
Abstract
Glucomannan-enriched squid surimi improves cholesterolemia and liver antioxidant status. The effect of squid surimi enriched with glucomannan or glucomannan plus spirulina on liver and heart structures and cell damage markers was tested in fa/fa rats fed highly saturated-hyper-energetic diets. Animals were fed 70% AIN-93M rodent diet plus six versions of 30% squid surimi for 7 weeks: control (C), glucomannan (G), and glucomannan plus spirulina (GS). The cholesterol-control (HC), cholesterol-glucomannan (HG), and cholesterol-glucomannan plus spirulina (HGS) groups were given similar diets that were enriched with 2% cholesterol and 0.4% cholic acid. G and GS diets versus C diet significantly inhibited weight gain and lowered plasma alanine aminotransferase and aspartate aminotransferase, liver steatosis, lipogranulomas, and total inflammation and alteration scores. The hypercholesterolemic agent significantly increased the harmful effects of the C diet. Liver weight, the hepatosomatic index, all damage markers, and total histological scoring rose for HC versus C (at least P < .05). The addition of glucomannan (HG vs. HC) improved these biomarkers, and non-additional effects from spirulina were observed except for the total liver alteration score. In conclusion, glucomannan and glucomannan plus spirulina blocked the highly saturated-hyper-energetic diet negative effects both with and without added cholesterol. Results suggest the usefulness of including these functional ingredients in fish products.
Introduction
N
Fish derivates are attractive matrices for functional food development due to their health benefits, 8,9 their excellent rheology, and their reputation among consumers as a health food. 10,11 Owing to their satiating, laxative, and hypocholesterolemic properties, soluble viscose fibers are a good candidate for use as a functional ingredient. However, flatulence, abdominal pain, and esophageal or lower gastrointestinal obstruction have been reported when large amounts are ingested. 12 The addition of glucomannan to a squid-surimi matrix, accompanied by a diet that is rich in saturated fat, improved the plasma lipoprotein profile 13 and antioxidant status, but caused a mild increase in liver inflammation markers in fa/fa rats. 14,15 However, the mechanisms linking glucomannan consumption with increased inflammation remain unknown. Spirulina platensis is a microalga that is rich in minerals and antioxidant compounds such as carotenoids and phycocyanin. 16 When incorporated into glucomannan-enriched squid surimi, spirulina did not modify the plasma lipoprotein profile or antioxidant status but did significantly reduce the liver inflammation markers observed. 13 –15
To the best of our knowledge, aside from our research, no studies have been conducted on the beneficial or detrimental effects of glucomannan-enriched surimi consumption on liver or heart damage in obese rats. This article hypothesizes that glucomannan-enriched and glucomannan plus spirulina-enriched squid surimi act as functional foods by reducing the detrimental effects of highly saturated fat, either cholesterol enriched or not, in fa/fa rats. The purpose of our study was to determine the effects of consuming large amounts of glucomannan-enriched squid surimi on liver and heart injury biomarkers and histological inflammation parameters, as well as fat infiltration in the presence of a hyper-energetic, hypersaturated fat diet, enriched with cholesterolemic agents or not, and to assess the effects of its consumption in fa/fa rats with NAFLD.
Materials and Methods
Diet preparation and experimental design
This study was approved by the Spanish Science and Technology Advisory Committee (project AGL 2008-04892-C03-02 and Consolider Ingenio 2010, CSD 2007-00016) and by the ethics committee of the Universidad Complutense of Madrid (Spain). All experiments were performed in compliance with Directive 86/609/EEC of November 24, 1986 (amended by Directive 2003/65/CE) for the protection of scientific research animals. Forty-eight growing male Zucker fa/fa rats with an initial body weight of ∼120 g were obtained from Harlan Laboratories Models (Harlan, SL, Barcelona, Spain). The animals were housed in metabolic cells in a temperature-controlled room (22.3°C ± 1.9°C) with a 12-h light–12-h dark cycle. Rats were fed commercial pellets (Panlab, Barcelona, Spain) for a 7-day adaptation period and were then distributed into six groups of eight animals each.
Six experimental semi-synthetic diets were prepared: The control diet with no added cholesterol (C) was composed of a homogeneous mixture of 70% rodent diet (AIN-93M purified rodent diet; Dyets, Inc., Bethlehem, PA, USA) and 30% freeze-dried, restructured squid surimi (with 15% microcrystalline cellulose); the glucomannan normocholesterolemic diet (G) consisted of a mixture of modified AIN-93 feed (AIN M no. 102635, 70%) and freeze-dried, restructured glucomannan-enriched squid surimi (30%); the glucomannan plus spirulina normocholesterolemic diet (GS) consisted of a mixture of modified AIN-93M no. 102635 feed (70%) and freeze-dried, restructured glucomannan plus spirulina-enriched squid surimi (30%); the cholesterol-enriched control diet (HC) was identical to the C diet but with 2% cholesterol (95–98% purity) and 0.4% cholic acid (98% purity) replacing an equal amount of corn starch (modified AIN-93, AIN-93M no. 102636 diet); the cholesterol-enriched glucomannan diet (HG) was identical to the G diet but enriched with cholesterol (2%) and cholic acid (0.4%); and the cholesterol-enriched glucomannan plus spirulina diet (HGS) consisted of the GS diet enriched with cholesterol (2%) and cholic acid (0.4%). Details of diet composition have already been reported. 13 Diets were approximately isoenergetic (minimum 3.10 kcal/g diet for HC diet, maximum 3.40 kcal/g diet for GS diet). Moreover, of the total dietary energy, ∼32% was from palm olein and 13.6% was from saturated fatty acids. Water and food were provided ad libitum over the 7-week experimental period. To avoid inter-assay variations, fasting rats were taken one at a time from each group, anesthetized, and euthanized by extracting blood from the descending aorta with a syringe. Livers and hearts were collected immediately after blood extraction, weighed, and preserved in a 10% formaldehyde aqueous solution.
Liver and heart damage biomarkers
Plasma alkaline phosphatase (ALP), alanine aminotransferase (ALT), aspartate aminotransferase (AST), creatine phosphokinase (CK), and lactate dehydrogenase (LDH) were analyzed by using commercial kits according to the manufacturer's instructions in a Hitachi 737 Automatic Analyzer (Hitachi Ltd., Tokyo, Japan).
Histological procedures
Samples from a major liver lobe and the heart were fixed in 10% buffered formalin, dehydrated in alcohol gradient and xylene, and embedded in paraffin. Five-micrometer sections were obtained in a microtome (JUNG RM 2155; Leica, Solms, Germany) and were stained with routine hematoxylin and eosin. The histological evaluation was performed on each liver, studying two sections per liver, amplified at 4 × , 10 × , and 20 × by using a digital photomicroscope (Olympus BX50, Tokyo, Japan) with a 50DP integrated camera (Olympus BX50, Tokyo, Japan).
The effect of diets on liver was evaluated by using a modified version of the semi-quantitative histological scoring system reported by Kleiner et al. 17 The scores employed were as follows: steatosis (0, absence; 1, mild; 2, moderate; 3–4, severe), considering steatosis grade (0, <5%; 1, 5–33%; 2, >33–66%; 3, >66%), and contiguous patches of microvesicular steatosis (0, absent; 1, present), hepatocyte ballooning (0, absence; 1, mild; 2, moderate-severe), lipogranulomas (0, 0; 1, <2 foci per 20 × field; 2, 2–4 foci per 20 × field; 3, >4 foci per 20 × field), lobular inflammation, large lipogranulomas, and periportal inflammation values (0, absence; 1, presence in the last three parameters).
Heart damage was assessed by using a slightly modified version of the semiquantitative score system developed by Gürses et al. 18 The following scores were applied: inflammation (0, absence; 1, mild; 2, moderate; 3, intense), thrombus (0, absence; 1, presence), myocyte necrosis (0, absence; 1, 1–3 necrotic cells; 2, >3 necrotic cells), necrosis spread, fibrosis, and vascular fibrosis values (0, absence; 1, small area affected; 2, <5% affected; 3, 5–10% affected; 4, >10% affected).
Total inflammation or alteration scores were determined by using the Kleiner et al. formula
17
:
Statistical analysis
Results were expressed as means and standard deviations. A two-way ANOVA (cholesterol and diet) was used. Pair wise comparisons of diet responses between groups (C, G, and GS; and HC, HG, and HGS) were made by using the Bonferroni test. The T2-Tamhane post hoc test was applied when variances were assumed to be unequal. The effect of cholesterol consumption (HC vs. C; HG vs. G; and HGS vs. GS) was evaluated by using the unpaired Student's t-test. Parameters not exhibiting normal distribution in the histological study (fat infiltration, ballooning, inflammation) were studied by using the Montecarlo exact bilateral signification method. Groups were then compared pairwise by using the Fisher exact statistical test. Significant differences were set at P < .05. The SPSS version 22.0 statistical analysis package (SPSS, Inc., Chicago, IL, USA) was used.
Results
Body weight gain, and liver and heart somatic indexes
Body weight gain, liver weight, and hepatosomatic indexes were significantly affected by supplementary cholesterol and diet type (all P < .001), and by the interaction between diet and cholesterol (all P < .05) (Table 1). The G and GS diet significantly limited weight gain compared with the C diet. This effect was greater when cholesterol plus cholic acid were included in the diet. Liver weight and hepatosomatic indexes sharply increased in the presence of supplementary cholesterol (all P < .001), but the addition of glucomannan lessened this effect on liver size (G and GS vs. C and HG and HGS vs. HC) with no additional effect from spirulina.
Mean values within a row with unlike letters were significantly different (P < .05, Bonferroni test).
Mean values within a column for the same parameter were significantly different from their no supplementary cholesterol counterparts: * P < .05 and *** P < .001.
Liver and heart damage biomarkers
All the enzymes tested and the AST/ALT ratio were significantly affected by supplementary cholesterol and diet type (all P ≤ .001), and by the diet-cholesterol interaction (all P < .01) (Table 2). In comparison to the C diet, HC diet boosted all enzyme activities (400–1500%, all P < .001). The addition of glucomannan to diets with no added cholesterol significantly lowered ALT and AST activities compared with the C group (at least P < .05). No noticeable differences were observed between the GS and G groups. The HG and HGS diets significantly reduced all enzyme activities (at least P < .05) compared with their HC counterpart.
Mean values within a row with unlike letters were significantly different (P < .05, Bonferroni test).
Mean values within a column for the same parameter were significantly different from their no supplementary cholesterol counterparts: * P < .05, ** P < .01, and *** P < .001.
ALT, alanine aminotransferase; ALP, alkaline phosphatase; AST, aspartate aminotransferase; CK, creatine phosphokinase; LDH, lactate dehydrogenase.
Histological procedures
Table 3 and Figure 1 summarize the major histological liver changes caused by glucomannan-surimi and cholesterol ingestion. Steatosis, hepatocyte ballooning, lipogranulomas, periportal and total inflammation and alteration scores were significantly affected by glucomannan diets and cholesterol (all P < .05). C and HC diets induced the maximum steatosis scores. G and GS diets significantly reduced steatosis compared with the C diet. A similar tendency (P > .05) was observed in the HG and HGS groups versus HC. Total inflammation and alteration scores were significantly higher in HC compared with C animals (55% vs. 42.5% and 83.75% vs. 63.75%, respectively, all P < .05).
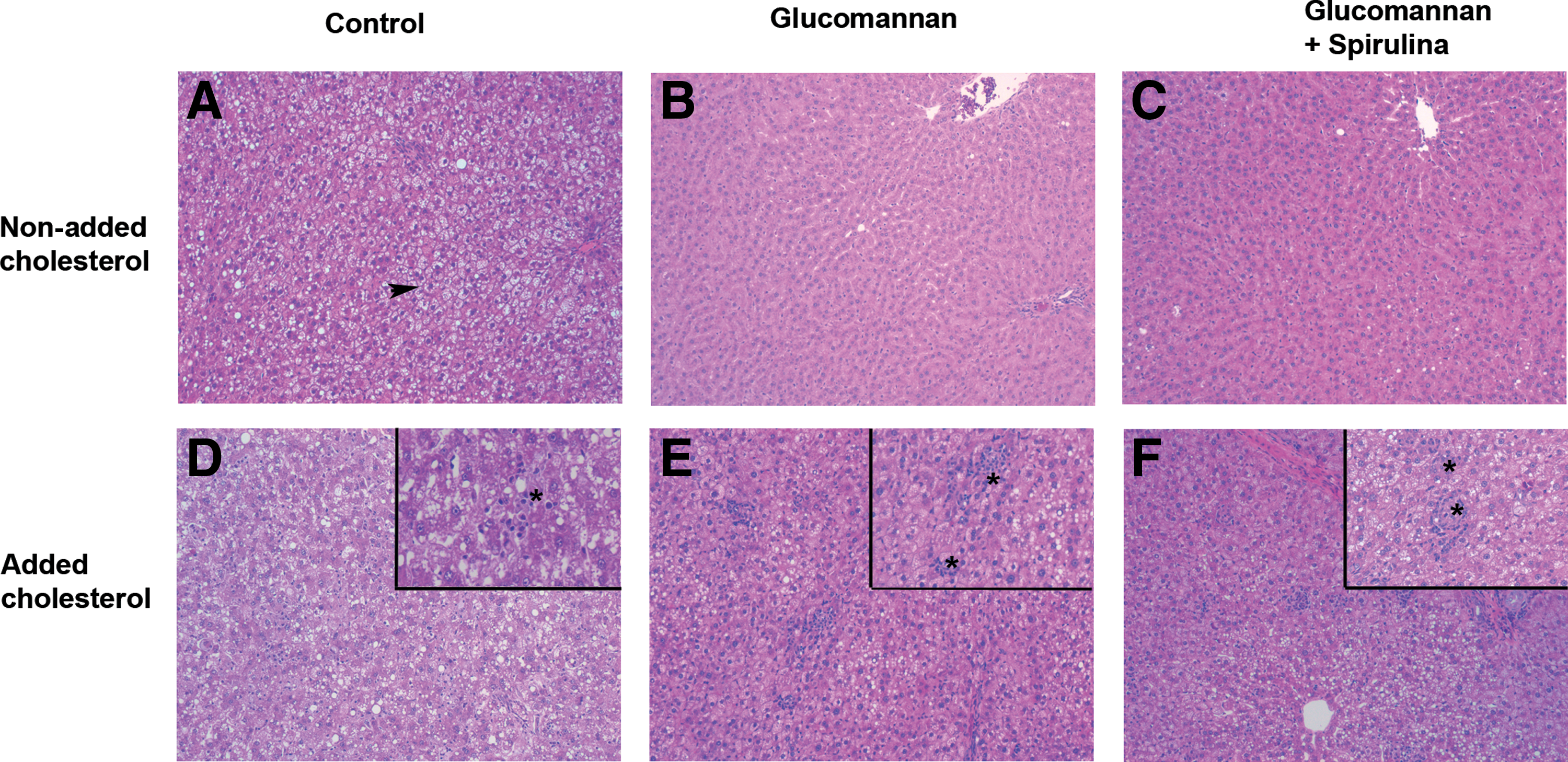
Comparison of different livers with no cholesterol
Steatosis values: 0, absence; 1, mild; 2, moderate; 3–4, severe. Hepatocyte ballooning values: 0, absence; 1, mild; 2, moderate-severe. Lipogranulomas values: 0, absence; 0, absence; 1, mild; 2, moderate; 3, severe. Lobular inflammation, large lipogranulomas, and periportal inflammation values: 0, absence; 1, presence.
Mean values within a column for the same parameter were significantly different from their no supplementary cholesterol counterparts (* P < .05, ** P < .01, and *** P < .001).
Montecarlo significance between normo- and hypercholesterolemic groups.
Montecarlo significance between the six experimental groups.
NA, non-applicable due to equivalent data distribution.
Total liver alteration scores were 8.75% and 16.25% for G and GS, respectively, which were significantly lower (P < .05) than C (63.75%). HGS presented a lower total liver alteration score (68.75%) than the HC and HG groups (83.75% and 82.5%, respectively).
Table 4 shows major heart histological changes. Extension of necrosis was the only parameter affected by diet (P = .036); cholesterol-fed rats tended to present higher scores. The addition of cholesterol negatively affected these heart scores (HC vs. C and HG vs. G; both P < .05).
Inflammation values: 0, absence; 1, mild; 2, moderate; 3, intense. Thrombus values: 0, absence; 1, presence. Myocyte necrosis values: 0, absence; 1, 1–3 necrotic cells; 2, >3 necrotic cells. Necrosis spread, fibrosis, and vascular fibrosis values: 0, absence; 1, small area affected; 2, <5% affected; 3, 5–10% affected; 4, >10% affected.
Mean values within a column for the same parameter were significantly different from their no supplementary cholesterol counterparts ( *P < .05).
Montecarlo significance between normo- and hypercholesterolemic groups.
Montecarlo significance between the six experimental groups.
Discussion
These results suggest that long-term ingestion of squid surimi with glucomannan or glucomannan/spirulina-enriched diets affects liver and heart structures, and it improves NAFLD biomarkers in growing Zucker fa/fa rats, mainly those fed cholesterol-enriched diets.
Body weight gain, and liver and heart somatic indexes
Both glucomannan and glucomannan plus spirulina restructured squid-surimi diets were eagerly consumed by growing fa/fa rats. Similarly, in a parallel study on Zucker fa/fa rats fed restructured pork, 19 glucomannan and glucomannan plus spirulina did not lower the amount of food ingested. As leptin resistance is a characteristic trait of fa/fa rats, 4 the satiating effect modulated by gastric leptin due to glucomannan expansion in the stomach does not affect these animals. This has been corroborated by similar or even lower ingestion data recorded in other studies. 2,20 As observed in other experiments with Zucker fa/fa rats, 21 results also suggest that high-energy, saturated fat diets induce obesity and considerable body weight gain. However, glucomannan added to squid surimi somehow limited dietary energy bioavailability and thus weight gain, coinciding with previous studies. 22,23 In contrast with other reports, 24 spirulina did not exhibit any significant effects on body weight.
In line with Beynen et al., 25 supplementary cholesterol added to the diet decreased growth rate in our study. Mahfouz and Kummerow 26 reported lower body weight gain in rabbits but not in rats fed cholesterol-enriched diets. The addition of cholesterol to the diet did not modify the effect of glucomannan or glucomannan plus spirulina on body weight.
Liver weight and the hepatosomatic index are used to investigate potential changes in liver size and hepatic steatosis. 27 C group values were similar to those of obese Zucker rats fed cocoa-rich diets. 28 The G diet reduced liver size to values similar to those found by Cordero-Herrera et al. 28 in lean Zucker rats.
Obese Zucker rat livers exhibit increased lipogenesis. 29 Gomez-Zorita et al. 30 suggest that resveratrol protects against the development of fatty liver by reducing oxidative stress and hepatic lipid content. As glucomannan or glucomannan plus spirulina exhibited antioxidant properties, 13,14 normalized lipemia, 15 and reduced liver weight (results of this study), we can hypothesize that experimental diets decreased liver lipogenesis and NAFLD in non-cholesterol-fed fa/fa rats.
HG and HGS diets drastically reduced hepatomegalia observed in HC rats, suggesting potential benefits of this fiber in preventing fatty liver. Spirulina in the diet did not have any effect on this parameter.
Heart weight and cardiosomatic index were similar to those observed by Noratto et al. in fa/fa rats. 31 Consumption of diets with added cholesterol tended to increase the cardiosomatic index.
Liver and heart damage biomarkers
ALT, AST, and ALP values of the C group coincide with those of Gomez-Zorita et al. 30 but are higher than those found in Sprague-Dawley rats affected by NAFLD disease. 32 Zucker fa/fa rats are a well-known model for NAFLD. 33 Low AST/ALT ratios have been identified as an NAFLD-independent predictor, 34 where ALT values are higher than AST. The AST/ALT ratios in the C group were approximately one third of those found in Wistar rats fed fried sardines 9 or restructured meat containing algae, 3 clearly suggesting that fa/fa rats are prone to NAFLD. The AST/ALT ratio was 0.91 in the C group and ∼1.6 in the G and GS groups, suggesting reduced hepatosteatosis in these non-cholesterol groups. A positive correlation between liver weight and the AST/ALT ratio was observed in cholesterol-fed animals.
The ALT/ALP ratio, identified as a cholestasis indicator, exhibits a significant positive correlation with liver weight and the hepatosomatic index (r = 0.465; P = .022; and r = 0.420, P = .041, respectively) in the non-cholesterol-fed groups. The ALT/ALP ratio in C rats was 3.6 times higher than that found in Wistar rats, 3 suggesting the presence of cholestasis in this fa/fa rat group. The addition of glucomannan sharply decreased this ratio, suggesting the positive effect of glucomannan on cholestasis. Based on the ALT/ALP ratio, severe cholestasis and hepatocellular damage could be hypothesized in cholesterol-fed rats, as was reported by Schultz Moreira et al. 3
Large amounts of CK and LDH are present in the heart and can be released into the bloodstream in cases of heart damage or inflammation. However, LDH is also a known marker of cell necrosis 35 and hepatocellular damage. 9,36 Both enzyme levels were relatively high in C rats in comparison to adult Wistar rats 37 but lower than those found in Sprague-Dawley rats, 38 suggesting that C diets have a mildly negative effect on the heart and liver. However, the addition of glucomannan (G and GS diets) decreased (P < .05) LDH activity but did not alter CK, suggesting liver cell protection. As CK and LDH activities increased in HC rats, it can be inferred that the ingestion of cholesterol clearly damaged both organs. Glucomannan added to hypercholesterolemic diets relevantly blocked the negative effects of hypercholesterolemia on both damage markers, but spirulina did not have any additional effect on those enzymes. Sánchez-Muniz et al. 9,36 found a marked increase in LDH activity in cholesterol-fed rats, which was sharply diminished by the action of omega-3 fatty acids.
Histological procedures
Figure 1 shows that C rat livers had relatively high hepatosteatosis based on their lobulillar space structures, with total alteration scores reaching 63.8%. As suggested by the AST/ALT ratio, hepatosteatosis was greatly reduced by the glucomannan and glucomannan plus spirulina diets. Yki-Jarvinen 1 and results of a previous study 13 unequivocally suggest that glucomannan should improve the action of insulin in inhibiting both the production of glucose and triglycerides transported by very low density lipoproteins (VLDL-triglyceride) levels, resulting in lower compensatory hyperinsulinemia and hypertriglyceridemia. However, periportal inflammation tended to increase slightly in G rats. As already mentioned, Vázquez-Velasco et al. 14 found that some inflammation markers (tumor necrosis factor alpha, inducible nitric oxide synthase) increased in G rat livers and this was the most likely related to the high degree of fermentation of glucomannan.
Total inflammation and total alteration but not steatosis scores rose in HC animals. The consumption of HG and HGS diets induced lower liver steatosis than HC rats, which was in line with the reduction of the AST/ALT ratio already mentioned. Spirulina, probably due to its minor compounds and omega-3 fatty acids, had additional positive effects on liver structure, decreasing total alteration score even at low doses. Omega-3 fatty acids have been found to partially block the negative effects of cholesterol ingestion on liver damage markers. 9,36 However, it should be noted that additional changes induced by spirulina were modest.
The relatively high activities of CK and LDH in C rats coincide with the myocyte necrosis and necrosis extension observed (Scoring 15 out of 16 and 15 out of 32, respectively). However, no other harmful histological changes were evident. G and GS diets did not have any effect on the heart parameters tested nor on the total alteration score. However, cholesterol ingestion (HC vs. C rats) negatively affected total heart alteration. Fibrosis tended to increase in HGS versus HG, suggesting a potential negative effect of spirulina plus glucomannan.
Total liver and heart alteration scores would appear to be useful markers, as significant correlations were found between these scores and the damage markers tested (data not shown). In fact, marker values and total alteration scores were lower in HG and HGS compared with HC animals and similar to those observed in C rats.
Potential limitations of this article include: (a) only growing male fa/fa rats were used; (b) the study was performed with only one amount of glucomannan or glucomannan plus espirulina added to squid surimi. Future studies should assess potential benefits of different doses of glucomannan/and glucomannan plus espirulina-squid surimis and should likewise include a wider age range of fa/fa rats.
This study included a large amount of fiber in an obesogenic diet to be tested on a metabolic syndrome animal model. When adapting a glucomannan-surimi diet to humans predisposed to or suffering from metabolic syndrome, the amount of this soluble fiber should be considered and probably lowered because of the side effects observed in humans. 12 Moreover, the amount of glucomannan added to surimi may be correlated to the severity of the symptoms of the metabolic syndrome. In any case, the significant effects observed on cholestasis markers may be particularly relevant for patients with a bile tract condition.
Conclusions
These results suggest that glucomannan and/or glucomannan plus spirulina added to squid surimi act as a functional food in an obese and NAFLD animal model. The authors, therefore, encourage future studies to assess the usefulness of these products in humans who are affected by non-alcoholic fatty liver disease.
Footnotes
Acknowledgments
This study was supported by the Spanish project AGL-2014–53207-C2-2R. The authors gratefully acknowledge the foreign fellowship for graduate studies granted by the Consejo Nacional de Ciencia y Tecnología (CONACYT) of México to Laura González-Torres. All authors have significantly contributed to this article and agree with its present version. M.V.-V. is the corresponding author and guarantor of the article, contributed to the study design, data discussion, and writing of the article; L.G.-T. and R.A.G.-F. contributed to data acquisition and analysis and writing of the article; and F.J.S.-M., S.B., J.B., and M.J.G.-M. contributed to data discussion and critically reviewed the article.
Author Disclosure Statement
No competing financial interests exist.