
Abstract
Chia oil has the highest recognized α-linolenic acid (ALA) content. ALA is associated with beneficial changes in plasma lipids and the prevention of cardiovascular diseases. Present article aims to analyze the effect of Chia oil–enriched restructured pork (RP) on aged rats in a nonalcoholic steatohepatitis (NASH) model. Groups of six male Wistar rats (1-year old) were fed the experimental diets: control RP diet (C) noncholesterol high saturated; cholesterol-enriched high-saturated fat/high-cholesterol control RP diet (HC) with added cholesterol and cholic acid; and Chia oil- or Hydroxytyrosol RP cholesterol–enriched high-saturated fat/high cholesterol (CHIA and HxT). Total cholesterol, hepatosomatic index, Nrf2, antioxidant, and inflammation markers were determined. CHIA reduced the hypercholesterolemic effect by lowering levels similar to C; also, ameliorated redox index. CHIA, despite high polyunsaturated fatty acids (PUFA) content, reduced thiobarbituric acid reactive substances (TBARS) and induced the lowest SOD protein synthesis but not a reduction on its activity. Chia oil activated the Nrf2 to arrest the pro-oxidative response to cholesterol and aging. Endothelial nitric oxide synthase (eNOS) system was lower in HxT than in CHIA, suggesting its antiatherogenic activity and related protective effect against high PUFA. Increase in tumor necrosis factor alpha (TNFα) was partially blocked by CHIA. Chia oil has the ability to prevent oxidative damage and modify the inflammatory response, suggesting adequate regulation of the antioxidant system. Results stress the importance of incorporating ALA into the diet.
Introduction
M
NAFLD is now one of the most common liver diseases worldwide. Its pathogenesis includes high dietary fat intake, insulin resistance, oxidative stress, and genetic background. It is also frequently associated with obesity, type 2 diabetes (T2DM), dyslipidemia, metabolic syndrome, and cardiovascular diseases, 5 degenerative diseases in which ageing is a relevant and known risk factor. As it has been suggested that there are similarities between 1-year-old rats and 60-year-old men, we selected rats of this age for our animal model. 6 In recent years, it has been suggested that NAFLD is impaired by a liver excess of ω-6 and deficiency of ω-3 polyunsaturated fatty acids (PUFA), which cause the impairment of ω-6/ω-3 ratio. 7
In the process of obtaining healthier meat products, a number of modifications could be made to add, reduce, or increase substances with proven biological properties (functional ingredients). 8 Previous studies by our group have shown the beneficial effects that restructured pork (RP) containing seaweed, glucomannan, or silicon have on plasma cholesterol and antioxidant status. 6,9,10
Some epidemiological studies have suggested that high dietary concentrations of α-linolenic acid (ALA, 18:3 ω-3) are associated with plasma lipid concentrations and improvements in cardiovascular diseases. 11,12 It has also been demonstrated that ALA, as precursor of eicosapentaenoic acid (EPA, C20:5 ω-3) and docosahexaenoic acid (DHA, C22:6 ω-3), is involved in the generation of anti-inflammatory lipid mediators due to the derivatives formed after its consumption. 13,14
In this regard, interest in ALA as a functional ingredient has grown in the recent years. However, ALA is not readily available as there are few foods that can provide adequate amounts to obtain the benefits associated with ω-3 consumption. 15
Salvia hispanica L. seeds, commonly known as chia seeds, are the richest source of ALA and contain high amounts of fiber and minerals. 11 Chia seed lipid content varies from 20% to 40%, with 60% of total lipids comprising ALA and 20% of linoleic acid (ω-6). 16,17 The remaining extracted oil contains 34/100 g of dietary fiber and 17/100 g of protein. 17 –19 Chia oil is also rich in magnesium and phenolic compounds (mainly quercetin and kaempferol). Chia seed has proved to be a potent antioxidant; 20,21 its calcium and potassium content suggests positive influence on high blood pressure. 22
As a powerful antioxidant with widely functional properties, hydroxytyrosol (HxT) has been used as a functional food ingredient, especially in PUFA-enriched muscle foods. HxT displays a great capacity to inhibit lipid oxidation in foodstuffs rich in fish lipids 23 and in precooked frankfurters enriched with ω-3 PUFA. 24 In addition, its in vivo antioxidant properties, when incorporated into RP in high fat diets, have already been evaluated by our group. 25
The hypothesis underlying this study was that using Chia oil as a functional ingredient in the meat product formulations has a positive effect on the antioxidant and inflammatory status partially arresting (blocking) the negative effects induced by the ingestion of cholesterol. The aim of this work is to evaluate whether chia oil–enriched RP may have beneficial properties by reducing liver oxidation and inflammation in aged rats, in the frame of a high-cholesterol/high-saturated fat diet as a NASH model.
Materials and Methods
Diet preparation and experimental design
Experimental diets were designed to have high-saturated fat/high-cholesterol/high-energy content, based on high amounts of sucrose and saturated fat. Three of these diets were enriched with hypercholesterolemic inductors (cholesterol plus cholic acid). Chia oil and hydroxytyrosol [HxT, 2-(3,4-dihydroxyphenyl)-ethanol; SeproxBiotech, Madrid, Spain] were used in diet preparation. Minced lean pork and lard were purchased at a local shop. Lean pork (849 g) and fat (151 g) were homogenized, and RP was prepared following standard procedure. 9 For Chia-RP, chia oil (152.2 g/kg fresh matter) was homogenized with lean pork and the rest of the components. In the case of HxT-RP, hydroxytyrosol (3.6 g/kg fresh matter) was previously homogenized with lard and then mixed with the rest of components. The different RP mixtures were freeze-dried in a LyoAlfa 10 freeze dryer (Telstar, Terrassa, Spain). Four experimental semisynthetic diets were prepared: (1) control RP diet (C), high-saturated fat with no added cholesterol; (2) cholesterol-enriched high saturated fat/high cholesterol control diet (HC), where maize starch was replaced by 1.26% cholesterol and 0.25% cholic acid; and (3) and (4) Chia- and HxT-RP cholesterol-enriched high saturated fat/high cholesterol diets (CHIA and HxT, respectively), similar to HC but incorporating chia oil and HxT. To formulate experimental diets, 217 g of RP and 783 g of a modified semisynthetic formulation (reference U8959 version 180; Panlab S.L., Barcelona, Spain) were mixed using serial dilutions until fully homogeneous. Diets were calculated to cover micronutrient requirements at the final concentration (Table 1). Twenty-four male Wistar rats were obtained from Harlan Laboratories Models, SL. (Barcelona, Spain). Animals were housed under controlled conditions (22.3°C ± 1.8°C and 12-h light/12-h dark cycle). Rats were fed commercial pellets (Panlab) during 1-week adaptation to environmental conditions. Water was provided ad libitum. When rats were 1-year old and weighed ∼500 g, they were distributed into four groups of six animals each, housed individually and fed the experimental diets daily for 8 weeks. Food consumption was measured daily and body weight once per week. No unexpected deaths were registered during the study. At the end of experimental period, rats were taken one at a time from each of the four groups, anesthetized with isoflurane (5%), and euthanized by extracting blood by heart puncture. Livers were dissected, weighed, and frozen using liquid nitrogen until analysis. All experiments were performed in compliance with Directive 86/609/EEC of November 24, 1986 (amended by Directive 2003/65/EEC of July 22, 2003) on the protection of scientific research animals. This study was approved by the Spanish Science and Technology Advisory Committee (project AGL 2011-29644-C02-02) and by the Ethics Committee of the Universidad Complutense de Madrid (Spain).
Data calculated considering as energy equivalent for carbohydrates: 16.73 kJ/g (4.0 kcal/g); fat, 37.65 kJ/g (9.0 kcal/g); protein 16.73 kJ/g (4.0 kcal/g).
As reported by Reeves et al. 58
Restructured pork composition available as Supplementary Data (Supplementary Table S1; Supplementary Data are available online at
C, control RP diet; CHIA, chia oil RP cholesterol–enriched high saturated fat/high cholesterol diet; HC, cholesterol-enriched high saturated fat/high cholesterol control RP diet; HxT, hydroxytyrosol RP cholesterol–enriched high saturated fat/high cholesterol diet; MUFA, monounsaturated fatty acids; PUFA, polyunsaturated fatty acids; RP, restructured pork; SFA, saturated fatty acids.
Plasma cholesterol
Plasma was separated by centrifugation at 2000 g for 10 min. Total plasma cholesterol was determined by the enzymatic colorimetric method using the Spinreact Kit (Sant Esteve de Bas, Girona, Spain).
Biochemical assays
Homogeneous samples of liver tissue were used for biochemical assays. Oxidized (GSSG) and reduced (GSH) glutathione levels in liver homogenates were determined by the Hissin and Hilf method. 26 Liver tissue was homogenized in a phosphate-EDTA buffer (0.1 M sodium phosphate and 0.005 M EDTA, pH 8) at 100 mg/mL, with the addition of 10 μL/mL of perchloric acid. It was then centrifuged at 10,600 g for 10 min at 4°C. Concentrations were calculated using a standard GSH and GSSG curve. Fluorescence was determined in a LS50 Perkin–Elmer (Baconfield, United Kingdom) fluorimeter at 350 and 420 nm (excitation and emission wavelengths, respectively). Results are expressed as μg glutathione/mg protein. The calculated redox index (RI) indicates the antioxidant status of tissue obtained as follows: RI = GSSG/(GSH+GSSG).
Lipid peroxidation (TBARS) was determined by measuring the malondialdehyde (MDA) formed after adding thiobarbituric acid [20 mM] to liver extracts. This assay is based on the ability of the thiobarbituric acid to react with certain compounds (reactive substances) resulting in lipid peroxidation, such as MDA. Absorbance levels were measured at 532 nm in triplicate. The calibration curve prepared with 1,1,3,3-tetraethoxypropane was taken as a reference to determine the final concentration of MDA, and results are expressed as mg MDA/mg protein. 27
Superoxide dismutase (SOD) activity was measured using a slight modification of the Nitroblue Tetrazolium (NBT) method.
28
This method is based on superoxide radical (O2
•−) production by auto-oxidation of hydroxylamine hydrochloride which, in the presence of NBT, is reduced to nitrite. The reaction between nitrite and EDTA forms colorimetric complexes that were measured at 560 nm. Catalase (CAT) activity was measured as described by Aebi
29
using hydrogen peroxide as substrate. The decomposition of H2O2 was followed directly by a decrease in absorbance at 260 nm. Endothelial nitric oxide synthase (eNOS) activity was determined by spectrophotometrically measuring the conversion of
Biochemical assays were standardized to liver homogenate protein concentrations determined according to Bradford. 31 Final enzyme activity results were expressed as IU/mg protein.
Western blotting and enzyme levels
Total liver protein lysates were obtained and separated in 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Gels were then blotted onto PVDF Amersham Hybond-P membranes (GE Healthcare, Buckinghamshire, United Kingdom) and incubated with their corresponding antibodies (anti-β-actin [A2228], anti-catalase [C0979], anti-superoxide dismutase [S2147] from Sigma-Aldrich, St. Louis, Missouri, USA; anti-glutathione peroxidase [Ab59546], anti-inducible nitric oxide synthase [Ab21775], anti-endothelial nitric oxide synthase [Ab5589] from Abcam, Cambridge, United Kingdom; anti-tumor necrosis factor alpha [sc-1350], anti-glutathione reductase [sc-32886] from Santa Cruz Biotechnology, Dallas, Texas, USA). β-Actin was used as loading control. Blots were developed by enhanced chemiluminescence using an Amersham ECL Plus Western Blotting Detection Reagent (GE Healthcare) according to manufacturer's instructions.
Extraction and quantification of RNA by reverse transcription-polymerase chain reaction
RNA samples were isolated from 100 mg of liver using TRI-Reagent (Sigma-Aldrich) according to manufacturer's instructions and treated with DNase I RNase-free reagents (Thermo Fisher Scientific, Waltham, Massachusetts, USA) to remove any contamination with genomic DNA. The yield and quality of RNA were assessed by measuring absorbance at 260, 280, and 310 nm and by electrophoresis on agarose gels (1%). Total RNA of each sample (1 μg) was reverse-transcribed to first-strand complementary DNA (cDNA) using a Revert Aid H Minus cDNA Synthesis Kit (Thermo Fisher Scientific).
Relative CAT, Mn-SOD, CuZn-SOD, glutathione reductase (GR), and nuclear factor erythroid 2-related factor 2 (Nrf2) mRNA levels were quantified with a LightCycler Real Time PCR Detection System (Roche diagnostics, Indianapolis, Indiana, USA), using SYBR Green (Biotools, Madrid, Spain) as the fluorescent binding dye. Detection was monitored by measuring the increase in fluorescence throughout the cycles.
All sample mRNA levels were standardized to their β-actin values and results expressed as fold changes in the threshold cycle (Ct) value relative to controls using the 2−ΔΔCt method. 32 PCR parameters were as follows: preincubation at 95°C for 10 min followed by 45 cycles of denaturation at 95°C for 10 sec with an annealing temperature of 60°C for each couple primer, extension 72°C for 15 sec, and cooling at 40°C for 30 sec. Primer sequences were as indicated in Table 2.
CAT, catalase; GR, glutathione reductase; Nrf2, nuclear factor erythroid 2-related factor 2; SOD, superoxide dismutase.
Statistical analyses
All experiments were performed in triplicate. Data were analyzed using the SPSS Statistics v22 software (IBM Corporation, Somers, NY, USA). Results are expressed as means and standard deviations. The student's t-test was used to analyze the effect of cholesterol (HC vs. C groups). One-way analysis of variance (ANOVA) followed by Bonferroni tests was used to assess the effect of CHIA and HxT groups versus HC group. Where variances were assumed to be unequal, the T2 of the Tamhane post hoc test was applied. Results were accepted as significant when P < .05.
Results
Body and liver weight
Table 3 shows the effects of cholesterol and chia oil- or HxT-RP addition on liver size and final body weight of aged rats fed the experimental diets. HC significantly increased liver weight and the hepatosomatic index, but lowered final body weight (all P < .05) compared to C rats. Liver weight (P = .001) and final body weight (P = .004) were significantly different among rats fed cholesterol-enriched diets. Final body weight showed no differences when comparing CHIA and HC groups. HxT was significantly lower than HC. Liver weight of HxT rats was significantly lower (P < .05) than those of HC, with no significant differences between CHIA and HC.
Mean ± SD (n = 6). ANOVA, statistical effect of ingredients in cholesterol-fed groups. P, statistical comparison between C and HC. Values with different superscript letters in the same row indicate significant differences (at least p < 0.05) among HC, HxT, and CHIA groups.
Hepatosomatic index = 100 × (liver weight/body weight).
Redox index = GSSG/(GSH+GSSG).
ANOVA, analysis of variance; GSH, reduced glutathione; GSSG, oxidized glutathione; SD, standard deviation.
Total plasma cholesterol
The effect that the addition of chia oil- or HxT-RP and cholesterol in diets had on plasma cholesterol is shown in Table 3. Dietary cholesterol significantly increased plasma cholesterol levels in HC (P = .037) compared to C rats. Significant differences were observed among HC, HxT, and CHIA groups (P = .002). HxT and CHIA showed lower plasma cholesterol content than HC (P < .05).
Oxidized and reduced glutathione levels and lipid peroxidation
Higher GSSG levels (P = .003) were exhibited in HC rats compared to their C counterparts. However, no differences were observed in GSH levels (P = .684) or the RI (P = .282) between these rats. HxT and CHIA rats exhibited significantly lower GSSG values (at least P < .05) than their HC counterparts. No significant differences were observed in the RI between CHIA and HxT rats (Table 3).
Significant differences were observed in lipid peroxidation (as TBARS) when comparing C and HC rats (P = .001). CHIA presented higher values than HxT, although both were lower compared to HC (P = .010).
Enzyme Western blot levels
Table 4 shows the protein levels from antioxidant enzymes determined by Western blot. SOD and GPx were higher, while CAT and GR were lower (at least P ≤ .05) in HC versus C rats. Significant differences among HC, CHIA, and HxT (ANOVA P ≤ .001) were found, with HxT exhibiting the highest protein level for CAT and GR. CHIA increased CAT levels with respect to HC, but lowered all of the enzyme protein levels versus HxT (at least P < .05).
Mean ± SD (n = 6). ANOVA, statistical effect of ingredients in cholesterol-fed groups. P, statistical comparison between C and HC. Values with different superscript letters in the same row indicate significant differences (at least P < .05) among HC, HxT, and CHIA groups.
Inflammation index = eNOS/(eNOS+iNOS).
eNOS, endothelial nitric oxide synthase; GPx, glutathione peroxidase; iNOS, inducible nitric oxide synthase; TNFα, tumor necrosis factor alpha.
Results from inflammation biomarkers such as protein levels determined by Western blot are shown in Table 4. HC did not present any significant differences in eNOS, inducible nitric oxide synthase (iNOS), or tumor necrosis factor alpha (TNFα) compared to C. HxT livers exhibited (at least P < .05) the lowest eNOS and higher iNOS and TNFα levels compared to HC rats.
Enzyme activities and expressions
Figure 1 shows the effect of experimental diets on antioxidant enzymes. CAT and SOD activities were significantly lower (P < .001), while eNOS activity was higher (P < .001) in HC rats versus C rats (Fig. 2). CAT, Mn-SOD, CuZn-SOD, and Nrf2 values were lower (P ≤ .001) in HC rats compared to C ones. Significant differences were found for activities and expressions between cholesterol-fed rat groups (ANOVA at least P = .044), except for GR (P = .513) and eNOS (P = .388) expressions and CAT (P = .663) activity. HxT exhibited the highest total SOD activity (at least P < .05) among all groups and exhibited higher Mn-SOD and CuZn-SOD expressions than HC rats (at least P < .05). CHIA presented the highest Nrf2 expression of all groups. HxT diet significantly affected the inflammation index with lower values than its HC and CHIA counterparts (P < .001).
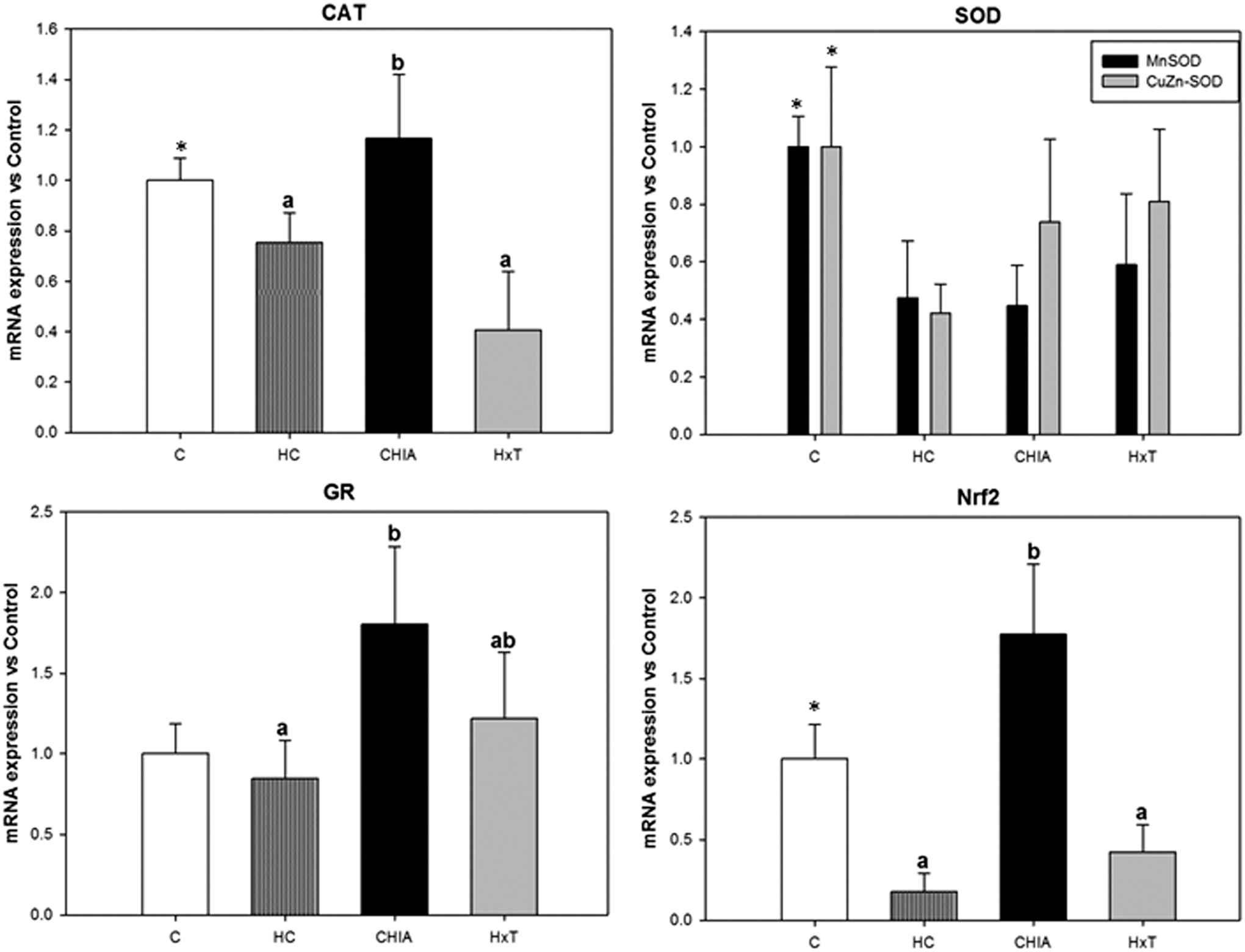
Effect of hydroxytyrosol or chia-restructured pork on liver enzymatic antioxidants determined by reverse transcription-polymerase chain reaction in aged rats. Mean ± SD (n = 6). *Indicate significant differences (at least P < .05) between C and HC. Bars with different letters indicate significant differences (at least P < .05) among HC, HxT, and CHIA. C, control RP diet; HC, cholesterol-enriched high saturated fat/high cholesterol control RP diet; HxT, hydroxytyrosol RP cholesterol–enriched high saturated fat/high cholesterol diet; CHIA, chia oil RP cholesterol–enriched high saturated fat/high cholesterol diet. CAT, catalase; SOD, superoxide dismutase; GR, glutathione reductase; Nrf2, nuclear factor erythroid 2-related factor 2; RP, restructured pork; SD, standard deviation.
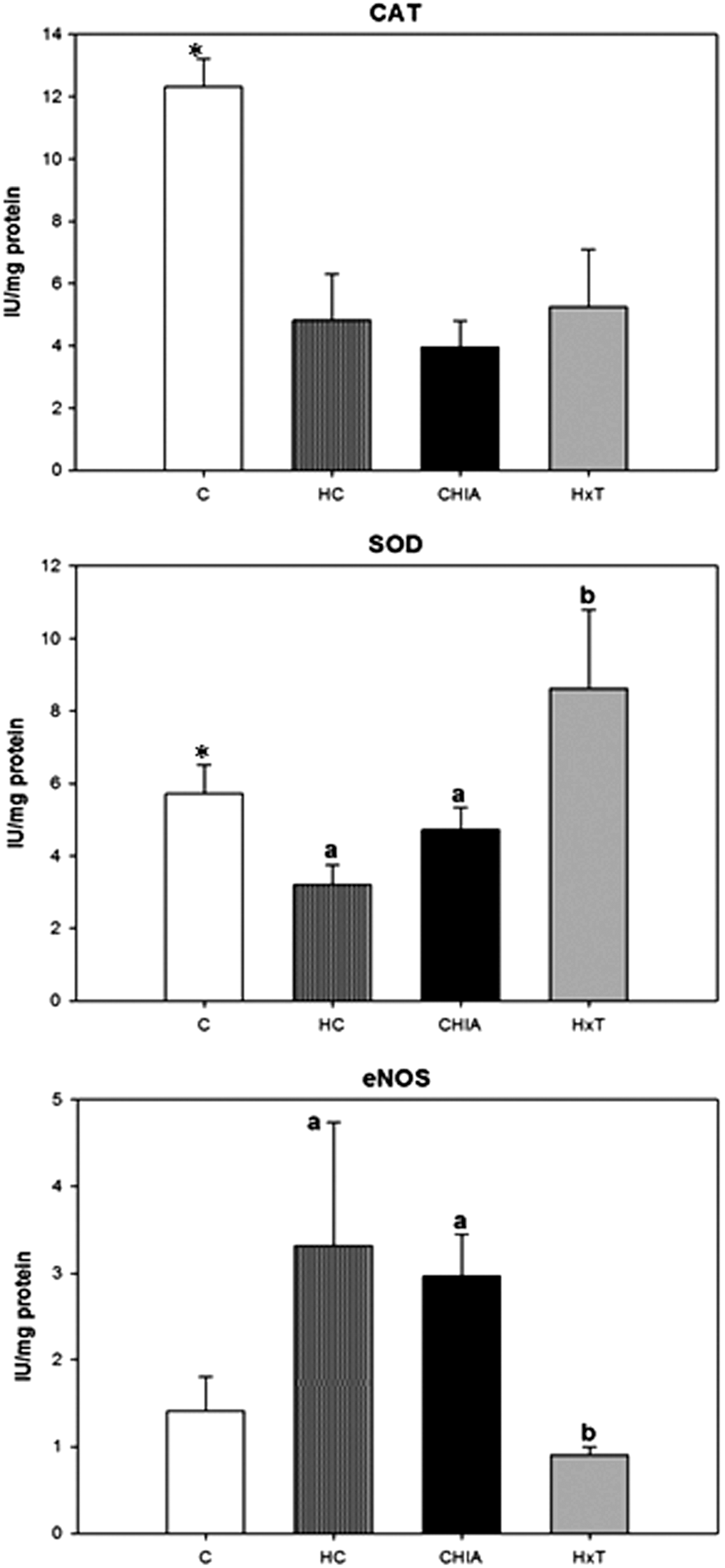
Effect of hydroxytyrosol or chia-restructured pork on enzymatic activity in liver of aged rats. Mean ± SD (n = 6). *Indicate significant differences (at least P < .05) between C and HC. Bars with different letters indicate significant differences (at least P < .05) among HC, HxT, and CHIA. eNOS, endothelial nitric oxide synthase.
Discussion
This study explored, for the first time, the antioxidant and anti-inflammatory effects of RP formulated with chia oil on aged rats as part of a NASH model. Our findings suggest that chia oil affects the antioxidant and inflammatory mechanisms in response to dietary atherogenic inductors and age.
Results suggest that the intake of a control RP incorporated in a high cholesterol–high fat diet produces the typical effects observed in NASH, such as increased plasma cholesterol or liver enlargement as has been shown in previous studies. 10,33 In agreement with Beynen et al., 34 cholesterol ingestion reduced body weight. Previous results of our group suggest the highly lipolytic effect of cholesterol ingestion affecting the expression of hormone-sensitive lipase adipocyte enzyme accounting for the decrease in body and adipose tissue fats. 35 Cholesterol esterification with fatty acids in the liver has been proposed as an important mechanism for the control of cholesterolemia. 36,37 The effect on body weight was generally lower in HxT-RP (although nonsignificant, P > .1), suggesting that this functional ingredient may modify the effect of dietary cholesterol on adipocyte fat and enzyme expression. Nonetheless, as no differences were observed in the food intake values between groups, the results may be attributed to other metabolic facts as the thermogenic effect of HxT, which enhances noradrenaline and adrenaline excretions, suppresses body fat accumulation, and decreases the weight of adipose tissue by increasing triglyceride catabolism as reported by Oi-Kano et al. 38 This hypothesis will be tested in future studies. Furthermore, CHIA diet reduced the hypercholesterolemic effect of dietary cholesterol observed in HC group by bringing plasma levels down to those of the C diet. Chia seeds and lipids have been described as cholesterol regulators owing to the effect of PUFA in lipid metabolism. 39 Olmedilla-Alonso et al. 40 found that the consumption of restructured meat products with added walnuts (rich in linolenic acids) lowers plasma cholesterol in high cardiovascular risk subjects.
The results of this work show HxTs ability to significantly reduce liver weight in comparison to the other cholesterol-enriched diets leading to a reduction in hepatic steatosis. 25 This effect was significantly lower in the case of the CHIA diet. Although, no clear explanation is available, the effect on the liver seems to be linked to the liver cholesterol-ester “pool” enlargement as found in a previous study on rats that were fed cholesterol- and sardine-enriched diets. 41
Ageing induces changes in several elements of the main endogenous antioxidant system in the case of older animals. 42 In addition, dietary components may directly affect defense mechanisms protecting against exogenous stress leading to unexpected levels of nonenzymatic antioxidants (e.g., glutathione levels) and modification of enzymatic pathways. Studies have shown that GSH decreases with age, whereas the GSSG and/or GSSG/GSH ratio increases. 42 Other studies have suggested that elevated levels of dietary fat lead to high lipid peroxidation indirectly modifying GSH levels and glutathione enzyme activity. 10,43 In fact, our findings show that GSH levels were significantly lower than those of GSSG in the C group. It has been shown that dietary fat and hypercholesterolemia affect cell membrane integrity thus affecting defense against oxidative damage 44 and, therefore, the exogenous inclusion of antioxidants may not be enough to reverse the negative effects of ageing. Nonetheless, HxT and CHIA diets improved the RI with respect to their HC counterpart, although the mechanism involved appears to be different, as CHIA tended to raise GSH levels and HxT to lower GSSG levels. Similar results on GSH and GSSG levels and the RI were observed following the consumption of walnut-enriched RP. 45 HxT exhibits antioxidant and antiatherogenic biological properties. 46,47 The incorporation of chia oil or HxT into RP compared to their HC counterparts in the frame of a NASH model reduced oxidative damage by regulating GSSG levels through increasing GR and decreasing GPx protein levels.
An increase in TBARS and/or MDA values indicates increased lipid peroxidation. The HC group displayed a significant increase of lipid peroxidation, fact that would lead to tissue damage and failure of the antioxidant defense mechanisms to prevent excess reactive oxygen species (ROS) formation. 48 These results concur with others found in cholesterol-fed rats. 9 CHIA diet, despite their high PUFA content, was able to reduce TBARS levels, with respect to HC diet.
Data also suggest that hydrogen peroxide, as ROS, was mainly controlled by the SOD-CAT pathway. Results indicated lower enzymatic activity levels of SOD and CAT than those found in previous studies in Wistar rats 9 but similar to Vázquez-Velasco et al. 10 in fa/fa rats. The CHIA diet induced the lowest SOD protein synthesis but not a reduction in its activity. This could be due to a sufficient increase in enzyme activity to reduce oxidative damage making additional protein synthesis of more protein unnecessary. The HxT diet did not bring about any change in CAT activity with respect to the HC diet only exhibiting a trend toward low CAT expression balanced by higher levels of CAT protein synthesis.
Nrf2 is a key component in cellular redox homeostasis involved in the attenuation of oxidative stress processes. Its activation is related to GSH synthesis, lipid metabolism, and inflammation. 49,50 Previous results obtained by our group clearly suggest that hypercholesterolemia induced a significant reduction in the mRNA levels of antioxidant enzymes giving rise to oxidative stress. 25,33 Nrf2 was significantly reduced by cholesterol ingestion explaining the arrest in CAT. Our findings suggest that chia oil and HxT, in minor proportion, were exogenous stimuli that promoted the activation of Nrf2 to limit the pro-oxidative response to dietary cholesterol and ageing. Previous studies have demonstrated that chia seeds are exogenous stimuli for the activation of detox alternative mechanisms in cells, by modifying the activity of certain antioxidant enzymes related to the Nrf2 pathway. 39 The incorporation of chia oil- or HxT-RP to the NASH model may activate Nrf2 and spark the expression of phase II detoxifying enzymes conferring additional antioxidant protection and, thus, enhancing the antioxidant defense system and damage prevention of reactive oxygen and nitrogen species. 51
eNOS and iNOS results are difficult to explain, as cholesterol ingestion increased eNOS activity, but reduced the expression of eNOS and iNOS. A previous study shows that cholesterol ingestion increased both eNOS and iNOS expression keeping the inflammation index stable with respect to the control diet. 52 However, the two studies differ in several aspects such as age, type of rats, and amount of hypercholesterolemic inducer used. Excessive NO formation by NOS may lead to peroxynitrite-induced injury 53 and also cause cell damage through the direct inhibition of mitochondrial respiratory chain enzymes or oxidative protein modifications such as the formation of nitrotyrosine. 54 In addition, eNOS is regulated by enzyme level and the interaction of protein with other components, and therefore, protein synthesis was not proportionally related with total enzyme activity. 55 The incorporation of Chia oil clearly has a different effect compared to HxT on the eNOS system, as eNOS protein levels and activity were lower in the HxT group. These results suggest the antiatherogenic activity of CHIA, but can also be related to a protective effect against the high PUFA content of this diet.
ROS has been found to induce TNFα expression. 56 TNFα results appear to be relevant, as the increase observed in this inflammation marker following the HC diet was partially blocked by HxT and CHIA diets. Mells et al. 57 reported increased TNFα in a high-fat, high-cholesterol fed metabolic syndrome murine model, suggesting a clear relationship between cholesterol ingestion and liver inflammation. The reduction of lipid peroxidation (as TBARS) may clearly affect TNFα expression. Unpublished results suggest that the incorporation of CHIA reduced the presence of inflammatory cell infiltration observed in HC rat, at least partially explaining the results observed for TNFα.
This article has the merit of being the first to study the effect that extracted chia oil incorporated into RP has on a number of antioxidant and inflammatory markers in aged rats in the frame of a NASH model. Moreover, the study was long enough to assess quite permanent effects. Chia was tested against a recognized and studied antioxidant and anti-inflammatory ingredient. In conclusion, when incorporated into pork matrices in the context of atherogenic diets, chia oil has the ability to prevent oxidative damage and modify the inflammatory response in aged rats. This protection was more effective in some cases than HxT and suggests regulation of antioxidant enzyme expression and activity in the main enzymatic system. These results stress the importance of incorporating ALA into the diet and conducting further studies on other animal and human models.
Footnotes
Acknowledgments
All authors have significantly contributed to the article and agree with the present version of the article. F.J.S.-M. and J.B. contributed to the study design, data discussion, and writing of the article. J.A.S.L. contributed to data acquisition and analysis and writing of the article. A.G. contributed to the data acquisition and analysis. M.B.-A., M.J.G.-M., M.E.L.-O., S.B., and J.B. contributed to the data discussion and made a critical review of the article. The present study was supported by AGL 2014-53207-C2-2-R. J.A.S.-L. received the foreign PhD studies fellowship from CONACYT-México.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.