
Abstract
In cell culture studies, dried plum (Prunus domestica L.) polyphenols increased osteoblast alkaline phosphatase (ALP) activity, mineralized nodule formation, and the expression of the bone marker genes runt-related transcription factor 2 (RUNX2) and osterix. The purpose of this study was to determine whether human serum collected 1 and 2 h after dried plum ingestion influenced osteoblast cell activity and gene expression. Five healthy women ingested 100 g of dried plum, and serum samples were collected at baseline (before dried plum ingestion) and 1 and 2 h postingestion of dried plum. MC3T3-E1 osteoblast cells were treated (2% of medium) with these serum samples for 3 or 9 days. Intracellular and extracellular ALP activities were significantly increased after 3 or 9 days of treatment with serum both postingestion time points, with no effect seen in baseline samples. Also, serum obtained 1 and 2 h postingestion significantly increased the mRNA expression of bone markers RUNX2 and connexin43 (CX43) after both 3 and 9 days of incubation periods. Finally, serum obtained 1 and 2 h postingestion increased the mRNA expression of β-catenin after 9 days of incubation. We conclude that osteoblast activity and function are increased by dried plum ingestion, which may, in part, explain its beneficial effects on bone health.
Introduction
A
Among fruits and vegetables tested, dried plum has shown promising results in the prevention and reversal of bone loss. Deyhim et al. 4 demonstrated that a diet with 5% of dried plum by weight could restore bone mass in ovariectomized female rats. When Bu et al. 5 compared supplementation with dried plum with treatment with parathyroid hormone in male rats, similar improvements in bone microarchitecture and bone mineral density (BMD) were observed in both groups. In addition, the bone-protective effects of dried plum are not limited to hormone deficiency-induced bone loss; one animal study has shown that dried plum (25% of diet) could also reduce bone loss because of aging. 6 Furthermore, dried plum has been shown to be more effective than other fruits with high antioxidant activity in restoring bone in osteopenic ovariectomized mice. 7 In two short- and long-term clinical trials, the effects of dried plum on bone biomarkers and BMD were measured in postmenopausal women. Arjmandi et al. 8 found that supplementation with 100 g dried plum over 3 months improved the bone formation markers, and Hooshmand et al. 9 showed that supplementation with 100 g dried plum over 1 year significantly improved the BMD by suppressing bone turnover in postmenopausal women.
Dried plum has a high content of polyphenols and there is growing evidence suggesting that polyphenols are responsible for the significant effect on bone metabolism. In vitro studies on osteoblast cells or preosteoblast cells suggest that dried plum polyphenols significantly affect the cells' activities and fates. 10 Preosteoblast cells treated with polyphenols extracted from whole dried plum demonstrated increases in osteoblast cell differentiation, mineralized nodule formation, and upregulation of bone marker genes. 10 Furthermore, in cell culture studies, dried plum polyphenols have strong anti-inflammatory and antioxidant properties, contributing to the regulation of bone remodeling. 11 All in vitro studies examining the effects of dried plum polyphenols have used polyphenols extracted from dried plum. A significant limitation of this approach is that polyphenols are metabolized in the body after dried plum ingestion, thus clinical insight is somewhat limited. In this study, we address this limitation by studying human blood serum collected in the hours after dried plum ingestion. Specifically, the purpose of this study was to investigate the effect of human serum collected 1 and 2 h after the ingestion of 100 g of dried plum on osteoblast cell activity and mRNA expression of bone formation markers in osteoblasts.
Materials and Methods
Human intervention
Participants
Five healthy women (age range 25–33 years) participated in this study. Sample size of five was considered adequate for in vitro cell culture studies. All women were premenopausal and were studied during the first week of their menstrual cycle. None of the women were on birth control and none had any previous report of bone loss. Subjects had no evidence of any cardiovascular disease or other chronic diseases. A bone density scan was performed using dual energy X-ray absorptiometry on each participant (Table 1). The Institutional Review Board at San Diego State University approved all procedures involving human subjects.
Mean ± SD; n = 5 for dried plum regimen.
BMD, bone mineral density; BMI, body mass index; SD, standard deviation.
Materials
Dried plum (Prunus domestica L.) was obtained from California Dried Plum Board (Sacramento, CA). All the reagents used for the blood draws were obtained from Fisher Scientific (Waltham, MA, USA).
Study design
The day before laboratory visit, subjects were provided with a standardized evening meal (2.25 g carbohydrate/kg, 0.5 g fat/kg, 0.375 g protein/kg). 12 Subjects were instructed to consume the provided meal before starting an overnight 10-h fast period. Blood samples were collected after 10-h fasting between 8:00 a.m. and 10:00 a.m. Subjects were asked to consume 100 g dried plum immediately after the baseline blood draw. The amount of dried plum was based on previous research demonstrating that consumption of 100 g dried plum prevented bone loss in osteopenic postmenopausal women. 9,13,14 Additional blood samples were collected 1 and 2 h postingestion of dried plum. The time intervals between dried plum ingestion and blood draws were defined based on anticipated polyphenol bioavailability kinetics. 15 Immediately after collection, blood samples were centrifuged at 1500 g for 10 min at 4°C, and serum was separated and stored at −80°C for later use on cell culture experiments.
Cell culture experiments
Materials and reagents
MC3T3-E1 mouse calvarial preosteoblastic cells were obtained from ATCC (Manassas, VA, USA). Fetal bovine serum (FBS), penicillin G–streptomycin, and trypsin/EDTA were purchased from Fisher Scientific. Minimum essential medium (α-MEM) without phenol red was purchased from Life Technologies (Carlsbad, CA, USA). Ascorbic acid and β-glycerophosphate were purchased from Sigma-Aldrich (St. Louis, MO, USA). The alkaline phosphatase (ALP) substrate colorimetric kit was purchased from Abcam (Cambridge, MA, USA). Unless otherwise listed, all other chemicals were reagent grade and obtained from Fisher Scientific (Pittsburgh, PA, USA).
Experiment design
In vitro cell culture experiments were performed using MC3T3-E1 preosteoblast cells isolated from mouse calvaria. The purpose of these experiments was to test the effect of human blood serum obtained 1 and 2 h after ingestion of 100 g dried plum on the activity of preosteoblast cells in culture. The cells were grown on 75 cm flasks for 48 h before start of the experiment. Once at 80% confluence, cells were plated on six-well plates at a density of 1 × 105 cells/well on α-DMEM with 1% penicillin and streptomycin and 10% FBS at 37°C in a humidified atmosphere of 95% air and 5% CO2. Cells were allowed to adhere for 24 h. Culture medium was replaced with differentiation medium: α-DMEM with 1% penicillin and streptomycin, 25 μg/mL of ascorbic acid, and 10 mM B-glycerophosphate without serum for 2 h. The medium was replaced with differentiation medium containing 2% human serum from each of the three time points (baseline, 1 and 2 h after ingestion of dried plum) or 2% FBS (negative control). Cells were incubated with treatment medium for either 3 or 9 days. Treatment medium was replaced every third day. 10 Both cell culture medium and cells were collected at 3 or 9 days from all treatment groups for ALP activity analysis and RNA extraction for gene expression analysis. All cell culture experiments were performed three times and each time in duplicate.
Extracellular and intracellular ALP activities
Cell culture medium was collected after 3 and 9 days of treatment to measure extracellular ALP activity. The level of ALP activity was measured by the conversion of p-nitrophenyl phosphate to p-nitrophenol at 405 nm spectroscopy according to the manufacture's protocol. ALP activity was expressed in nanomole of p-nitrophenol produced per minute per μg/mL of protein. For intracellular ALP activity, cells were collected after 3 and 9 days of treatment, and lysed according to the manufacture's protocol (Abcam) before analysis.
Gene expression analysis using real-time polymerase chain reaction
Materials and reagents
RNA extraction kit RNeasy and QIAshredder columns were purchased from Qiagen (Germantown, MD). Real-time polymerase chain reaction (RT-PCR) primers, SYBR green mix, RNase-free water, plates, and cDNA reverse transcription kit were purchased from BioRad (Hercules, CA, USA).
RT-PCR gene expression
Total cellular RNA was extracted using the RNeasy extraction kit according to manufacturer's directions. RNA was eluted in 30 μL water and stored at −80°C for later use. The RNA purity and concentration were determined by optical density (OD) measured at 260 and 280 nm. RNA was reversed transcribed into cDNA using the following conditions: reverse transcription at 58°C for 10 min, inactivation of reverse transcriptase at 95°C for 30 sec, and hold at 4°C. Each cDNA reaction mixture (20 μL) was diluted 200 × and stored at −20°C. For specific gene expression RT-PCR analysis, the primers listed in Table 2 were used. All the cDNA samples were quantified and 40 ng of cDNA of each sample was loaded into each reaction mixture using the following conditions: polymerase activation and cDNA denature at 95°C for 30 sec, denature at 95°C for 15 sec, and annealing/extension at 60°C for 30 sec for 40 cycles. Finally, post-PCR melting curves confirmed the specificity of single-target amplification. CFX96 (Biorad, Irvine, CA, USA) was used for all the RT-PCR experiments and the CFX96 software was used to analyze the data.
Transcript sequence (5′–3′).
Cx43, connexin43; OPG, osteoprotegerin; RANKL, receptor activator of nuclear factor kappa-B ligand; RUNX2, runt-related transcription factor 2
Statistical analysis
Data are presented as mean ± standard deviation. Values were compared using t-tests to determine that means were significantly different. SPSS software (version 22.0) was used to perform the statistical analysis. Significance was acceptance at P ≤ .05.
Results
Effect of human serum after dried plum ingestion on intracellular and extracellular ALP activities
After 3 days of treatment, intracellular ALP activity increased twofold only in serum collected 2 h postingestion of dried plum in comparison with the baseline serum (Fig. 1). After 9 days of treatment, intracellular ALP activity increased gradually in serum collected 1 and 2 h postingestion of dried plum (Fig. 1). Treatment with human serum 2 h postingestion of dried plum increased intracellular ALP activity fourfold higher compared with treatment with baseline human serum after 9 days. In addition, treatment with human serum collected 1 and 2 h postingestion of dried plum significantly increased (P < .05) extracellular ALP activity after both 3 and 9 days of treatment (Fig. 2).
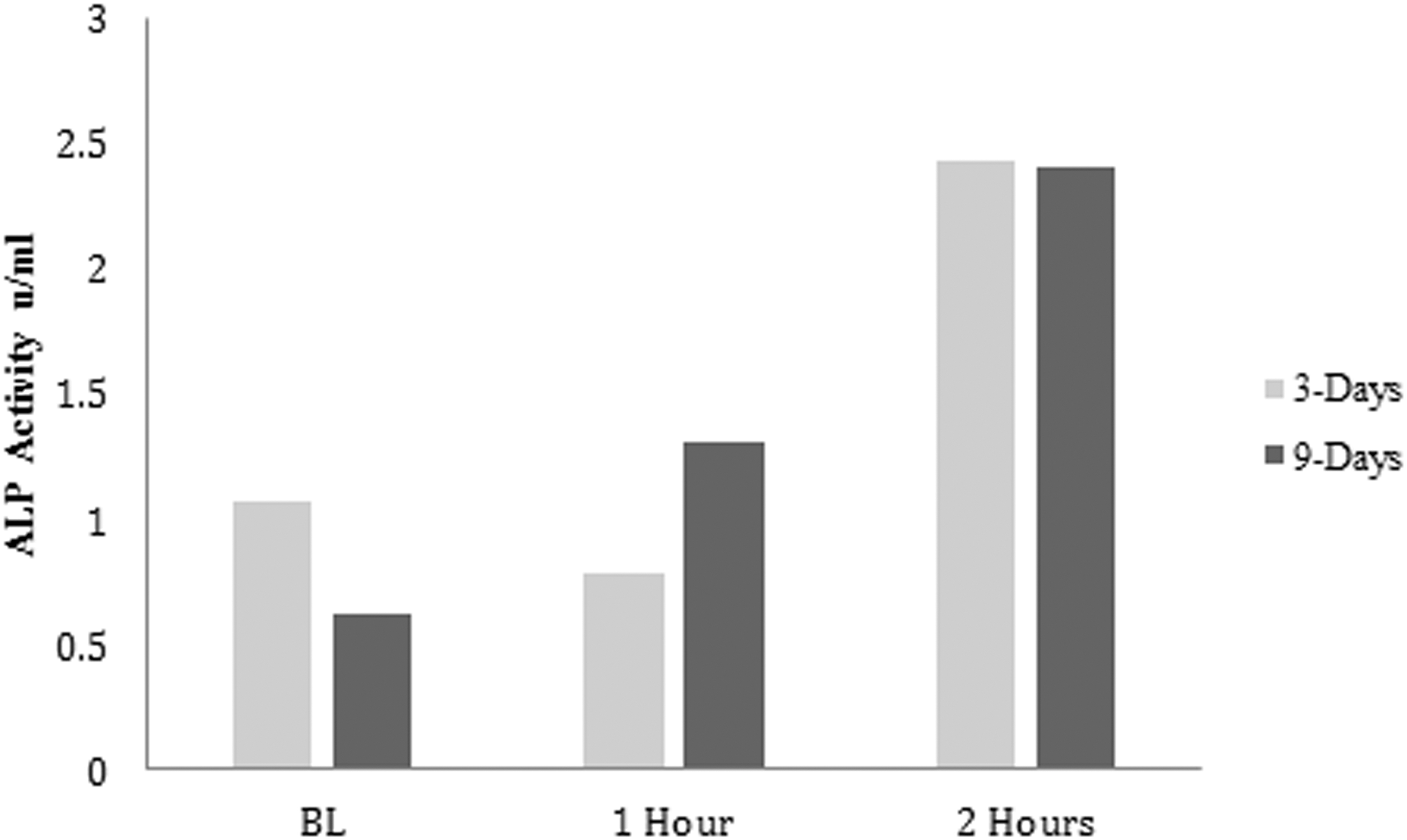
The effect of treatment with human serum postingestion of dried plum on intracellular ALP expression after 3 and 9 days of treatment. Cells were plated at a density of 1 × 105 cells/mL, treated with human serum from BL, 1, and 2 h postingestion of dried plum for 3 and 9 days. This assay was performed on one cell culture experiment. ALP, alkaline phosphatase; BL, baseline.
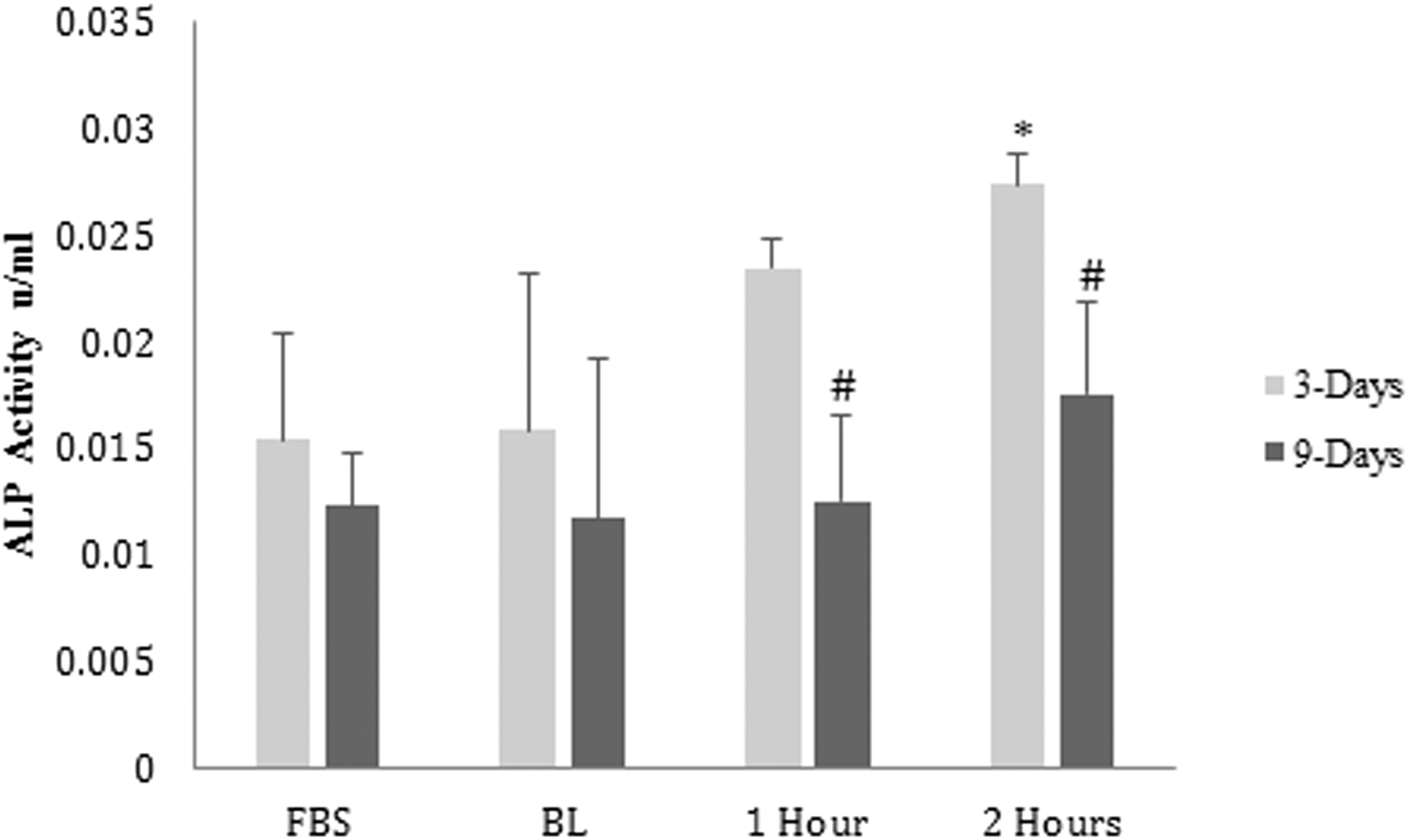
The effect of treatment with human serum postingestion of dried plum on extracellular ALP expression after 3 and 9 days of treatment. Cells were plated at a density of 1 × 105 cells/mL, treated with human serum from BL, 1, and 2 h postingestion of dried plum for 3 and 9 days. This assay was performed in triplicates. Bars with an asterisk (*) were significantly different (P < .05) compared with the group treated with the baseline serum for 3 days, and bars with a pound sign (#) were significantly different (P < .05) compared with the group treated with the baseline serum for 9 days. FBS, fetal bovine serum.
Effect of human serum after dried plum ingestion on gene expression
Three days of treatment of MC3T3-E1 osteoblast cells with human serum collected 1 h postingestion of dried plum increased the expression of the transcription factors runt-related transcription factor 2 (RUNX2) and osterix 1.7-fold and 1.5-fold, respectively (Fig. 3). Treatment with human serum collected 2 h postingestion of dried plum increased expression of RUNX2 1.9-fold (Fig. 3). In addition, treatment with human serum collected 2 h postingestion of dried plum increased the mRNA expression of β-catenin (Fig. 3). There was no detectable change in expression of osteoprotegerin (OPG) and connexin43 (Cx43) after 3 days of treatment with serum collected 1 and 2 h after dried plum ingestion.
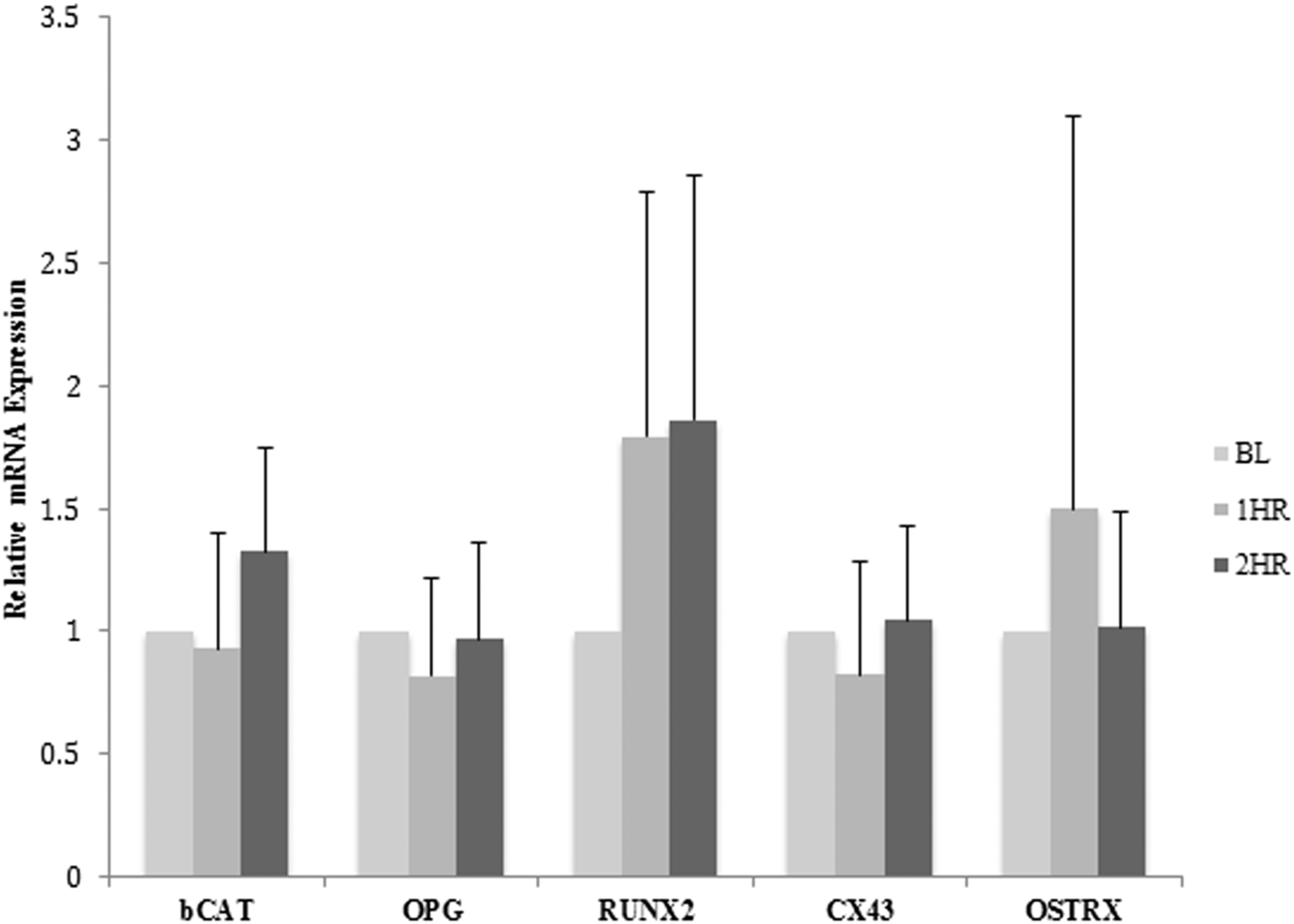
Effect of treatment with human serum postingestion of dried plum for 3 days on relative gene expression of osteoblasts. Relative alteration in gene expression levels of bone marker genes and β-catenin (bCAT) after treatment with human serum collected at BL, 1 h (1HR), and 2 h (2HR) postingestion of dried plum for 3 days. Cells were plated at a density of 1 × 105 cells/mL. Cx43, connexin43; OPG, osteoprotegerin; OSTRX, osterix; RUNX2, runt-related transcription factor 2.
Treatment of MC3T3-E1 osteoblast cells with human serum collected 1 and 2 h postingestion of dried plum for 9 days increased the expression of the transcription factor RUNX2 and the gap junction Cx43 (Fig. 4). β-catenin mRNA expression was also increased after 9 days of treatment with human serum collected 1 and 2 h postingestion of dried plum. Expression of RUNX2 and Cx43 increased up to 1.5-fold and 1.9-fold, respectively, whereas the expression of β-catenin increased up to 2.5-fold after 9 days of treatment (Fig. 4). There were slight increases in expression of OPG after 9 days of treatment with human serum 1 and 2 h postingestion of dried plum. No change in osterix was detected after 9 days of treatment (Fig. 4) in contrast to 3 days of treatment. No change in receptor activator of nuclear factor kappa-B ligand gene expression was detected in MC3T3-E1 cells treated with human serum collected 1 and 2 h postingestion of dried plum after 3 or 9 days of treatment (data not shown).
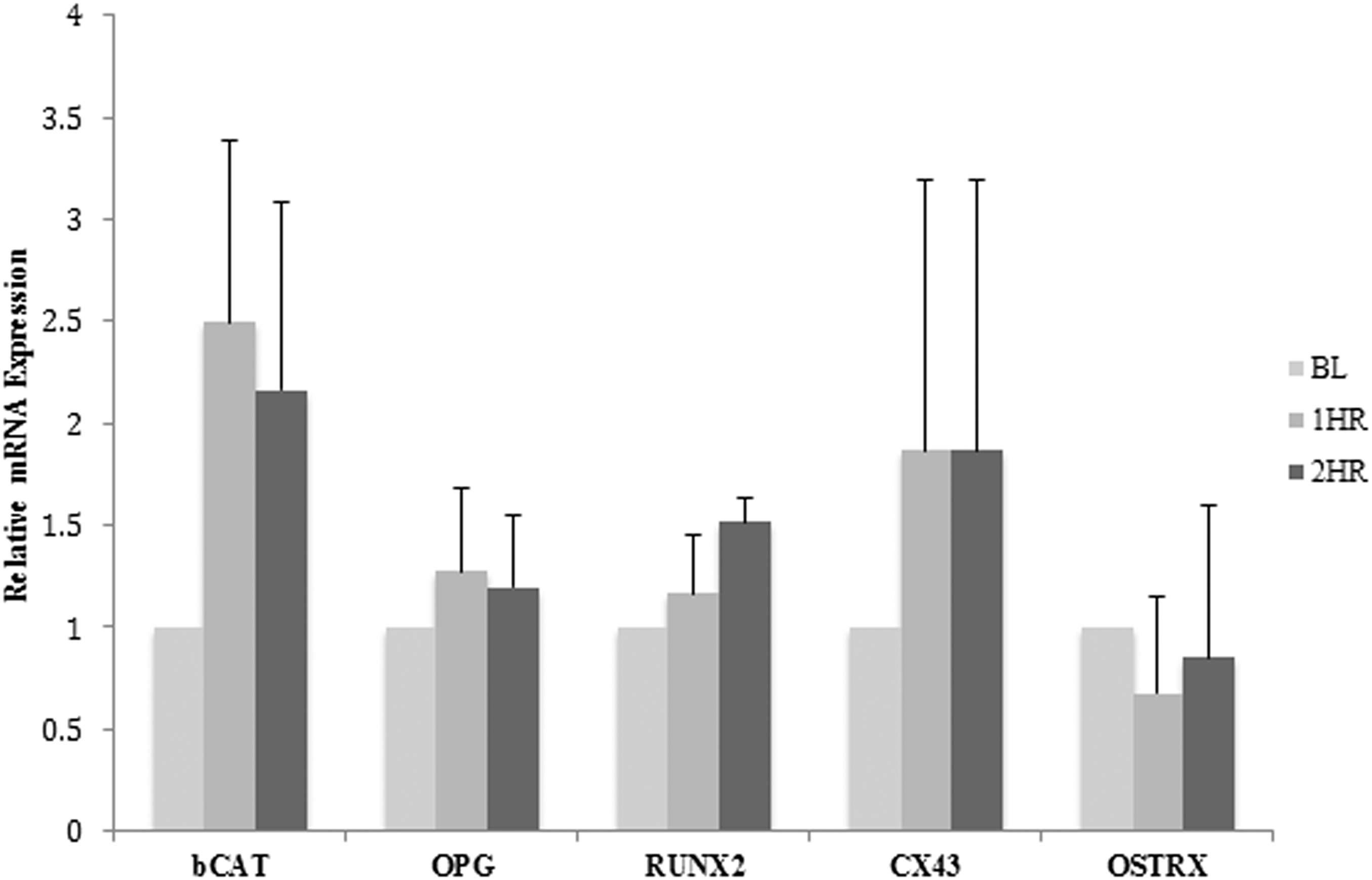
Effect of treatment with human serum postingestion of dried plum for 9 days on relative gene expression of osteoblasts. Relative alteration in gene expression levels of bone marker genes and β-catenin (bCAT) after treatment with human serum collected at BL, 1 h (1HR), and 2 h (2HR) postingestion of dried plum for 9 days. Cells were plated at a density of 1 × 105 cells/mL.
Discussion
This study demonstrates that human serum collected 1 and 2 h postingestion of dried plum stimulates osteoblast activity as shown by increases in intracellular and extracellular ALP activities. Changes in ALP activity were at least partly caused by the upregulation of the expression of transcription factors and genes involved in bone formation and osteoblast activity.
ALP is known to promote osteoblast activity by providing the inorganic phosphate needed for the bone mineralization process. 10 The increases in ALP activities were concomitant with increases in transcription factor RUNX2 and gap junction Cx43, which are two recognized bone formation markers. 16 RUNX2 is a transcription factor that is essential for osteoblast differentiation from pluripotent cells. 17 Active nuclear RUNX2 upregulates skeletal genes and consequently the osteoblast phenotype. 16 Mutation of the RUNX2 gene in mice models resulted in mice with cartilaginous skeletons and demonstrated that RUNX2 is required for bone formation. 18 Cx43 is the most highly expressed gap junction protein in bone, and recent research has found that it plays an essential role in bone remodeling and function. 19 Cx43 forms gap junctions that allow communication between adjacent cells or between a cell and the extracellular matrix. 20 Mutation of Cx43 leads to osteoblast dysfunction shown by a significantly reduced expression of bone markers and delayed ossification. Also, recent animal studies have shown that Cx43 plays a critical role in preventing osteoblast and osteocyte apoptosis. 21 Cx43 null mice exhibited an increase in apoptosis rate when treated with bisphosphonates, suggesting that Cx43 is required for bisphosphonates to be effective in protecting against apoptosis. 20 The role of Cx43 in the prevention of bone cell apoptosis was also shown by in vitro studies demonstrating that bisphosphonates treatment of osteocytes revealed that Cx43 hemichannels are required for bisphosphonates effectiveness. 17
In addition to the effects on RUNX2 and Cx43 expression levels, treatment with human serum 1 and 2 h postingestion of dried plum stimulated the expression level of β-catenin, a key signaling molecule in the Wnt signaling pathway. Previous research has shown that absence of β-catenin caused early onset of severe osteoporosis, as well as a decrease in OPG mRNA compared with the wild type mice. 22 Several studies have suggested that the polyphenols in the dried plum are responsible for the effect on the bone cells. 9 –11,13,14,23 By treating cells with human serum collected after dried plum ingestion, our method takes into consideration the fact that dried plum polyphenols are metabolized in the body after dried plum ingestion. This method more closely resembles the state of the dried plum polyphenols in the circulation in human studies that have demonstrated that ingestion of 100 g of dried plum significantly increases BMD in postmenopausal women. 9,14
In summary, we have demonstrated that treatment with human serum 1 and 2 h postingestion of dried plum for 3 and 9 days led to an increase in osteoblast activity and function by increasing ALP activity and parallel increased gene expression of bone markers and β-catenin. This in vitro research reinforces research demonstrating that dietary ingestion of dried plum may favorably improve bone formation and provides plausible mechanisms that may be responsible. The gene expression level alteration presented in this study resulted from a short-term acute treatment with the human serum. Therefore, it may or may not represent the long-term effects of dried plum on bone remodeling. It is also important to note that the effects of human serum after acute dried plum ingestion on osteoblast cell culture reported here may not represent an environment where osteoblasts and osteoclasts interact in the human body. In addition, polyphenols were not measured in this study in blood samples because of limited quantity of blood samples collected. Future studies should investigate the effects of human serum postingestion of dried plum on cocultures of osteoblasts and osteoclast cells. Also, further studies are needed to clarify whether Wnt signaling plays a role in the mechanism by which dried plum ingestion stimulates bone formation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.