
Abstract
High levels of alpha-tocopherol, the usual vitamin E supplement, are reported to decrease bone mass in rodents; however, the effects of other vitamin E forms on the skeleton are unknown. To test the hypothesis that high intakes of various vitamin E forms or the vitamin E metabolite, carboxyethyl hydroxy chromanol, were detrimental to bone status, Sprague-Dawley rats (n = 6 per group, 11-week males) for 18 weeks consumed semipurified diets that contained adequate alpha-tocopherol, high alpha-tocopherol (500 mg/kg diet), or 50% Tocomin (250 mg mixed tocopherols and tocotrienols/kg diet). Vitamin E status was evaluated by measuring plasma, liver, and bone marrow vitamin E concentrations. Bone density, microarchitecture (cross-sectional volume, cortical volume, marrow volume, cortical thickness, and cancellous bone volume fraction, trabecular number, thickness, and spacing), and cancellous bone formation were assessed in the tibia using dual-energy X-ray absorptiometry, microcomputed tomography, and histomorphometry, respectively. In addition, serum osteocalcin was assessed as a global marker of bone turnover; gene expression in response to treatment was evaluated in the femur using targeted (osteogenesis related) gene profiling. No significant differences were detected between treatment groups for any of the bone endpoints measured. Vitamin E supplementation, either as alpha-tocopherol or mixed tocotrienols, while increasing vitamin E concentrations both in plasma and tissues, had no effect on the skeleton in rats.
Introduction
O
Vitamin E is lipid soluble and functions as a chain-breaking antioxidant, preventing propagation of lipid peroxidation and damage to cellular membranes. 10,11 In addition to its antioxidant function, high levels of vitamin E have been shown to decrease bone mass in rodent models. Fujita et al. 9 found that growing mice fed diets fortified with the alpha-tocopherol form of vitamin E (600 mg/kg diet) for 8 weeks had 20% lower cancellous bone volume. In addition, they observed that growing rats fed similar diets for 8 weeks had significantly lower BMD compared with rats consuming normal levels of vitamin E. Furthermore, their in vitro studies using isolated mouse bone marrow cells revealed that alpha-tocopherol induces the expression of dendritic cell-specific transmembrane protein, which promotes osteoclast fusion and maturation. These findings suggest that high-dietary intakes of vitamin E may slow bone accrual during growth and potentially accelerate age-related bone loss in adults, which poses a threat to people consuming vitamin E supplements.
The most frequently consumed vitamin E supplement in the United States contains alpha-tocopherol; 11.3% of adults consume single vitamin E supplements (>400 IU/day), and more than 30% consume multisupplements containing vitamin E. 12 In addition to alpha-tocopherol, other forms of vitamin E, such as tocotrienols, are found in supplement form. Alpha-tocopherol is the most biologically active form of vitamin E and its concentrations are maintained in circulation by the function of the hepatic alpha-tocopherol transfer protein. 11 By contrast, tocotrienols are absorbed and transported to the liver, where they are metabolized to the water-soluble metabolite, carboxyethyl hydroxy chromanol (CEHC). 10 CEHCs are water soluble for excretion in urine, are detectable in tissues, 13 and are more potent anti-inflammatory agents than are tocopherols. 14,15 Importantly, liver alpha-tocopherol, at high concentrations, is also metabolized to CEHC, but to a lesser extent compared with tocotrienols. 11
Vitamin E supplements have been viewed as safe and protective against increased oxidative stress 12 ; however, recent evidence has shown that alpha-tocopherol may reduce bone accrual and thus may lower peak bone mass. 9 In addition, an increase in bone resorption is problematic for older populations who are already at risk for osteoporosis. 1 It is unknown whether these adverse skeletal effects are specific to alpha-tocopherol or if other vitamin E supplements also impact bone loss. We hypothesize that the in vivo effects of vitamin E on bone metabolism involve not only elevated tocol concentrations but also are dependent on increased CEHC concentrations. We sought to investigate the mechanisms for the potential adverse consequences of vitamin E on bone by evaluating both alpha-tocopherol and non-alpha-tocopherols, and their CEHC metabolites on bone mass, density, microarchitecture, and turnover. Herein, we examined the effects of long-term vitamin E supplementation on the rodent skeleton, using two common vitamin E supplements, alpha-tocopherol and mixed tocols (e.g., 50% Tocomin). We hypothesize that 50% Tocomin will have lower adverse skeletal effects due to its rapid metabolism into CEHC. 16 Given these proposed findings, individuals will be able to better select a vitamin E supplement.
Materials and Methods
Animal study design
The Oregon State University (OSU) Institutional Animal Care and Use Committee approved all procedures, and the institutional guide for the care and use of laboratory animals was followed. Ten-week male Sprague-Dawley rats (Charles River) were housed in plastic cages with hard wood chips, kept on a 12-h light/12-h dark cycle, and maintained on an AIN-93G diet (Harlan) and water ad libitum for 7 days to acclimate to the OSU animal facility. The rats were housed two per cage throughout the study.
Experimental diets were based on AIN-93G (Harlan) including (1) adequate-dietary alpha-tocopherol (A-AT, 60 IU all-rac-alpha-tocopheryl acetate/kg diet), (2) high-dietary alpha-tocopherol (H-AT, added 500 mg RRR alpha-tocopherol/kg diet), or (3) high-dietary mixed tocols (H-T3, added 500 mg 50% Tocomin/kg diet with tocols 50% by weight). Rats were randomly assigned to one of the three treatment groups (A-AT, n = 6; H-AT, n = 6; H-T3, n = 7). The rats were maintained on their respective diets for 18 weeks.
To label mineralizing bone for assessment of formation, the rats were injected with calcein, a fluorochrome (15 mg/kg body weight; Sigma Chemical) at 11 and 3 days before euthanasia. At necropsy, blood was collected in 10-mL vacutainer tubes containing 1 mg/mL ethylene diamine tetraacetic acid (Becton, Dickinson and Company). Plasma was obtained by centrifugation (1500 g, 15 min) and stored at −80°C until analysis. Livers were excised, frozen in liquid nitrogen, and stored at −80°C. Tibiae and lumbar vertebrae were excised and placed in 70% ethanol for dual-energy X-ray absorptiometry (DXA), microcomputed tomography (μCT), and histomorphometry analyses. Right femora were removed, cleaned of soft tissue, frozen in liquid nitrogen, and stored at −80°C for mRNA isolation and gene expression analysis. Bone marrow from left femora was collected and homogenized in PBS, frozen in liquid nitrogen, and stored at −80°C.
Vitamin E and CEHC analysis
Diet, plasma, bone marrow, and liver vitamin E concentrations were determined by high-performance liquid chromatography (HPLC) with electrochemical detection as previously described. 17 Briefly, diet, plasma, bone marrow, or liver was saponified with ethanolic KOH, extracted with hexane, dried under nitrogen, resuspended in ethanol:methanol (1:1), and injected into the HPLC system. Vitamin E was detected electrochemically in oxidizing mode with 500 mV potential. Peak areas were integrated using Shimadzu Scientific 4.2 Class VP software package. Values were quantitated using calibration standards prepared using authentic vitamin E compounds. The working linear range for tocotrienol quantitation was 2–200 pmol injected and the lower limit of detection was 1 pmol injected (this is approximately equivalent to 0.06 μmol tocotrienol/L plasma or 0.01 nmol/g tissue).
Plasma, bone marrow, and liver alpha- and gamma-CEHC concentrations were determined by HPLC with mass spectrometry as previously described. 18,19 Plasma and bone marrow CEHCs were extracted using diethyl ether from acid-hydrolyzed samples 19 ; livers were homogenized and then CEHCs were extracted using diethyl ether from sulfatase/β-glucuronidase-hydrolyzed samples. 18 Aliquots of ether fractions were collected and dried under nitrogen. The samples were resuspended in 1:1 (v/v) water:methanol containing Trolox (Sigma) as the internal standard and injected using liquid chromatography/mass spectrometry with a Waters 2695 Separations Module and a Micromass ZQ2000 with an electrospray ionization probe (Waters). Concentrations were calculated by comparison to peak areas generated from a standard curve using authentic compounds with correction based on the internal standard, Trolox.
Densitometry
Total tibia bone mineral content (BMC, g), bone area (cm2), and BMD (g/cm2) were measured using DXA (PIXImus 2; Lunar Corporation).
μCT analysis
Tibiae and second lumbar vertebrae were scanned using a Scanco μCT40 scanner (Scanco Medical AG) at a voxel size of 16 × 16 × 16 μm (55 kVp X-ray voltage, 145 μA intensity, and 200 msec integration time). Filtering parameters sigma and support were set to 0.8 and 1, respectively. The threshold value for evaluation was determined empirically and set at 245 (gray scale, 0–1000). Cortical bone in the tibia diaphysis and cancellous bone in the proximal tibia metaphysis and epiphysis were evaluated (Fig. 1). Automated contouring was used to delineate cortical bone from nonbone. Following, all cortical slices were examined visually for inclusion of cancellous struts originating from the endocortex (extremely rare at this site) and manually removed when present. For cortical bone, 62 slices (992 μm in length) were evaluated and cross-sectional volume (cortical and marrow volume, mm3), cortical volume (mm3), marrow volume (mm3), and cortical thickness (μm) were determined. For proximal tibia metaphysis, 50 slices (800 μm in length) of cancellous bone were measured starting 800 μm distal to the growth plate. The entire cancellous compartment (29 ± 1 slices, 465 ± 16 μm, mean ± standard error of the mean [SEM]) was assessed in the tibial epiphysis. Analysis of the lumbar vertebra included the entire region of secondary spongiosa between the cranial and caudal growth plates (282 ± 6 slices, 4512 ± 96 μm) (Fig. 1). Cancellous bone measurements in the tibia and lumbar vertebra included cancellous bone volume fraction (bone volume/tissue volume, %), trabecular thickness (mean thickness of individual trabeculae, μm), trabecular number (number of trabecular intersects per unit distance, mm−1), and trabecular spacing (distance between trabeculae, μm).
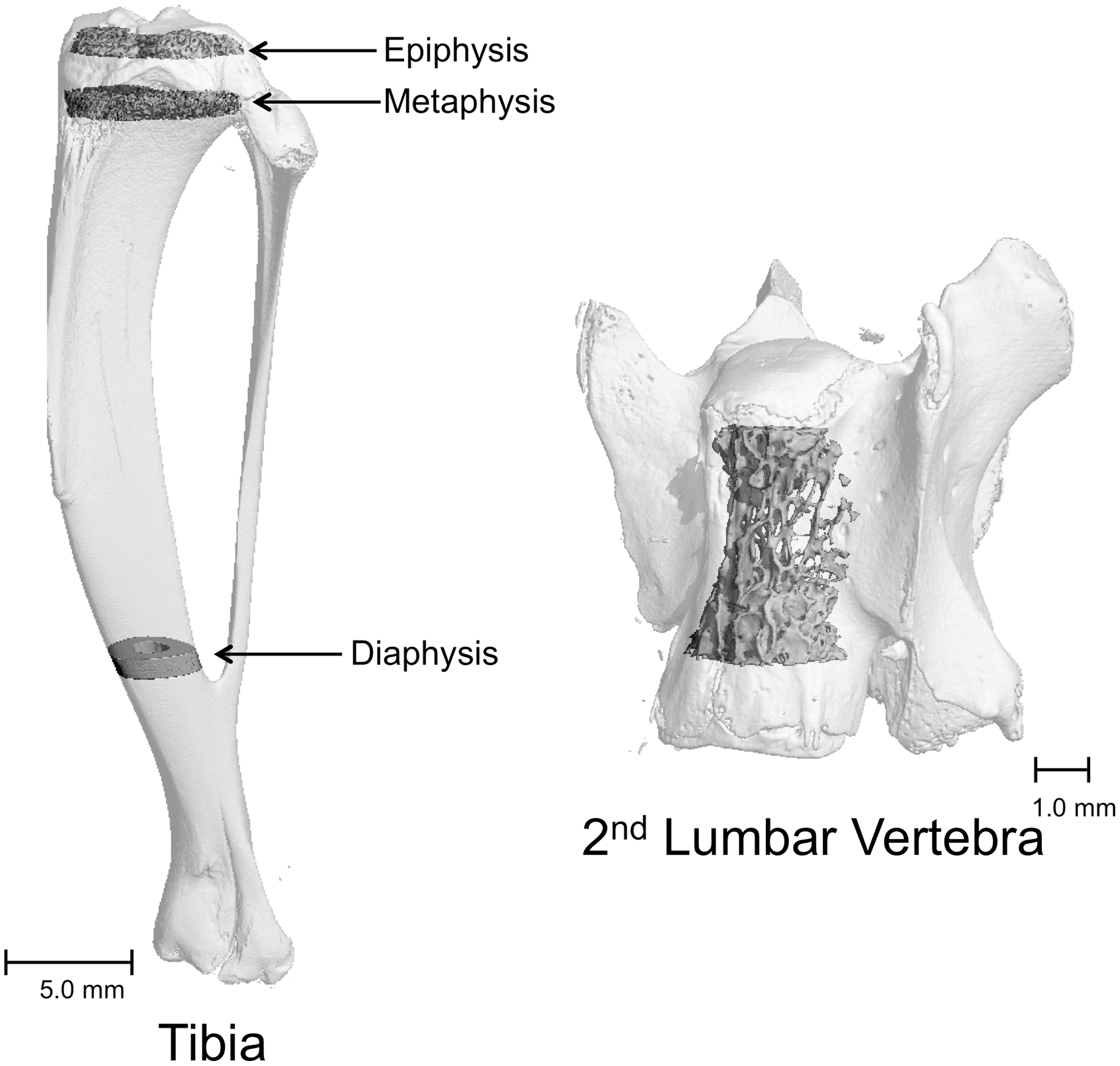
Tibia and lumbar vertebra sampling sites are indicated.
Histomorphometry analysis
Preparation of tibiae for histomorphometric evaluation was completed as previously described. 20 Briefly, the proximal tibia was dehydrated in a graded series of ethanol and xylene and embedded in modified methyl methacrylate. Sections (5 μm thick) were cut with a vertical bed microtome (Leica/Jung 2165) and affixed to slides precoated with a 1% gelatin solution. Sections were mounted unstained for measurement of fluorochrome labeling. The sampling site for tibial metaphysis was located ∼0.75–1.00 mm distal to the growth plate. Fluorochrome-based measurements of bone formation included (1) mineralizing perimeter (mineralizing perimeter/bone perimeter: cancellous bone perimeter covered with double plus half single label normalized to bone perimeter, %), (2) mineral apposition rate (the distance between the two fluorochrome (calcein) markers that comprise a double label divided by the 8 day interlabel interval, μm/day), and (3) bone formation rate (bone formation rate/bone perimeter: calculated by multiplying mineralizing perimeter by mineral apposition rate normalized to bone perimeter, μm2/μm/year). All data were collected using the OsteoMeasure System (OsteoMetrics, Inc.).
Serum osteocalcin assay
Serum gla-osteocalcin was measured using the MK126 rat Gla-osteocalcin high-sensitivity enzyme immunoassay kit from Clontech.
Gene expression analysis
Femora were pulverized with a mortar and pestle in liquid nitrogen and then homogenized in TRIzol (Invitrogen). Total RNA was isolated according to the manufacturer's protocol, and mRNA was reverse transcribed into cDNA using SuperScript III First-Strand Synthesis SuperMix for quantitative reverse transcription–polymerase chain reaction (Invitrogen). Osteogenic gene expression was determined using a rat osteogenesis RT2 profiler PCR array (Qiagen) according to the manufacturer's protocol. Gene expression was normalized to beta-2 microglobulin, hypoxanthine phosphoribosyltransferase 1, and lactate dehydrogenase A, and relative quantification was determined by the delta delta Ct method using RT2 Profiler PCR Array Data Analysis software version 3.5 (Qiagen).
Statistical analysis
One-way analysis of variance (ANOVA) followed by Tukey post-hoc test was used to evaluate differences among treatment groups (Prism 6; GraphPad software). ANOVA assumptions of equal variances between groups were met as evidenced by Bartlett's test. Differences were considered significant at P < .05. All data are expressed as mean ± SEM.
Results
Tocopherol and tocotrienol concentrations in plasma, bone marrow, and liver
Sprague-Dawley rats, 11 weeks old, were fed semipurified diets containing A-AT, H-AT, or H-T3 for 18 weeks. Animal weights increased over the duration of the study (not shown). Final average weights were not significantly different among groups (A-AT = 671 ± 35, H-AT = 682 ± 64, H-T3 = 679 ± 58 g). H-AT diets contained 518 mg alpha-tocopherol/kg, while the H-T3 diets contained 234 mg mixed tocopherols and tocotrienols/kg diet. Both of these diets contained gamma-tocopherol and gamma-tocotrienol, but H-AT diets had four times more gamma-tocopherol and the H-T3 diet 40-times more gamma-tocotrienol (Table 1). Thus, the experimental conditions provided nearly a sevenfold range of alpha-tocopherol intakes and a variety of other vitamin E forms to assess varying tocol intakes on bone metabolism.
A-AT, adequate-dietary alpha-tocopherol; H-AT, high-dietary alpha-tocopherol; H-T3, high-dietary mixed tocols.
Plasma and liver alpha-tocopherol concentrations approximately doubled in rats consuming the H-AT diet compared with rats consuming the A-AT or H-T3 diets (Fig. 2A, C). Significant differences in bone marrow alpha-tocopherol were not detected with treatment (Fig. 2B). Liver gamma-tocopherol concentrations were also higher in rats consuming the H-AT diet compared with the H-T3 diet (Fig. 2F). However, bone marrow alpha-tocopherol and gamma-tocopherol concentrations in plasma and bone marrow were unchanged with any of the dietary treatments (Fig. 2B, D, and E). The H-T3 diet with its ∼10-fold higher tocotrienol concentrations increased alpha- and gamma-tocotrienols in the rats' plasma, bone marrow, and liver (Fig. 2G–L). As expected, tocotrienols were not detected in samples from rats fed the A-AT and H-AT diets.
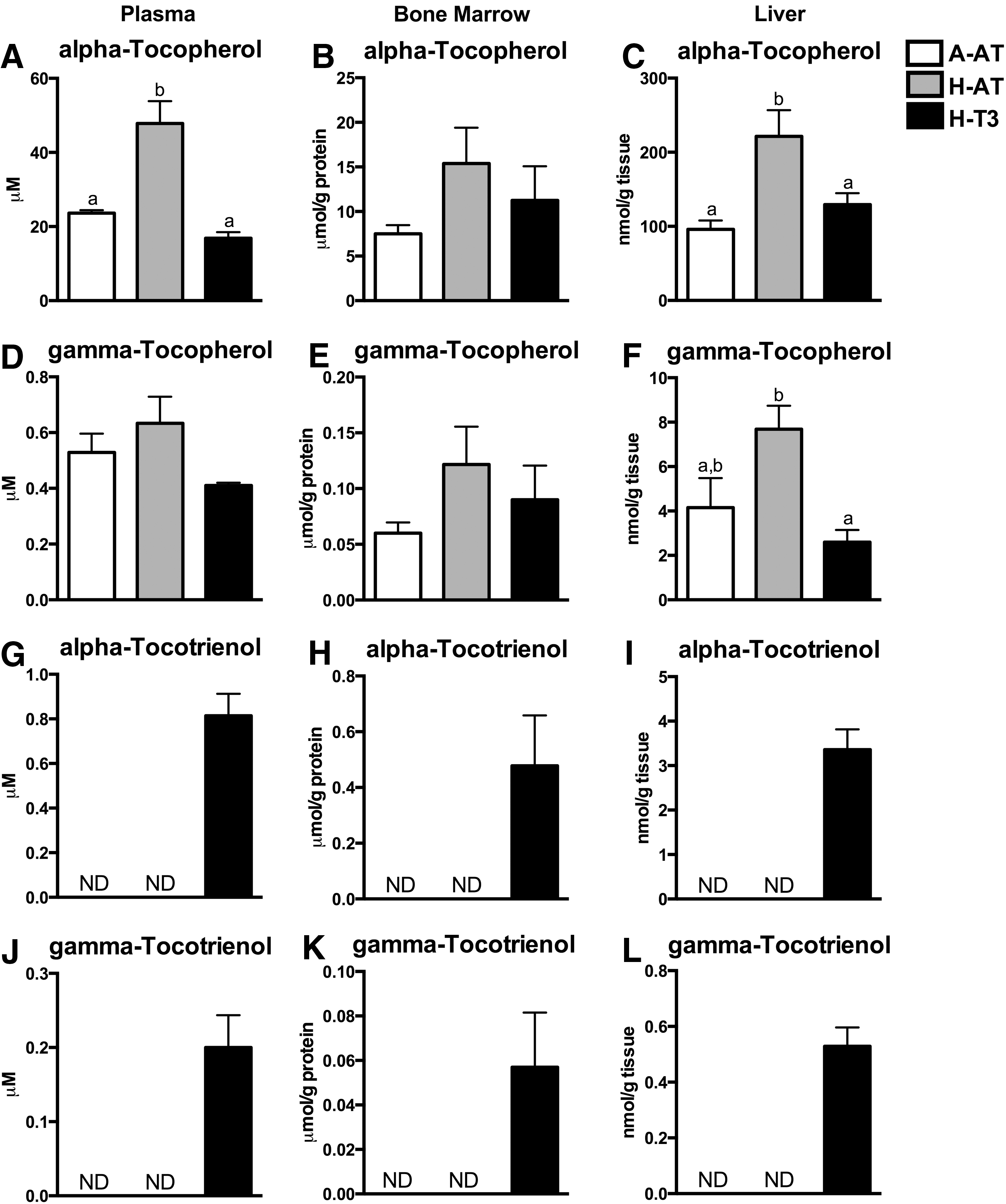
Vitamin E concentrations in plasma, bone marrow, and liver. Alpha-tocopherol concentrations
Alpha-CEHC concentrations in plasma, bone marrow, and liver were fivefold higher in rats fed the H-AT diets and twofold higher in rats fed the H-T3 diet compared with the rats fed A-AT diets (Fig. 3A–C). In addition, plasma and liver concentrations of gamma-CEHC were significantly increased in the H-AT and H-T3 diet groups compared with the A-AT group (Fig. 3D, F). Bone marrow gamma-CEHC concentrations were unaffected by dietary treatments (Fig. 3E).
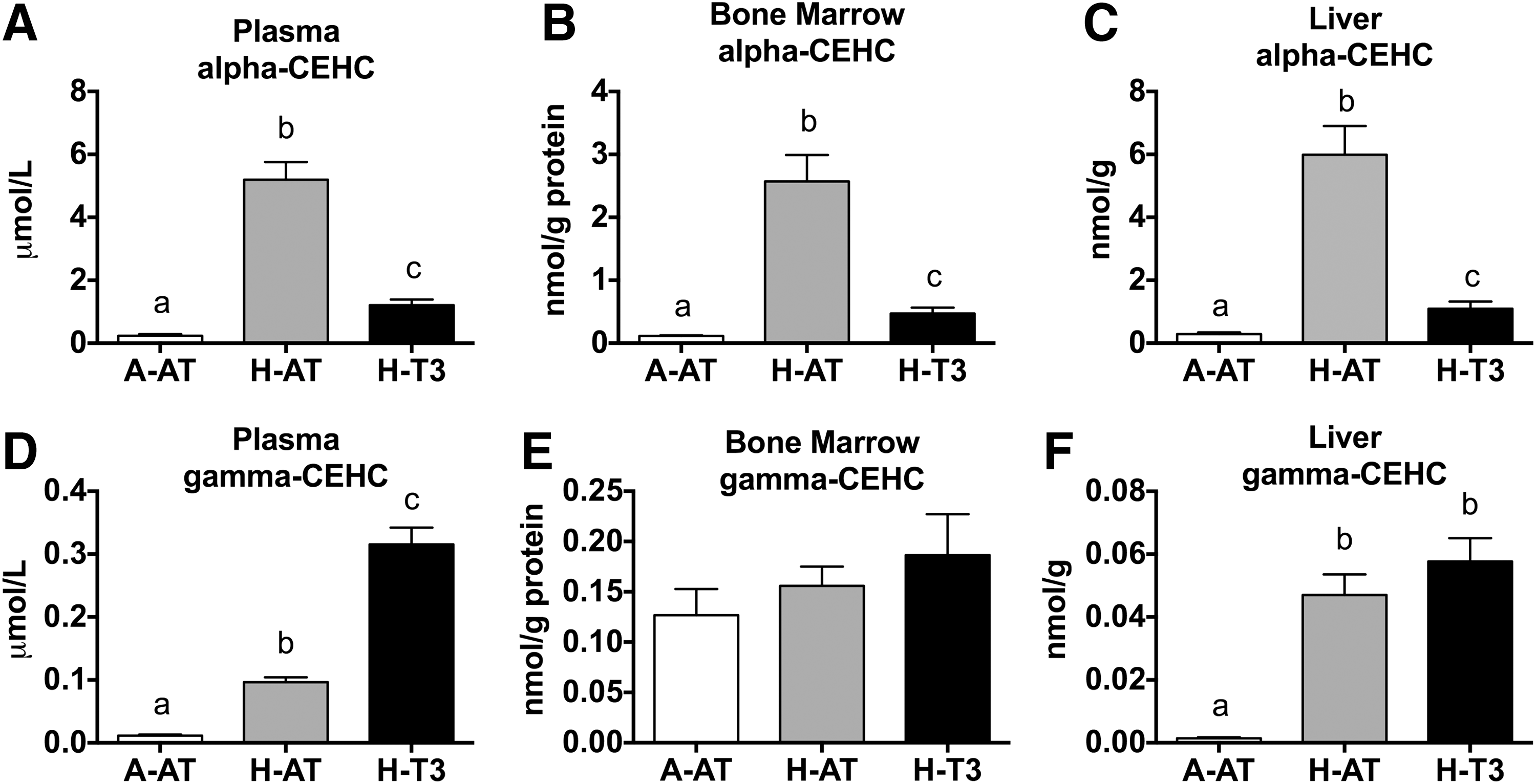
High intakes of dietary vitamin E increased plasma, bone marrow, and liver CEHC concentrations.
Bone mass and bone microarchitecture
Bone mass of tibia was assessed using DXA. No significant differences among diet groups were found for BMC, bone area, or BMD (Fig. 4). Bone microarchitecture was assessed using μCT analysis of tibia diaphysis, epiphysis, metaphysis, and second lumbar vertebra. The rats consuming A-AT, H-AT, or H-T3 diets did not demonstrate any significant differences with regard to tibia diaphysis cross-sectional volume, cortical volume, marrow volume, or cortical thickness (Fig. 5 and Supplementary Table S1; Supplementary Data are available online at
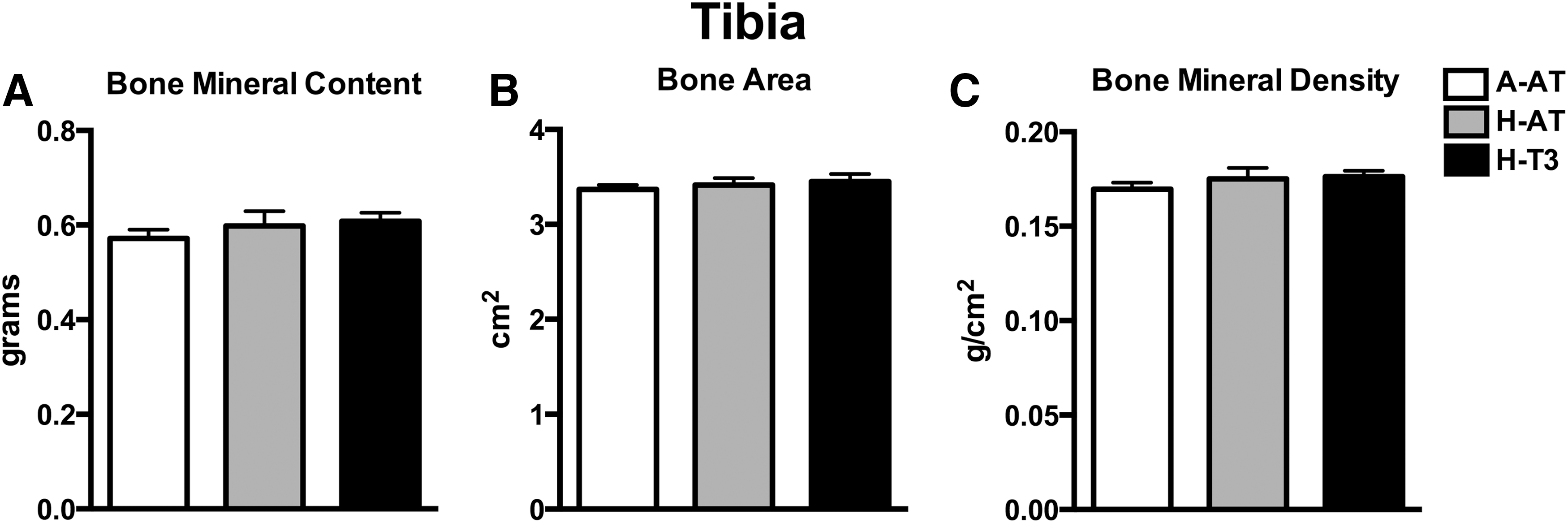
High intakes of dietary vitamin E had no effect on bone mineral content and density. Tibia bone mineral content
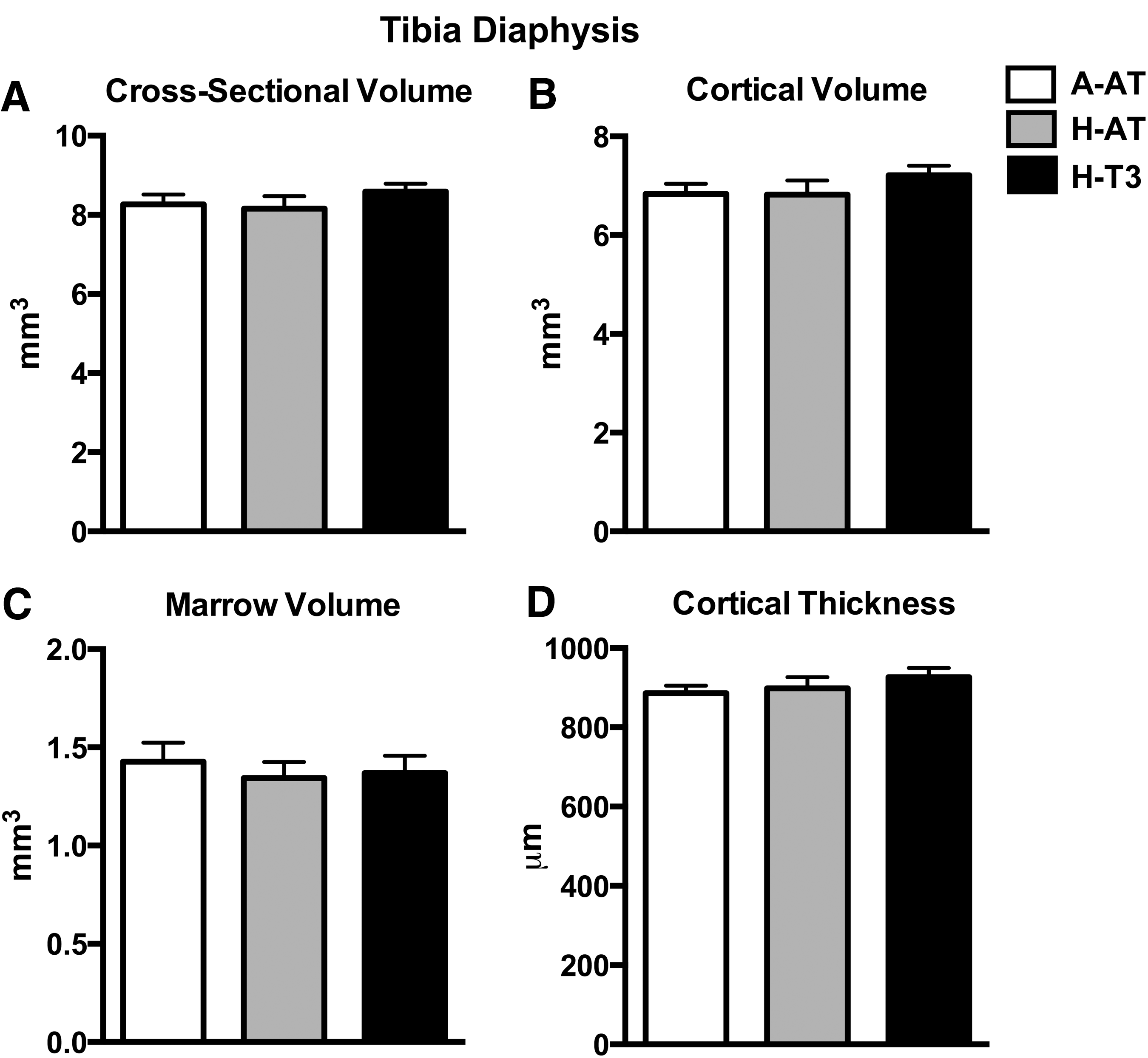
High intakes of dietary vitamin E had no effect on cortical bone microarchitecture. Tibia diaphysis cross-sectional volume
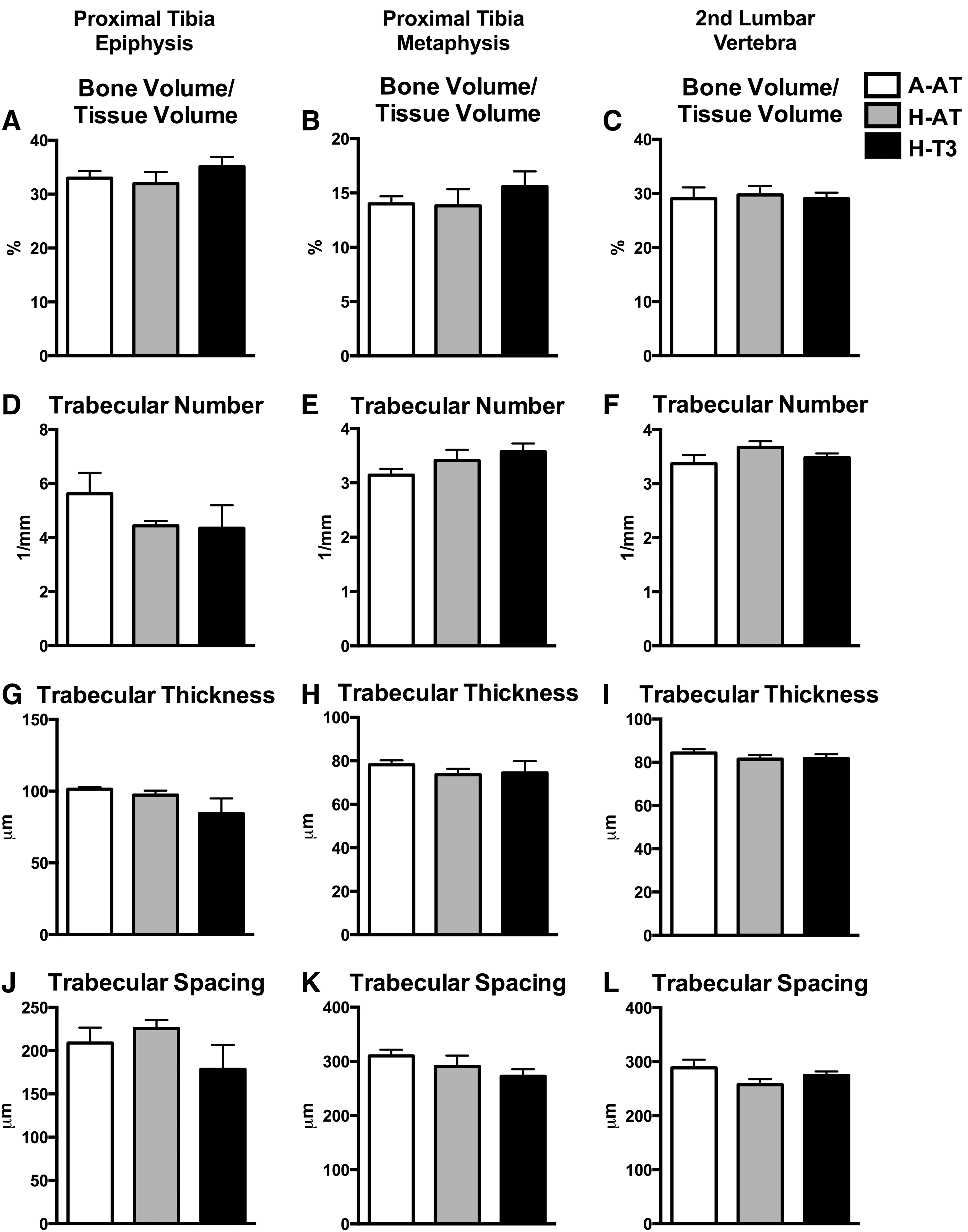
High intakes of dietary vitamin E had no effect on cancellous bone microarchitecture. Tibia epiphysis
Bone formation
Bone formation was assessed using histomorphometry and measuring osteocalcin, a global serum marker of bone turnover. No significant differences were observed among diet groups for mineralizing perimeter, mineral apposition rate, or bone formation rate in tibia metaphysis (Fig. 7A–C). Also, there were no significant differences in serum osteocalcin among diet groups (Fig. 7D).
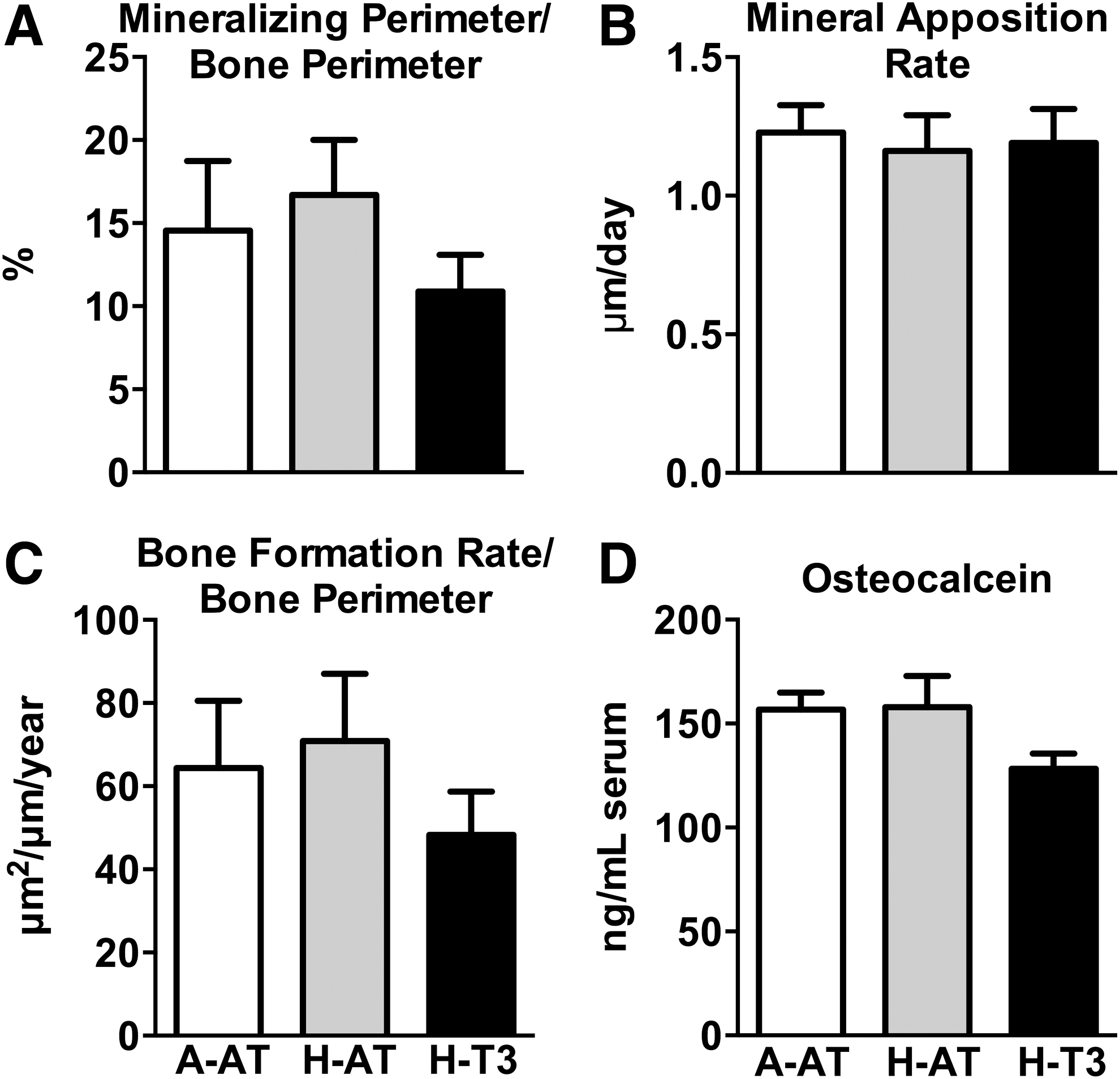
High intakes of dietary vitamin E had no effect on bone formation and turnover. High intakes of vitamin E had no effect on mineralizing perimeter
Bone marrow osteogenic gene expression
Femora from rats consuming A-AT, H-AT, and H-T3 diets were assessed with regard to gene expression using an 84-gene PCR array, focusing on genes related to osteogenesis and bone turnover. Femurs from rats consuming the H-AT diet did not show any genes that are differentially expressed compared with femurs from rats consuming the A-AT diet (Fig. 8A). However, femurs from rats consuming the H-T3 diet were found to have some alterations in osteogenic gene expression compared with those from rats consuming the A-AT diet (Fig. 8B). Genes that were significantly decreased include annexin V (−0.2-fold), growth differentiation factor-10 (−1.0-fold), matrix metalloproteinase-8 (−0.5-fold), mothers against decapentaplegic homolog-5 (−0.4-fold), and osteopontin (−0.6-fold). Genes that were significantly increased include bone morphogenic protein receptor, type 1B (1.8-fold), fibroblast growth factor-2 (0.5-fold), and noggin (1.0-fold). Importantly, neither H-AT nor H-T3 diets had a significant effect on expression of key genes related to osteoblast differentiation (Runx2 and Sp7), osteoblast number and activity (osteocalcin, collagen 1a1, collagen 2a1, osteopontin, and alkaline phosphatase), or osteoclast differentiation (colony stimulating factor 1 and RANKL).
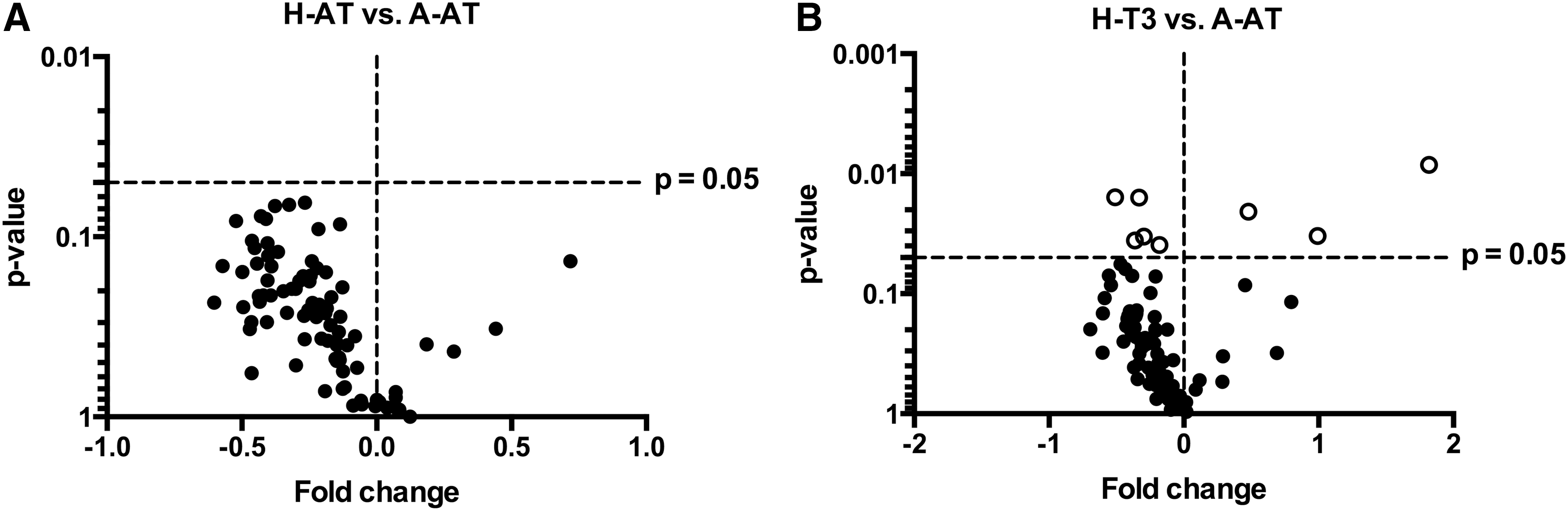
High intakes of dietary vitamin E had almost no effect on osteogenic gene expression. H-AT diet had no effect on targeted (osteogenesis related) gene profiling
Discussion
Rats were fed A-AT, H-AT, or H-T3 diets for 18 weeks, but no significant changes among diet groups were observed for tibial bone mass and density, cortical bone microarchitecture in tibia, cancellous microarchitecture at multiple skeletal sites, or global and local bone turnover. Changes in expression of genes related to osteogenesis and bone turnover in femur in response to treatment were minimal. Given these findings, we conclude that high-dietary levels of alpha-tocopherol or mixed tocols have little or no impact on the rodent skeleton.
Our findings are in contrast to the study reported by Fujita et al., 9 who fed diets containing 600 mg alpha-tocopherol/kg diet for 8 weeks to rapidly growing C57BL6 mice (4 weeks old) and rapidly growing Sprague-Dawley rats (8 weeks old). They observed a 20% lower lumbar vertebra cancellous bone volume fraction in mice and lower BMD in the femora of rats. However, other published reports 21 –24 have shown no adverse effects on the skeleton of rodents fed diets containing high levels of vitamin E. Iwaniec et al., 21 using skeletally mature Sprague-Dawley rats, fed high alpha-tocopherol diets (333 mg/kg diet) for 13 weeks and evaluated femoral bone mass, density, and microarchitecture. No significant differences were observed between control and high vitamin E-fed rats. The authors concluded that high-dietary alpha-tocopherol has no effect on the skeleton in skeletally mature rats. In addition, Arjmandi et al. 22 examined the effects of high-dietary vitamin E in adult (6 months old) and aged (24 months old) C56BL6 mice. Mice were fed for 4 weeks with 500 mg alpha-tocopherol/kg diet. The high vitamin E diet had no effect on mechanical bone parameters or serum markers of bone turnover in adult mice. By contrast, high-dietary vitamin E significantly improved bone strength in aged mice. The authors concluded that the high-dietary vitamin E diet had no effect on the skeleton in adult rodents and was beneficial for bone in aged rodents.
The discrepancy between outcomes relative to the effects of vitamin E on bone status may be due to the age of rodents studied. Iwaniec et al. 21 and Arjmandi et al. 22 used models whose skeletons were mature at the time the high vitamin E-containing diets were initiated, and therefore closely mimicked bone turnover during adulthood. 25 In adult animals, there is a gradual age-related cancellous bone loss. 26 Moreover, these models are more closely representative in age of humans who likely take vitamin E supplements. 27
In the present study, we chose to use 3-month-old Sprague-Dawley rats whose skeletons were growing slowly and had reached skeletal maturity by the end of the study. 25 We used this model to determine the effects of high levels of vitamin E on peak bone mass. Given our findings, we conclude that if high-dietary vitamin E slows bone acquisition during growth, it is temporary and not observed once rodents reach skeletal maturity. This conclusion is consistent with prior reports, 21 –24 in which high levels of vitamin E did not impact age-related bone loss when treatment was initiated in skeletally mature rodents.
In addition to the discussed rodent models, high-dietary vitamin E has also been examined in human clinical studies, showing no effect or modest beneficial effects on the skeleton. Ruiz-Ramos et al. 28 completed a clinical trial consisting of 135 subjects (mean age = 68) who received a placebo, 500 mg/day ascorbic acid +400 IU/day alpha-tocopherol, or 1000 mg/day ascorbic acid +400 IU/day alpha-tocopherol. After 1 year of supplementation, subjects receiving the high antioxidant dose had increased hip BMD compared with baseline measurements, and subjects receiving placebo or low-dose supplementation had no significant change in hip BMD from baseline. In addition, Chavan et al. 29 examined the effects of high-dietary vitamin E on serum markers of oxidative stress in 75 osteoporotic subjects (age range = 45–70). Subjects were given vitamin E (400 IU/day; Evinal), vitamin C (500 mg/day), or a combination for 90 days. Vitamin E supplementation significantly decreased serum levels of malondialdehyde in osteoporotic subjects compared with baseline measurements, suggesting an improvement in oxidative status from dietary vitamin E.
In the present study, we fed A-AT, H-AT, or H-T3 diets to assess additionally the role of the vitamin E metabolite, CEHC, on bone. As expected, high-dietary intakes of vitamin E significantly increased plasma and tissue CEHC concentrations. Plasma, bone marrow, and liver alpha-CEHC were significantly increased with H-AT diet compared with other diet groups. In addition, plasma gamma-CEHC was significantly increased in H-T3 rats compared with other diet groups. Nonetheless, there were no effects on bone parameters. Notably, others have found beneficial effects of tocotrienols on bone, as reviewed, 30 but these comparisons were relative to osteopenic rats; our controls had normal bones.
The present study has several limitations. The study focused on bone accretion in slowly growing male rats using single doses of A-AT, H-AT, or H-T3. We chose growing rats, in part, because negative effects of high levels of vitamin E on bone were not apparent in skeletally mature animals. 21 We performed the present study in one gender because evidence is lacking for a major sex difference in skeletal response to vitamin E. However, the absence of sex differences should be confirmed in future studies using multiple doses of vitamin E. Although the skeletons of rats and humans share many similarities, an important limitation of rodents is the absence of intracortical bone remodeling. As such, rodents are generally considered to be most appropriate as models for bone growth and cancellous bone turnover. Consequently, the effect of high vitamin E intake on intracortical bone needs to be evaluated in an animal model with intracortical bone remodeling.
In summary, we demonstrate that long-term intakes of high-dietary alpha-tocopherol or mixed tocols have no effect on bone in slowly growing male rats. This conclusion is supported by multiple studies in the literature, as described, in rodents and humans, which reported no adverse skeletal effects from high vitamin E intakes. We conclude that high intakes of vitamin E do not have an impact on the rat skeleton.
Footnotes
Acknowledgments
This research was supported, in part, by funds from the Helen P. Rumbel endowment to the Linus Pauling Institute, OSU. Alpha-tocopherol was a gift from Archer Daniels Midland. The 50% Tocomin was a gift from Carotech.
Author Discloure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.