
Abstract
Vitexin is a C-glycosylated flavone found in various medicinal plants with several proven biological properties such as anti-inflammatory, antispasmodic, antimicrobial, cytotoxic, and antioxidant activities. Considering that the human population consumes many foods that contain this flavonoid, the objective of this study was to investigate the genotoxic and the antigenotoxic potentials of vitexin by the SMART (Somatic Mutation and Recombination Test) in Drosophila melanogaster wings, in vivo. To evaluate the genotoxic activity, larvae obtained from standard (ST) and high bioactivation (HB) crosses were chronically treated with different concentrations of vitexin (0.15, 0.3, and 0.6 mM). For the evaluation of antigenotoxic activity, the same vitexin concentrations were associated with two damage inducing agents: doxorubicin (0.2 mM) and benzo[a]pyrene (1.0 mM). The results obtained for genotoxic activity showed that vitexin did not induce a statistically significant increment in the frequency of mutant spots, when compared to control. The results for the evaluation of antigenotoxicity indicated that the flavonoid statistically reduced the frequency of mutant spots, when compared to those treated with only the damage inducing agents. Thus, this article presents results that have demonstrated the antigenotoxic activity of vitexin, which could be applied in new studies for the development of drugs with chemoprotective effects.
Introduction
P
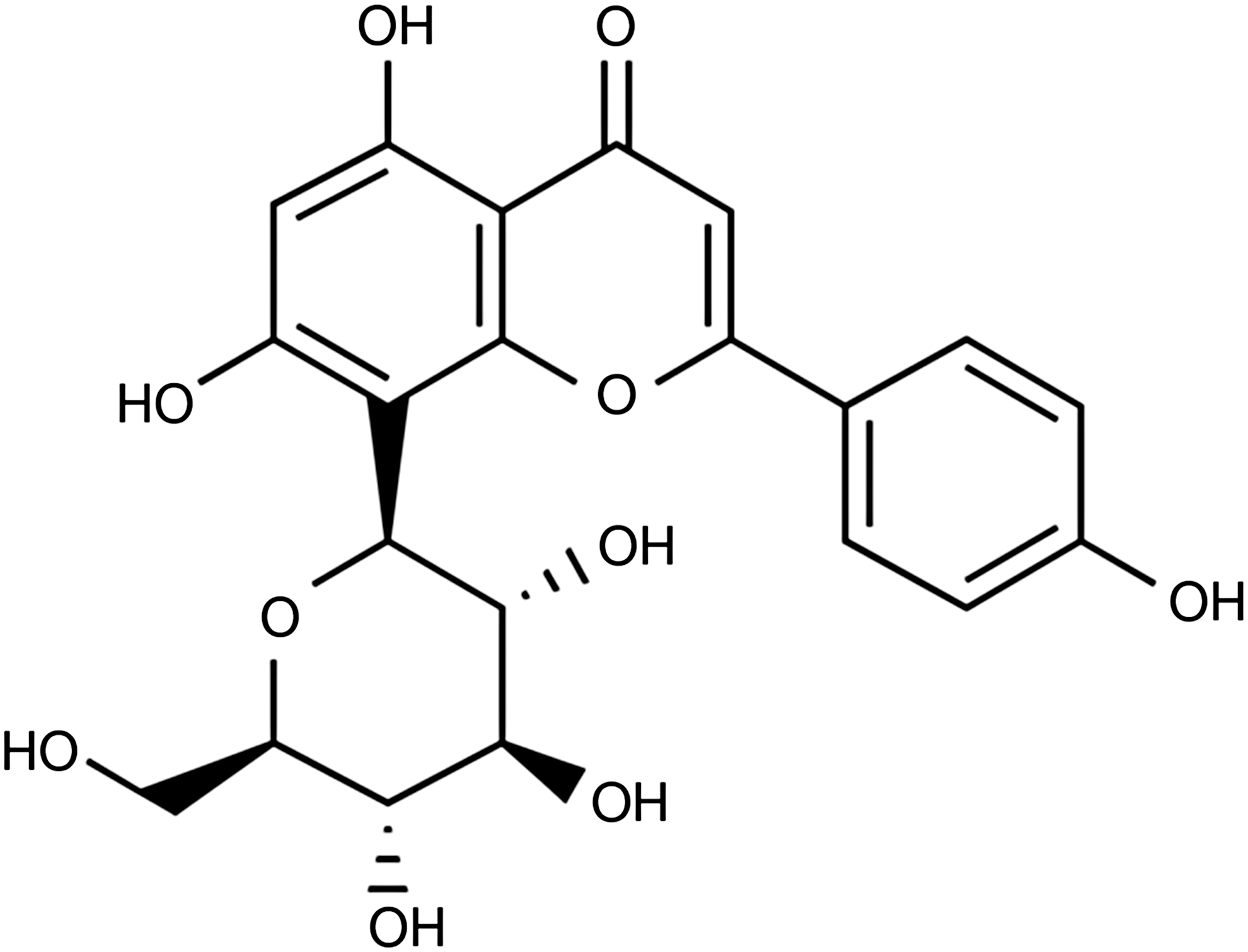
Chemical structure of flavonoid vitexin. 5
Several tests are used to evaluate the genotoxic and antigenotoxic activities of chemical compounds and natural products, including the Somatic Mutation and Recombination Test—SMART. This assay has the advantage of using Drosophila melanogaster as a model organism, which possesses a short time life cycle (about 10 days at 22°C); well-defined morphological characters; an enzymatic system similar to that of mammals, which allows for the metabolism of xenobiotic agents; and a large number of mutant strains, which are genetically well characterized. 11 The SMART is an inexpensive test that produces reliable, unambiguous, and reproducible results. This assay detects a wide spectrum of genetic modifications, including point mutation, deletion, mitotic recombination, and mitotic nondisjunction, 12 although the test is more sensitive for the detection of recombination events. 13
Considering the biological applications of vitexin and the absence of studies on the genotoxic and antigenotoxic potentials of this flavonoid, this study was developed to investigate these activities by using the SMART assay in D. melanogaster wing cells, in vivo.
Materials and Methods
Chemicals
Vitexin (CAS 49513; Sigma) was dissolved at the time of use (20 mL of Milli-Q water, 1% Tween-80, Fluka, and 3% ethanol; Synth). Isomer conversion of vitexin into isovitexin was not considered because its biological activities are similar to that of vitexin, except for the antioxidant activity. Doxorubicin (DXR, CAS 23214-92-8-Biorrub®—Biosintética Ltda), a topoisomerase inhibitor agent, and the DNA-adduct forming agent Benzo[a]pyrene (BaP, CAS B1760; Sigma) were all used as genotoxic agents.
Somatic Mutation and Recombination Test
D. melanogaster strains were kindly provided by and cultivated at the Laboratory of Cytogenetics and Mutagenesis of the State University of Mato Grosso do Sul—Mundo Novo Unit.
The SMART assay was performed through experimental crossings by using three strains containing recessive markers, located in chromosome 3 of the wing imaginal disc of D. melanogaster cells, the multiple wing hairs (mwh) marker (3–0.3), and the flare (flr3 , 3–38.8) marker; [1] a strain of mwh with a y genetic constitution: mwhjv; [2] a strain of “flare-3” (flr3 ), with flr3/In(3LR)TM3ri pp sep l(3)89Aa bx34e and a BdS genetic constitution; [3] a strain of “ORR; flare-3” (ORR; flare-3), with ORR;flr3/In(3LR)TM3, ri pp sep l(3)89Aa bx34e and a BdS genetic constitution. This last strain had chromosomes 1 and 2 from the Oregon R (R) lineage (DDT resistant), containing genes that are responsible for the high level of metabolizing enzymes of the cytochrome P(CYP)6 A2 type. 12 Two different crossings were performed among these strains: (1) ST—a standard cross between “mwh” males and “flr3” virgin females; (2) HB—a high bioactivation cross between “mwh” males and “ORR; flr3” virgin females. 12 Eggs from both crossings were collected over an 8-h period in culture flasks containing a solid agar–agar base (0.04 g/mL), covered by a layer of biological yeast supplemented with sugar. Third instar larvae (72 ± 4 h) were washed with tap water and collected by the use of a thin threaded sieve. The larvae groups were transferred to glass vials containing an alternative medium (instant mashed potato flakes), 1.5 g treated with three concentrations of vitexin (0.15, 0.3, and 0.6 mM). These choices were based on preliminary studies that indicated that concentrations higher than 0.6 mM were toxic to D. melanogaster (IC50 ≥ 260 μg/mL). The concentrations used presented a survival rate higher than 80%. Positive control was prepared with DXR (0.2 mM) and BaP (1 mM); negative control received Milli-Q water, Tween-80 (1%), and ethanol (3%).
The emerging adults carried two kinds of genotypes: mwh +/+ flr3 or mwh+/+ TM3, Bds, and were collected and fixed in 70% ethanol. The wings were mounted on slides in Faure's solution (30 g of Arabic gum, 50 g of chloral hydrate, 20 mL of glycerol, and 50 mL of H2O) and examined for the occurrence of different types of mutant spots, under a light microscope with 400 × magnification.
Statistical analysis
Data were tabulated by the type and size of the mutant spots and subsequently received a statistical treatment according to the methodology described by Frei and Würgler, 14 who used the two-tailed chi-square test for proportions, with a significance level α = β = 0.05. The statistical diagnosis was (+) positive; (−) negative; and (i) inconclusive.
The frequency of each type of mutant spot per individual of a treated series was compared to respective controls, by using the conditional binomial test of Kastenbaum and Bowman. 15 To evaluate the negative results, the multiplication factor (m) was introduced into the test. It was considered to be as follows: m = 2, for simple small spots and total spots due to their high spontaneous frequencies; and m = 5, for large single and twin spots that rarely arose spontaneously. 14 Thus, a criterion was established for a positive diagnosis, which required a frequency of mutations in the treated group that was m times higher than the frequency was obtained in the control group. 16
The calculations of the percentages of inhibition (% I) were carried out from the frequencies of the mutations that were obtained from the individuals who were treated with the samples that were associated with DXR or BaP, according to Abraham 17 : % I = (the frequency of the mutations of DXR or BaP) − (the frequency of the mutations of the samples associated with DXR or BaP) × 100/the frequency of the mutations of DXR or BaP.
Based on the clone induction frequency per 105 cells, the percentages of recombination (% R) were calculated as a mutation frequency: (FM) = the frequency of the clones in the individuals of BH/the frequency of the clones that were obtained in the marked descendant (MH) individuals. The recombination frequency was (RF) = 1- FM. 18
Results
Genotoxic activity of vitexin
Table 1 shows the results that were obtained for the descendants of ST and HB who were treated with vitexin, and which indicated no genotoxic activity; that is, this did not induce a statistically significant increment in mutant spot frequency when compared with the control group (P ≤ .05).
D: Statistical diagnoses according to Frei and Würgler 14 : +, positive; −, negative; i, inconclusive; m, multiplication factor for the assessment of significantly negative results. Significance levels α = β = 0.05.
Including rare flr3 single spots.
MSP, small simple spots, when up to two affected cells with up to three cells away.
MSG, large simple spots, when more than two affected cells with up to three cells away.
MG, twin spots, flr3 and mwh cells observed simultaneously.
TM, total spots.
DXR, doxorubicin; flr3 , flare-3; mwh, multiple wing hairs.
In the ST cross, the total frequency of spots per individual that were observed in the control group was 0.28, whereas treatments with vitexin presented a total frequency of spots that ranged from 0.23 to 0.48. For the HB cross, the total frequency of spots per individual that were observed in the control group was 0.30, whereas treatments with vitexin presented a total frequency of spots that ranged from 0.08 to 0.20.
Antigenotoxic activity of vitexin that was associated with the genotoxic agent BaP
Before evaluating the antigenotoxic activity of vitexin that was associated with BaP, a pilot test was conducted to determine the concentrations of BaP to be used, as there were no reports in the literature with this compound for the SMART. It was determined that a concentration of 1 mM BaP was significantly genotoxic, so it could be associated with the flavonoid vitexin. These results are shown in Tables 2 and 3. For the ST, HB, and trans-heterozygous MH, treated with BaP, the results indicated a statistically significant difference in the total frequency of mutant spots when compared to the control group (P ≤ .05).
Diagnostic and Statistical according Br and Würgler (1988): +, positive; f +, weakly positive; −, negative; i, inconclusive. m, the multiplication factor for the evaluation of significantly negative. Significance level α = β = 0.05.
Including simple flr3 spots.
Considering the mwh clones for simple mwh stains and the twin spots.
Just simple mwh spots can be seen in subjects heterozygous mwh/TM3, since the chromosome balancer TM3 does not contain the mutant gene flr3 .
Induction frequencies corrected for spontaneous incidence estimated from negative controls.
Calculated according to Abraham. 17
BaP, benzo[a]pyrene.
Diagnostic and Statistical according Br and Würgler (1988): +, positive; f +, weakly positive; −, negative; i, inconclusive. m, the multiplication factor for the evaluation of significantly negative. Significance level α = β = 0.05.
Including simple flr3 spots.
Considering the mwh clones for simple mwh stains and the twin spots.
Just simple mwh spots can be seen in subjects heterozygous mwh/TM3, since the chromosome balancer TM3 does not contain the mutant gene flr3 .
Induction frequencies corrected for spontaneous incidence estimated from negative controls.
Calculated according to Abraham. 17
For the ST descendants, the total frequency of spots per individual that were observed in the control group was 0.28, whereas those treated with BaP showed an overall spot frequency of 0.68. For the HB cross, the total frequency of spots per individual that were observed in the control group was 0.30, while those treated with BaP showed an overall spot frequency of 1.58.
The results of the analyses of ST, HB, and the trans-heterozygous MH, treated with vitexin associated with BaP, indicated a reduction in the overall frequency of spots per individual, at all of the concentrations tested, when compared to the treatments that only had BaP. In the ST cross, the total frequency of spots per individual that were observed in the treatments that only had BaP was 0.68, whereas in the treatments of vitexin associated with BaP showed a total frequency of spots ranging from 0.35 to 0.05. For the HB cross, the total frequency of spots per individual that were observed in the treatments that only had BaP was 1.58, whereas in the groups treated with vitexin that were associated with BaP ranged from 1.00 to 0.90. Thus, vitexin protected the DNA of the BaP-induced damage effect and presented an antigenotoxic activity in the SMART.
In the ST cross, the concentrations of 0.15, 0.3, and 0.6 mM of vitexin that were associated with BaP inhibited 81.7%, 81.7%, and 93.9% of the genotoxic events caused by BaP, respectively (Table 2). In the HB cross, the percentages of inhibition ranged 45.2%, 45.2%, and 52.8%, respectively, for the same concentrations (Table 3).
From the results observed in the ST, HB, and trans-heterozygous MH that had been treated with vitexin associated with BaP, the heterozygote balanced descendants were analyzed to determine the frequency of recombination. In the ST cross, cotreated with BaP and vitexin, the frequency of clone formation by cell division ranged from 0.71, 0.71, and 0.61 × 10−5, respectively, to concentrations of 0.15, 0.3, and 0.6 mM of vitexin, while the frequency that was observed for BaP was 1.38 × 10−5 (Table 2). For the descendants of HB, cotreated with vitexin and BaP, the frequency of cell division by the clones ranged from 2.04, 2.04, and 1.84 × 10−5, respectively, to concentrations of 0.15, 0.3, and 0.6 mM of vitexin, while those observed for BaP were 3.22 × 10−5 (Table 3).
The percentages of recombination showed that BaP caused 93.8% of recombination in the ST cross and 93.1% of recombination in the HB cross. Vitexin treatments that were associated with BaP caused 100% of a recombination in the ST cross and 100%, 100%, and 78.8%, respectively, of a recombination in the HB cross.
Comparing the frequencies of clone formation by cell division, the trans-heterozygous MH, when compared with the frequencies obtained in the heterozygous balanced descendants of the ST cross and the HB cross, simultaneously treated with vitexin and BaP, showed a predominantly recombinogenic activity (Tables 2 and 3).
Antigenotoxic activity of vitexin that was associated with the genotoxic agent DXR
Considering the negative results that were observed in the genotoxicity test, vitexin was also evaluated for its antigenotoxic activity when associated with the genotoxic agent DXR (0.2 mM).
The results for the evaluation of the antigenotoxicity of vitexin when associated with DXR are listed in Tables 4 and 5. For the ST, HB, and the trans-heterozygous MH treated with DXR, the results showed a statistically significant increment in the total frequency of mutant spots, when compared to the control group (P ≤ .05).
Diagnostic and Statistical according Br and Würgler (1988): +, positive; f +, weakly positive; −, Negative; i, inconclusive. m, the multiplication factor for the evaluation of significantly negative. Significance level α = β = 0.05.
Including simple flr3 spots.
Considering the mwh clones for simple mwh stains and the twin spots.
Just simple mwh spots can be seen in subjects heterozygous mwh/TM3, since the chromosome balancer TM3 does not contain the mutant gene flr3 .
Induction frequencies corrected for spontaneous incidence estimated from negative controls.
Calculated according to Abraham. 17
Diagnostic and Statistical according Br and Würgler (1988): +, positive; f +, weakly positive; −, negative; i, inconclusive. m, the multiplication factor for the evaluation of significantly negative. Significance level α = β = 0.05.
Including simple flr3 spots.
Considering the mwh clones for simple mwh stains and the twin spots.
Just simple mwh spots can be seen in subjects heterozygous mwh/TM3, since the chromosome balancer TM3 does not contain the mutant gene flr3 .
Induction frequencies corrected for spontaneous incidence estimated from negative controls.
Calculated according to Abraham. 17
In the ST cross, the total frequency of spots per individual that were observed in the control group was 0.28, whereas those treated with DXR showed a total frequency of 10.33 spots. In the HB cross, the total frequency of spots per individual that were observed in the control group was 0.30, while those treated with the DXR showed a total frequency of 17.90 spots.
The results of the analyses of the ST, HB, and the trans-heterozygous MH treated with vitexin associated with DXR indicated a reduction in the total frequency of spots per individual in all of the concentrations tested when compared to the treatments that only had DXR. In the ST cross, the total frequency of spots per individual that were observed in the DXR treatments was 10.33, while in the treatments with DXR that were associated with vitexin showed an overall frequency of spots ranging from 4.10 to 1.78. In the HB cross, the total frequency of spots per individual that were observed in the treatments that only had DXR was 17.90, whereas in the groups treated with DXR that were associated with vitexin ranged from 6.50 to 1.70. Thus, vitexin protected the DNA of the DXR action and presented an antigenotoxic activity in the SMART.
The concentrations of 0.15, 0.3, and 0.6 mM of vitexin that were associated with DXR inhibited 61.9%, 79.3%, and 85.09%, respectively, regarding the genotoxic events caused by DXR in the ST cross. In the HB cross, the percentages of inhibition were 64.79%, 92.04%, and 88.91%, respectively (Table 4).
Given the results that were observed with the ST, HB, and trans-heterozygous MH treated with vitexin associated with DXR, an analysis was performed of the heterozygote balanced descendants to determine the frequency of recombination. In the ST cross, cotreated with DXR and vitexin, the frequency of the cell division clone formation was 8.40, 4.81, and 3.63 × 10−5, respectively, for concentrations of 0.15, 0.3, and 0.6 mM of vitexin, whereas with DXR, the frequency observed was 21.15 × 10−5 (Table 5). For the HB descendants cotreated with vitexin and DXR, the frequency of clone formation per cell division was 13.31, 3.48, and 4.61 × 10−5, respectively, for concentrations of 0.15, 0.3, and 0.6 mM of vitexin, whereas with DXR, the frequency observed was 36.68 × 10−5 (Table 5).
The percentages of recombination showed that DXR resulted in 100% of a recombination in the ST cross and for the treatment of vitexin that was associated with DXR, the percentages of recombination were 95.41%, 84.47%, and 100%, respectively. For the HB descendants, DXR resulted in 98.42% of recombination and the vitexin treatments that were associated with DXR resulted in 92.76%, 100%, and 100%, respectively (Table 5).
Comparing the frequency of clone formation per cell division of the marked trans-heterozygous descendants, when compared with the heterozygous balanced descendants of the ST cross and the HB cross, simultaneously treated with DXR, showed a predominantly recombinogenic activity.
Discussion
Genotoxicity assays are normally used to identify extracts/substances that have the ability to interact with nucleic acids at low concentrations. When a genotoxic agent interacts with DNA, this interaction may lead to chromosomal aberrations and/or changes in DNA structure that may affect the fidelity of the message and lead to irreversible changes in the cell. 19 The consequences of such DNA impairment could be the establishment of and/or predisposition to diseases, increased morbidity/mortality, changes in heritable characteristics, impaired reproductive capacity, and cancer development. 20 In the present study, we have investigated the genotoxic and antigenotoxic activity of the flavonoid vitexin by the SMART, in vivo. This test is a versatile and efficient tool to detect the genotoxic and antigenotoxic effects of several chemicals, such as plant extracts, which can cause a wide spectrum of genetic abnormalities, mitotic recombination, deletion, point mutation, and chromosome loss. 21,22
In assessing the possible genotoxic actions of vitexin, data indicated that this compound showed no genotoxic activity. It did not cause damage to the DNA in the somatic cells of D. melanogaster wings at the tested concentrations.
There is no literature information about the genotoxic activity of the flavonoid vitexin, but other flavonoids have been evaluated by the SMART, for example, kaempferol, quercetin, and quercetin 3-D-glucoside. The data obtained therein showed that at the concentrations evaluated, these compounds did not induce somatic mutations or recombination in D. melanogaster wings, with an exception being made for quercetin, which was genotoxic at 50 μM. 23 This information suggests that flavonoids are, in general, a nongenotoxic group of substances.
Vitexin that was isolated from the fruits and leaves of the plant Euodia danielli showed no cytotoxic activity on different human tumor cell lines (A549, Skov-3, SKMEL-2, XF, and HTC15). 24 However, vitexin that was isolated from Acer palmatum decreased cell viability, suppressed tumor growth, and induced apoptosis in the OC2 tumor cell line. 25 According to Papi et al., 10 vitexin showed no cytotoxic activity for the CaCo, Lovo, MCF-7, and MDA-MB-231 cell lines, nevertheless, it induced the apoptosis pathway of these cells.
Vitexin was capable of protecting the DNA against BaP genotoxic action and thus presented an antigenotoxic activity in the SMART. An analysis of the results indicated that vitexin, when it was associated with BaP, reduced the frequency of mutant spots in the descendants of the ST and HB crosses. A reduction in the frequency of mutant spots was observed in all of the concentrations, when compared with the results of BaP.
For the HB descendants, the reduction in the frequency of spots was higher than those for the ST cross. These differences in the responses that were observed are due to the high levels of CYP-450, xenobiotic metabolizing enzymes, which make these individuals more sensitive to detect promutagens and procarcinogens. 12 This is also due to the BaP mechanism of action that needs to be metabolized by this enzyme complex to become a genotoxic agent, promoting cell cycle disturbances, chromosome instability, chromosomal aberrations, DNA damage, and apoptosis, which could be related to cancer development. 26,27
Regarding the percentages of inhibition and recombination when vitexin was associated with BaP, there was an inhibition in the frequency of mutant spots that were predominantly a recombinogenic activity in the ST and HB descendants.
For the antigenotoxic activity, it was observed that vitexin protected the DNA from the DXR actions and hence presented an antigenotoxic activity, probably by scavenging free radicals generated by the DXR. 28,29
The results have shown that the flavonoid vitexin that was associated with DXR reduced the frequency of mutant spots in the ST and HB descendants. It was observed that there was a reduction in the frequency of mutant spots for all of the tested concentrations of vitexin and thus demonstrating its antigenotoxic activity.
Regarding the percentages of inhibition and recombination, DXR inhibited the frequency of mutant spots in ST and HB descendants, and a predominantly recombinogenic activity was observed. According to Spencer et al., 30 this result is likely due to the fact that the homologous recombination is the main pathway used in the repair of DNA double-strand breaks generated by DXR.
By analyzing the frequency of clone formation per cell division, in the trans- heterozygous MH, as obtained in the heterozygous balanced descendants of the ST and HB cross, simultaneously treated with vitexin and DXR, there was also a recombinogenic activity. The observed results have suggested that under these experimental conditions, vitexin has an antigenotoxic effect, probably due to scavenging free radicals generated by the DXR, but it did not inhibit the recombinogenic activity of DXR.
These results can be attributed to the antioxidant capacity of vitexin, as flavonoids act as antioxidant agents by several mechanisms, including a free radical scavenging and a metal chelation, as well as the mediation of enzyme inhibition. Furthermore, they are extremely safe, with a low toxicity, making them excellent candidates for chemopreventive agents. 31,32
Vitexin and isovitexin, isolated from Ficus deltoidea leaves, exert a neuroprotective effect on SH-SY5Y neuroblastoma cells, which strongly correlates with its antioxidant capacity. 33 In general, flavonoids inhibit the bioactivation of carcinogens by repressing the pre-biotransformation of several enzymes or by blocking membrane receptors that are involved in the uptake of precarcinogens. 34
The results of this study have indicated that the flavonoid vitexin, as found in several medicinal plants and herbal medicines, showed no genotoxic activity. It did not increase the frequency of mutant spots when using the SMART on D. melanogaster. In addition, this compound showed an antigenotoxic activity by significantly reducing the frequency of mutant spots (small simple spots, large simple spots, and twin spots) that were induced by the genotoxic agents BaP and DXR. This was probably due to the antioxidant activity exerted by the flavonoid. Thus, this article has presented results that have demonstrated the antigenotoxic activity of vitexin and its possible action to be applied in new studies for the development of potential chemopreventive medicines.
Footnotes
Acknowledgment
The authors thank the Coordination of Improvement of Higher Level Personnel (CAPES), Brazil, for the scholarship support.
Author Disclosure Statement
No competing financial interests exist.