition led to apoptosis. 40,41 It has also been shown that NF-κB is deacetylated and activated by SIRT3. Activation of NF-κB by SIRT3 increased the expression of SOD2, thus making cells resistant to OS-mediated cell damage. 13 The target genes of NF-κB may therefore vary depending on the cellular context.
Oxidative damage can form adducts of proteins with carbohydrates and lipids and may create carbonyl groups and intramolecular cross-links. These may impair enzymes and structural proteins which, if not removed, can aggregate and cause significant cellular toxicity. Mitochondrial proteins are at an increased risk of oxidative damage.
Lon is the major protease in the mitochondrial matrix that may be regulated by NF-κB and SIRT3. 42,43 Lon degrades oxidized and damaged proteins and acts as a chaperone by assisting the assembly and folding of mitochondrial proteins. A study has shown that Lon induction significantly increased protection against the accumulation of carbonylated proteins and increased cell viability during OS. 44
We found increased expressions of NF-κB and Lon in the 10 μM curcumin-treated hyperglycemic groups, similar to SIRT3 expression, which could suggest regulation of Lon by NF-κB and SIRT3. No differences were observed in the groups treated with 5 μM curcumin when compared with the untreated group.
We also observed increased Hsp70 expression in the 10 μM curcumin-treated hyperglycemic groups. Hsp70 is most closely linked to cytoprotection and has the ability to prevent protein aggregation and other protein modifications. It may also prevent oxidative injury through improving cell protein and endogenous AO stability. 45 Hsp70 has been shown to be part of the protein quality control and cell-protective network with Lon in the recognition and removal of damaged proteins.
Following curcumin administration, there were increased expressions of SIRT3, AO defense, and protein quality control under hyperglycemic conditions, notably in the 10 μM curcumin-treated hyperglycemic groups. Furthermore, the 10 mM glucose treatment yielded higher SIRT3 and AO expressions than the 30 mM glucose treatment following 10 μM curcumin administration. These results suggest that selective concentrations of curcumin may improve the AO response and cell survival under intermediate hyperglycemic conditions.
Since SIRT3 has already been shown to increase AO defense and cell survival, the protective effects of curcumin may occur through SIRT3-mediated mechanisms.
This study may be improved by using a diabetic in vivo model, which may help to establish a more holistic response. A wider range of curcumin concentrations over a chronic time period may also help better elucidate the effect of curcumin in a diabetic model. Inhibition of SIRT3 is required to determine the role of SIRT3 in the protective effects of curcumin.
Future studies include interrogating a more defined cell survival pathway following curcumin administration. Investigating the effect of curcumin on the epigenome may enable the determination of differential gene regulation during prediabetes and DMII.
In conclusion, mitochondrial dysfunction and OS play a key role in the pathogenesis of several metabolic disorders. Curcumin targets AO defense and may improve disease conditions by conferring resistance to OS-induced damage through SIRT3-mediated mechanisms.
Footnotes
Acknowledgments
The authors thank the National Research Foundation and UKZN (CHS Strategic funds) for funding. We also acknowledge the assistance of Dr. D. Moodley (Harvard Medical School).
Author Disclosure Statement
No competing financial interests exist.