
Abstract
Wine contains various polyphenols such as flavonoids, anthocyanins, and tannins. These molecules are responsible for the quality of wines, influencing their astringency, bitterness, and color and they are considered to have antioxidant activity. Polyphenols, extracted from grapes during the processes of vinification, could protect the body cells against reactive oxygen species level increase and could be useful to rescue several pathologies where oxidative stress represents the main cause. For that, in this study, red and white wine, provided by an Italian vinery (Marche region), have been analyzed. Chromatographic and morphofunctional analyses have been carried out for polyphenol extraction and to evaluate their protective effect on human myeloid U937 cells exposed to hydrogen peroxide. Both types of wines contained a mix of phenolic compounds with antioxidant properties and their content decreased, as expected, in white wine. Ultrastructural observations evidenced that wines, in particular red wine, strongly prevent mitochondrial damage and apoptotic cell death. In conclusion, the considered extracts show a relevant polyphenol content with strong antioxidant properties and abilities to prevent apoptosis. These findings suggest, for these compounds, a potential role in all pathological conditions where the body antioxidant system is overwhelmed.
Introduction
P
In this study, we investigated the contribution of phenolic compounds in Italian wines obtained from grapes grown in the Marche region. Therefore, the aim of this study was, first of all, to characterize red and white wine phenolic composition and their antioxidant power by means of chemical analyses. Then, wine extract effects against oxidative stress and cell death have been studied, through morphofunctional approaches, in a hematopoietic cell line (U937) exposed to a pro-oxidant injury in vitro. 19
Materials and Methods
Wines from vinery
The wines chosen are Grosso Agontano (Conero Doc Riserva, red old wine), Guasco (Rosso Conero Doc, red young wine), and Serra Fiorese (Verdicchio dei Castelli di Jesi Doc, Classico Riserva, white wine).
Sample preparation for chemical assays
Two hundred milliliters of wine samples have been diluted with 200 mL of distilled water and then extracted with ethyl acetate (60 mL × three times). The ethyl acetate extracts have been pooled together and concentrated under reduced pressure at 40°C until small volume (3–5 mL) and diluted to 10 mL with methanol.
2,2-Diphenyl-1-picrylhydrazyl radical assay
One milliliter of wine sample (appropriately diluted with citrate buffer/methanol to keep the absorbance in the range of 0.200–0.800 AU) has been added to a solution of 1 mL 2,2-diphenyl-1-picrylhydrazyl radical (DPPH) methanolic solution (0.12 mg/mL) and 1 mL of citrate buffer (pH = 3.0, 10 mM). The absorbance of the reaction mixture has been measured at 517 nm after 20 min. Calibration has been performed using caffeic acid as standard, in the concentration range of 0.150–0.275 mg/mL, following the same procedure. The calibration curve has been used to calculate antioxidant activity. The determination has been carried out in triplicate.
High-performance liquid chromatography (HPLC)-DAD-mass spectrometry analysis
Separation has been accomplished using a Waters 2795 Alliance HPLC System and a Waters 2996 Photodiode Array Detector. The photodiode array detector (DAD) has been coupled to electrospray ionization (ESI) in both positive and negative ion modes in a Micromass ZQ mass spectrophotometer (Micromass Ltd.). Nitrogen gas has been used as a nebulizing and drying gas, and different fragmenting voltages have been applied. For data acquisition, MassLynx V4.1 software (Micromass Ltd.) has been used. The samples have been analyzed on reverse phase column (LiChroCART 250-4 Purospher Star RP18, 5 μm as column, and LiChroCART 4-4 Purospher Star RP18, 5 μm as guard column; Merck) at a constant temperature (30°C). The injection volume was 20 μL. The mobile phase flow rate was 0.8 mL/min.
The solvents used were acetonitrile (A) and 0.1% formic acid in water (B), with the following mobile phase gradient for phase A: isocratic elution 5% for 2 min, linear gradient elution from 5% to 15% for 2 min, to 30% for 8 min, to 60% for 10 min, and maintained at 60% for 3 min more. The total acquisition time was 25 min. UV/Vis spectra were recorded from 210 to 400 nm, using a diode array detector. The mass spectrometry (MS) detector operated at a capillary voltage of 3.5 kV, a cone voltage of 30 V, an extractor voltage of 1 V, a source temperature of 100°C, a desolvation temperature of 300°C, a desolvation gas flow of 500 L/h, and cone gas flow of 50 L/h. Full-scan spectra were recorded over the range m/z 100–350 amu. The identification of polyphenols has been accomplished by mass and UV-Vis spectrometry and literature data. 20
Cell culture and apoptosis induction
U937 Human myelomonocytic cell line was grown in RPMI 1640, supplemented with 10% heat-inactivated fetal bovine serum, 2 mm glutamine, 1% antibiotics, and was maintained at 37°C in humidified air with 5% CO2.
For apoptosis induction, U937 cells (seeded at 1 × 106 cells/mL) have been exposed to 0.5 mM H2O2 for 24 h 19 and cell behavior has been checked by means of inverted microscopy (IM). Cell viability has been monitored through trypan blue (TB) exclusion assay.
Antioxidant treatments
Different doses of extracts have been evaluated on cell viability to find a no toxic concentration for cells (10-2 μg/mL for Grosso Agontano; 0.004–0.001 μg/mL for Guasco; and 0.002–0.0004 μg/mL for Serra Fiorese). Then, the extract doses have been tested against H2O2 damage to identify the best extract concentration. Therefore, cells were pretreated with 2 μg/mL Grosso Agontano, 0.001 μg/mL Guasco, and 0.0004 μg/mL Sierra Fiorese for 24 h before and after the apoptosis induction.
Cell viability monitoring with or without antioxidant administration
TB exclusion assay 21 has been employed to monitor possible polyphenol cytotoxic effect, to choose the optimal extract dose, and to evaluate if antioxidant administration, before apoptosis induction, improves cell viability.
Acridine orange and propidium iodide nuclei staining
Acridine orange (AO) and propidium iodide (PI) are fluorescent nuclear probes useful to detect viable, apoptotic, and necrotic cells. In living cells, AO diffuses through the plasma membrane intercalating into DNA and emitting green fluorescence. Cells show bright green nuclei when undergoing early apoptosis. Late apoptotic cells are stained by both AO and PI (orange nuclei), while the necrotic ones by PI alone (red nuclei). 2,19
Cells were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) pH 7.4 for 30 min. Then, they were deposited on polylysinated coverslips in Petri dishes. After two washing steps in PBS, cells were pretreated with RNasi A 10 μg/mL in PBS for 30 min and then exposed to an equal mixture of PI (1 μg/mL; Life Technologies) and AO (1 μg/mL; Life Technologies) diluted in PBS at room temperature in the dark for 10 min. 19 Samples have been observed with a Leica TCS-SP5 Confocal Laser Scanning Microscope (CLSM) connected to a DMI 6000 CS Inverted Microscope (Leica Microsystems CMS GmbH; AO and PI excitation were at 488 and 500 nm, respectively, and their emission signals were detected at 617 and 525).
Transmission electron microscopy
Pellets have been immediately fixed in 2.5% glutaraldehyde in 0.1 M in phosphate buffer (pH 7.3). Then, samples have been postfixed in 1% OsO4 in the same buffer and dehydrated with ethanol embedded in araldite, as previously described. 1 Thin sections, collected on nickel grids and stained with uranyl acetate and lead citrate, have been observed with a CM10 electron microscope (FEI Italia SRL, Milano, Italy).
Mitochondrial behavior at CLSM
Mitochondrial viability has been evaluated by means of CLSM. In particular, mitochondrial behavior has been monitored through 10-N-nonyl acridine orange assay (NAO, to evaluate cardiolipin peroxidation events, thus monitoring lipid changes due to oxidative stress increase) and mitochondrial transmembrane potential performed through JC-1 probe (to evaluate organelle functionality). Cells have been incubated with 50 nM NAO for 10 min at room temperature or 2 μg/mL JC-1 for 20 min at 37°C.
Samples have been observed, directly on coverslip, with a Leica TCS-SP5 CLSM connected to a DMI 6000 CS Inverted Microscope (Leica Microsystems CMS GmbH; NAO excitation was at 504 nm and emission at 653 nm; JC-1: red fluorescence intensity [excitation, 562 nm; emission, 608 nm] and green fluorescence intensity [excitation, 503 nm; emission, 547 nm]).
Statistical analyses
All data are presented as mean ± standard deviation. Student's t-test has been applied in statistical analyses to compare results. *P < .05 and **P < .005 have been considered as the significance threshold.
Results
Chemical analyses
The antioxidant activities of extracts have been evaluated by DPPH assay and the results have been reported in Table 1. The extracts have been also analyzed by HPLC-DAD-MS (ESI) and the main detected compounds have been reported in Table 2. Cromatographic analysis evidenced some differences in the wine composition. In particular, as expected, Grosso Agontano exhibited the major content in polyphenols, that is, gallic acid, catechin, ethyl gallate, myricetin, and Quercetin. The other red wine, Guasco, showed similar peaks to Grosso, but lower in concentration. Serra Fiorese exhibited a low content of polyphenols and the major peaks were associated with catechin, epicatechin, caffeic, and coumaric acid (Table 2).
Antioxidant activities of wine extracts determined by DPPH assay.
CAEAC, caffeic acid equivalent antioxidant capacity; DPPH, 2,2-diphenyl-1-picrylhydrazyl radical.
Identification and quantification of the major constituents in wine extracts by HPLC-DAD-MS analysis.
Characteristic wave length.
The concentration of the compound was determined at 280 nm using Catechin as external standard and it has been expressed in μg/mL.
The concentration of the compound has been determined at 320 nm using caffeic acid as external standard and it has been expressed in μg/mL.
The concentration of the compound has been determined at 360 nm using Quercetin as external standard and it has been expressed in μg/mL.
HPLC, high-performance liquid chromatography; MS, mass spectrometry.
Biological assays
Different doses of wine extracts have been tested on cells. TB staining revealed that at the highest concentrations (10 μg/mL for Grosso Agontano, Fig. 1B; 0.004 μg/mL for Guasco, Fig. 1E; 0.002 μg/mL for Serra Fiorese, Fig. 1H), the molecules induced a relevant cell viability decrease (Fig. 1). After a deep dose reduction (4 μg/mL for Grosso Agontano, Fig. 1C; 0.002 μg/mL for Guasco, Fig. 1F; 0.001 μg/mL for Serra Fiorese, Fig. 1I), an improvement of cell viability could be observed and quantified. At lower concentrations (2 μg/mL for Grosso Agontano, Fig. 1D; 0.001 μg/mL for Guasco, Fig. 1G; 0.0004 μg/mL for Serra Fiorese, Fig. 1J), the antioxidants did not influence cell viability. The best doses have been chosen considering the absence of toxic effects on cell culture and the capacity to display a protective action against the oxidant injury. In fact, lower concentrations induced a number of cell death amount comparable with the oxidant treatment, indicating a scarce antioxidant effect (data not shown).
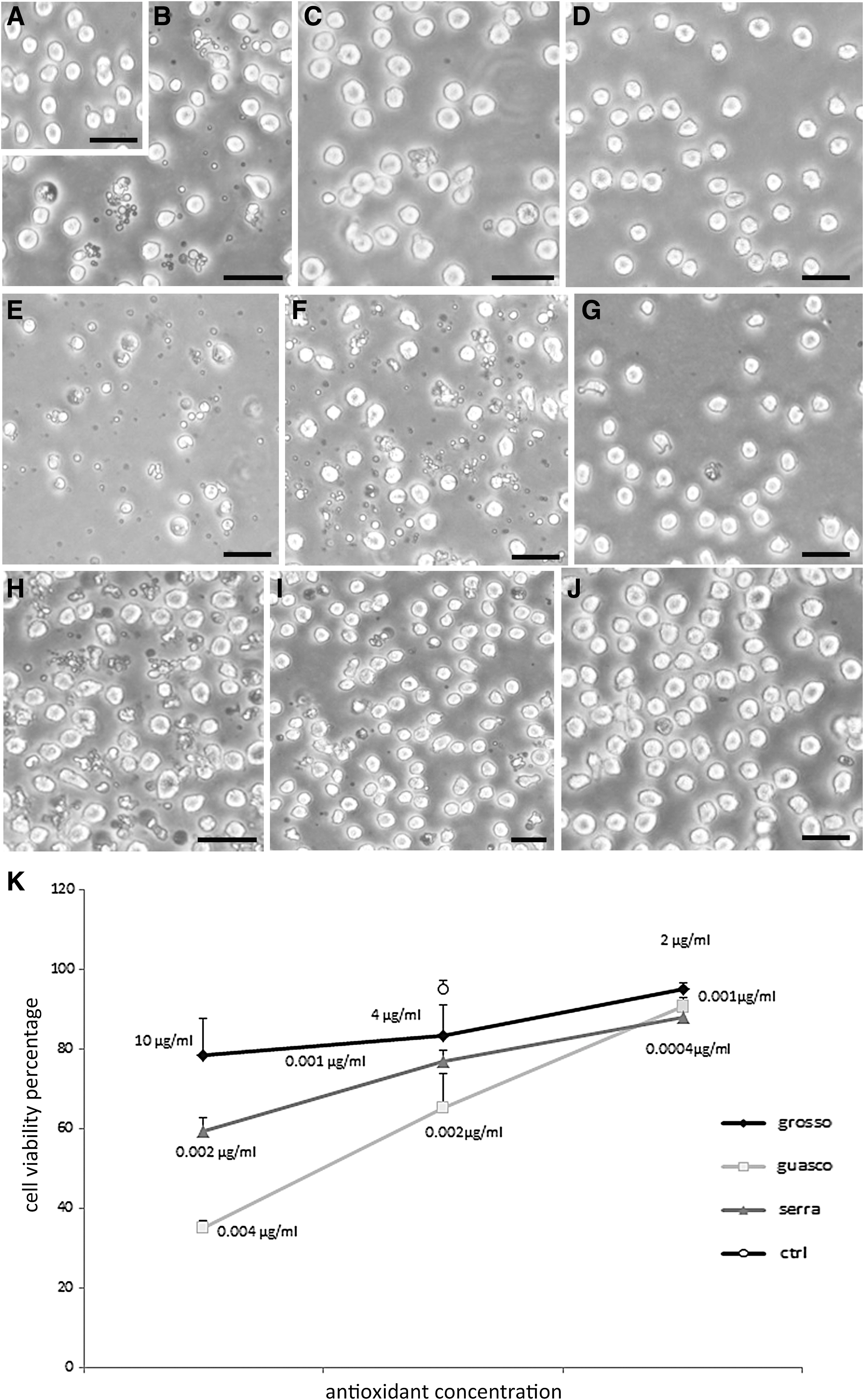
IM micrographs show control
The best antioxidant doses were 2 μg/mL for Grosso Agontano, 0.001 μg/mL, for Guasco, and 0.0004 μg/mL for Sierra Fiorese.
U937 cells have been treated with these antioxidant concentrations, 24 h before 0.5 μM H2O2 exposure, to counteract the oxidant injury.
Figure 2 shows the protective effect of wine compounds against H2O2 at IM. If compared with control condition, where cell viability percentage was about 94.6% (Fig. 2A), the trigger induced a viability decrease to 42.5% as clearly displayed at IM (Fig. 2B). Pretreating samples with polyphenols before oxidation, the number of living cells increased (75.1% for Grosso Agontano, Fig. 2C; 59% for Guasco, Fig. 2D; and 48.5% for Serra Fiorese, Fig. 2E). According to chemical analyses, the antioxidant protection resulted statistically significantly for Grosso Agontano as shown in the cell viability graph (Fig. 2). The extract effects have been evaluated also after oxidative damage induction, showing the absence of protective effect (Fig 2F, G, H). Since H2O2 induces both apoptosis and necrosis, we have investigated polyphenol ability to counteract apoptosis or necrosis using the AO/PI double staining. This analysis showed that all wines, in particular Grosso Agontano, were able to prevent apoptotic cell death. In fact, comparing the number of apoptotic cells treated with H2O2 and that of cells pretreated with extracts before the injury, a reduction of dead cell value could be appreciable. In particular, this is statistically relevant in the case of Grosso (**P < .005). Antioxidants were able to reduce necrotic cell death, but not in a statistically significant manner (graph, Fig. 3).
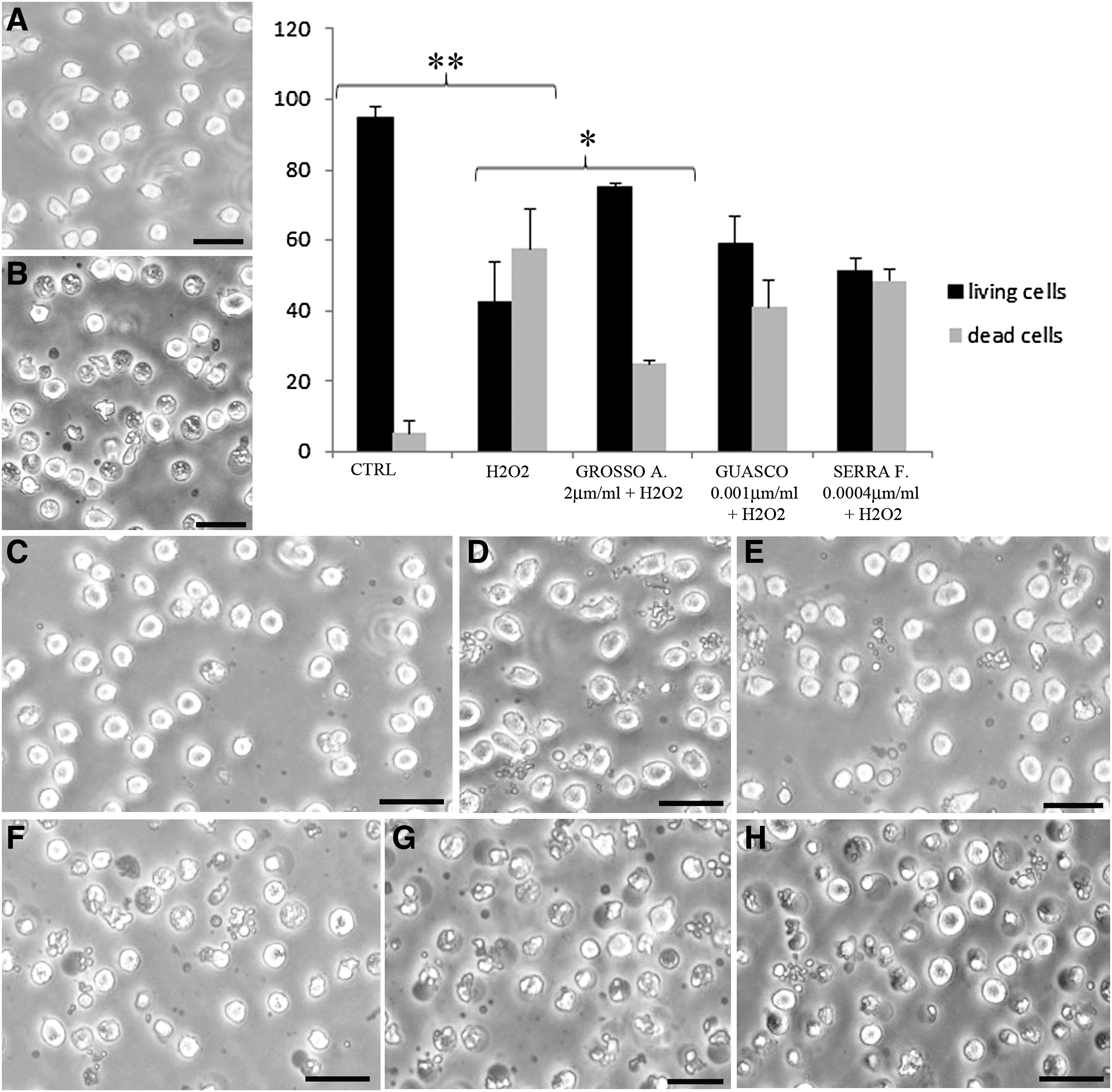
IM micrographs show control
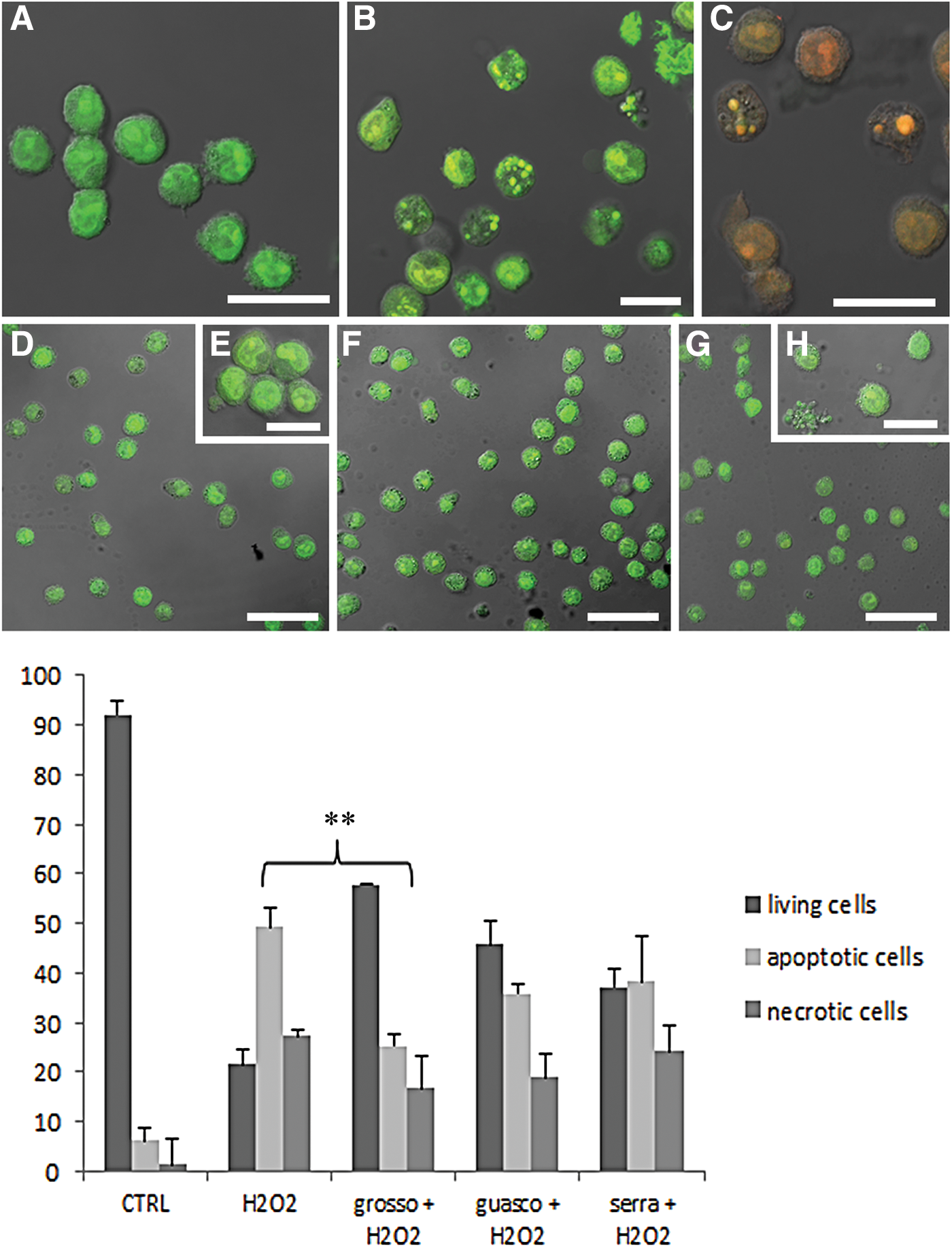
CLSM micrographs of AO/PI nuclear staining in control
In control condition (Fig. 3A), cells showed a good viability with preserved green nuclei. After H2O2 exposure, early apoptotic (green bright fluorescence, Fig. 3B) and late apoptotic or necrotic stages (orange fluorescence, Fig 3C) appeared. These death patterns decreased in polyphenol-treated samples (Fig. 3D–H), where cells showed green fluorescence similar to control condition.
As evidenced in Figure 3, Grosso Agontano administration before pro-oxidant exposure induced a significant decrease of apoptotic cell number and, to a lesser extent, of the necrotic ones. Guasco and Serra Fiorese wines induced only a moderate reduction of cell death and besides that they showed a satisfying effect in delaying apoptosis. This can be due to the fact that Guasco is a young red wine and Serra Fiorese a white wine and for their nature with lower antioxidant content.
Ultrastructural analyses (Fig. 4) confirmed quantitative data. Control condition (Fig. 4A) showed a preserved cell morphology, while after H2O2 administration, apoptosis could be observed with its typical features: chromatin condensation, cup-shaped masses, and apoptotic bodies (Fig. 4D). Some cells in secondary necrosis appeared. In pretreating cells with wine extracts (Fig. 4G, L, O), particularly in the case of Grosso Agontano (Fig. 4G), an improvement of cell condition could be detected.
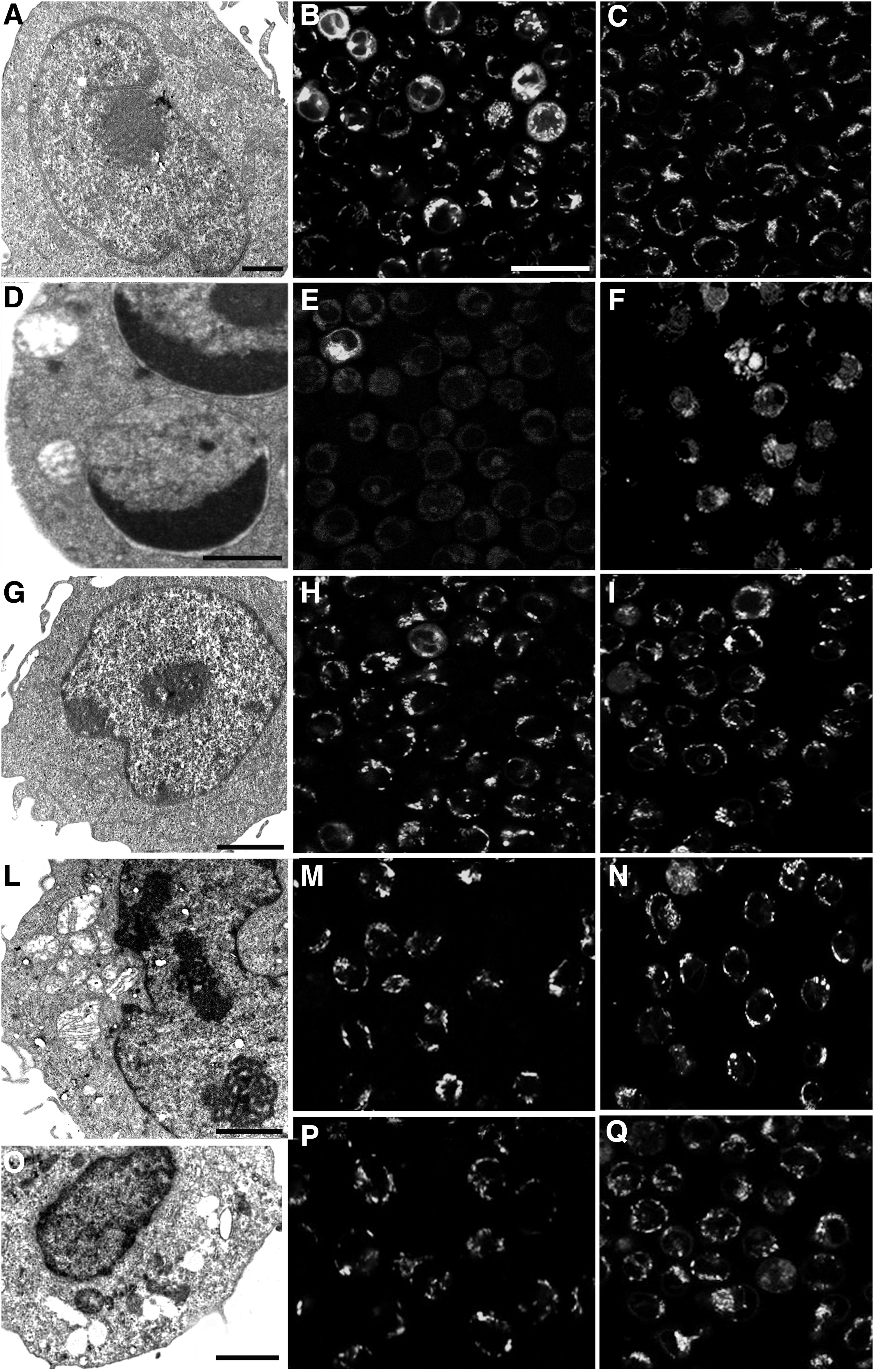
TEM
Moreover, mitochondrial behavior has been monitored at confocal microscopy, which allowed analyzing mitochondrial membrane integrity and its transmembrane potential (Fig. 4). In control samples, high NAO fluorescence intensity appeared (Fig. 4B), suggesting the preserved mitochondrial presence. On the other hand, after H2O2 treatment (Fig. 4E), the fluorescence decreased due to structural cardiolipin degeneration, which led to membrane integrity loss and cytochrome c leak, an event strictly correlated with apoptosis. 21 The number of mitochondria with oxidized cardiolipin decreased after polyphenol pretreatment (Fig. 4H, M, P), which favored the maintenance of their integrity. Mitochondrial transmembrane potential has been scored through JC1 probe revealing a red fluorescence intensity (i.e., a good mitochondrial functionality) in control condition (Fig. 4C), which decreased after oxidative damage, becoming yellow (Fig. 4F). Antioxidant administrated before H2O2 exposure has been able to preserve and improve mitochondrial transmembrane potential in U937 cells (Fig. 4I, N, Q); this is particularly evident after Grosso Agontano treatment (Fig. 4Q).
Discussion
The antioxidant activity of wine is largely dependent on its phenolic content. The proportion of compounds depends on the variety of grape, on the growing region, on the method of production, and on the aging process. Moreover, any variation in the vinification process that introduces a difference in phenolic composition of the wine could influence its antioxidant activity. 15 Wine comprises water, ethanol, glycerol, polysaccharides, different types of acids, and phenolic compounds. These latter can be divided into flavonoids and nonflavonoids. Flavonoids, which account for over 85% of the phenolic components in red wine, include different molecular families such as flavonols (i.e., myricetin, quercetin), flavones, and anthocyanidins. 22 Red wine is known to contain 10-fold more phenolic compounds than white wine. 23 Most of flavonoid beneficial effects 24 –26 are due to their potent antioxidant and free radical scavenging properties, as well as their ability in modulating many cellular enzyme functions. 27 –30
In this study, the antiapoptotic activity of phenolic compounds, extracted from red and white wine, has been demonstrated in a cell model exposed to oxidative damage. As described in the Results section, Grosso Agontano spectrum showed a relevant content in polyphenols with respect to other wines, conferring to this extract a significant ability in preventing apoptosis induced by reactive oxygen species (ROS) increase. In particular, cromatographic analysis revealed that it contained a consistent quantity of gallic acid, catechin, ethyl gallate, myricetin, and quercetin, with scavenger activity against a variety of ROS. 31 –43 Thus, Grosso Agontano ability to counteract H2O2-induced cell death in U937 cell line could be due to the synergic effect of these compounds. Moreover, caffeic and gallic acid 44,45 known for their antioxidant ability, at least in part, could contribute to enhance the antioxidant properties of Grosso Agontano, although their presence is lower than the other compounds.
Guasco beverage showed a minor ability to prevent cell death. It presented the highest antioxidant power (expressed as EAC), but it must be used on cell culture diluted 1000 times if compared with Grosso Agontano concentration. Chromatographic analysis for Guasco content revealed lowest peaks of phenolic compounds except for caffeic acid.
Finally, this work evidenced that the white wine Serra Fiorese, under conditions which promote oxidative stress, was able to maintain the cellular functions, but with a minor exit if compared with red wines. Serra Fiorese showed a spectrum with abundant catechin, epicatechin, caffeic, and coumaric acid presence. The latter, present with a low peak in red wine, has been described as antioxidant agent 46 and could be responsible for Serra Fiorese antioxidant properties.
These findings revealed that vinery products, thanks to the geographic location and to the vinification process, are rich in polyphenol content. In particular, Grosso extract is able to prevent U937 apoptotic cell death, counteracting H2O2-induced oxidative damage, maintaining mitochondrial membrane integrity, and enhancing the recovery of mitochondrial function.
Footnotes
Acknowledgments
The authors are indebted to Garofoli Company for kindly providing red and white wines. The research was supported by Urbino University.
Author Disclosure Statement
No competing financial interests exist.