
Abstract
3,3′-diindolylmethane is a major in vivo metabolite of indole-3-carbinol, a bioactive compound found in cruciferous vegetables. Although 3,3′-diindolylmethane has been implicated to possess antitumorigenic and anti-inflammatory properties, the effect of 3,3′-diindolylmethane on adipogenesis has not been explored previously. Thus, the present study was conducted to determine if 3,3′-diindolylmethane affects adipogenesis using 3T3-L1 adipocytes and Caenorhabditis elegans. Treatment of 3,3′-diindolylmethane significantly reduced fat accumulation without affecting viability in 3T3-L1 adipocytes. 3,3′-diindolylmethane suppressed expression of peroxisome proliferator-activated receptor γ (PPARγ), CCAAT-enhancer-binding protein α (C/EBPα), fatty acid binding protein 4 (FABP4), and perilipin. In addition, 3,3′-diindolylmethane activated AMP-activated protein kinase α (AMPKα), which subsequently inactivated acetyl CoA carboxylase (ACC), resulting in reduced fat accumulation. These observations were further confirmed in C. elegans as treatment with 3,3′-diindolylmethane significantly reduced body fat accumulation, which was partly associated with aak-1, but not aak-2, orthologs of AMPKα catalytic subunits α1 and α2, respectively. The current results demonstrate that 3,3′-diindolylmethane, a biologically active metabolite of indole-3-carbinol, may prevent adipogenesis through the AMPKα-dependent pathway.
Introduction
O
As one bioactive found in cruciferous vegetables, indole-3-carbinol was reported to be effective in reducing fat accumulation in both in vivo murine obesity model 3,4 and in vitro murine adipocyte studies. 5,6 However, it is known that indole-3-carbinol forms a condensation product, 3,3′-diindolylmethane, under the acidic environment of the stomach. 7 Based on the observation that 3,3′-diindolylmethane, not indole-3-carbinol, has been shown to be the active component in vivo, 8 it is generally suggested that 3,3′-diindolylmethane is a major metabolite that mediates beneficial and functional activities of indole-3-carbinol. In comparison to indole-3-carbinol, there is currently no report of 3,3′-diindolylmethane and fat accumulation. Thus, the purpose of the current study was to determine the role of 3,3′-diindolylmethane in adipogenesis. For the first time, we tested effect of 3,3′-diindolylmethane on adipogenesis using 3T3-L1 adipocytes, which is a well-established adipocyte model. 9 We also tested effect of 3,3′-diindolylmethane on accumulation of triglycerides in Caenorhabditis elegans. C. elegans is a free-living nematode that has been extensively utilized in life science research due to its various advantages over other vertebrate animal models. 10 In particular, genes of C. elegans are completely sequenced, and relevance of their genes to humans is well understood. 11 –13 It was also previously reported that the central mechanisms of energy metabolism are conserved from mammals to C. elegans and its lipid metabolisms were reviewed well recently. 11,14 The current study was also to determine the potential mechanism focusing on AMP-activated protein kinase α (AMPKα) in both models.
Materials and Methods
Materials
3T3-L1 murine preadipocytes were purchased from American Type Culture Collection (Manassa, VA, USA). Nematode strains Wild-type N2, RB754 [aak-2(ok524)] and AGD397 [aak-1(tm1944) III; aak-2(ok524) X; uthEx202] and Escherichia coli OP50 used in this study were obtained from Caenorhabditis Genetics Center. 3,3′-diindolylmethane was purchased from Enzo Life Science (Farmingdale, NY, USA) and dissolved in dimethyl sulfoxide (DMSO) with 0.1% final concentration. Antibodies for CCAAT-enhancer-binding protein α (C/EBPα, cst-8178), peroxisome proliferator-activated receptor γ (PPARγ, cst-2435), fatty acid binding protein 4 (FABP4, cst-3544), perilipin (cst-9349), pAMPKα (cst-2535), total AMPKα (cst-5831), pACC (cst-11818), and total acetyl CoA carboxylase (ACC, cst-3676) were purchased from Cell Signaling (Danvers, MA, USA). Antibody for actin (sc-1616) was purchased from Santa Cruz (Dallas, TX, USA). 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was purchased from Sigma Aldrich (St. Louis, MO, USA). Bovine calf serum (BCS) and fetal bovine serum (FBS) were purchased from GE Healthcare Life Science (Pittsburgh, PA, USA) and Quality Biological Inc (Gaithersburg, MD, USA), respectively. All other chemicals used were obtained from Fisher (Waltham, MA, USA) or VWR (Radnor, PA, USA), unless otherwise specified.
Maintenance and differentiation of 3T3-L1 preadipocytes
The 3T3-L1 cells were maintained with Dulbecco's modified Eagle's medium (DMEM) containing 10% BCS at an atmosphere of 5% CO2 at 37°C, and medium was replaced every 2 days. For experiments, the cells were plated at a concentration of 3 × 105 cells/well in a 60 mm culture dish and grown to 100% confluency. After 2 days postconfluence, 3T3-L1 cells were incubated with differentiation induction medium (DMEM containing 10% FBS, 1 μg/mL insulin, 0.5 μM dexamethasone, and 0.5 mM isobutylmethylxanthine). After 48 h induction, the cells were incubated with maintenance media (DMEM containing 10% FBS and 1 μg/mL insulin) supplemented with different doses of 3,3′-diindolylmethane for indicated days before differentiation was assessed, and cell lysate was extracted.
Determination of fat accumulation in 3T3-L1 adipocytes
After treatment, the cells were washed with ice-cold phosphate buffer saline (PBS) twice, fixed with 10% buffered formalin for 10 min, and washed twice with distilled water. The cells were stained with 0.2% (final concentration) Oil Red O solution for 10 min at room temperature. After washing with distilled water four times, stained cells were visualized and then dissolved in 0.5 mL isopropanol. Absorbance of the isopropanol extract was measured at 500 nm using a microplate reader (BioTek Instruments, Winooski, VT, USA).
Cell viability
To determine the cell viability, MTT assay was performed as previously described. 15 Briefly, MTT was added to each well containing differentiated 3T3-L1 adipocytes and incubated for 3 h at 37°C. The absorbance was measured in an ELISA plate reader (BioTek Instruments).
Western blot
The cells were washed with ice-cold PBS thrice and lysed in radioimmunoprecipitation assay buffer with a mixture of protease inhibitors and phosphatase inhibitors (Sigma Aldrich, St. Louis, MO, USA). The bicinchoninic acid protein assay was used to quantify the protein concentration (Pierce, Rockford, IL, USA), according to manufacturer's instruction. Next, 30 μg of proteins were mixed with 5x loading buffer and denatured at 95°C for 5 min. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was performed to isolate protein in a size dependent manner, followed by transfer to a nitrocellulose transfer membrane. The membrane was blocked using 5% nonfat milk in Tris-buffered saline with Tween 20 (TBS-T) and incubated with the relevant antibodies in 3% bovine serum albumin (BSA) in TBS-T at 4°C overnight. The membrane was then washed and incubated with a secondary antibody conjugated with horseradish peroxidase for 1 h at room temperature. Chemiluminescence was detected with Pierce ECL Western blotting substrate (Thermo Scientific, Rockford, IL, USA), and the image was developed using ChemiDoc MP Imaging System (Bio-Rad, Hercules, CA, USA).
C. elegans culture
C. elegans were cultured in nematode growth media seeded with E. coli OP50 at 25°C according to a standardized protocol. 10 Synchronized worm population was achieved using beaching method as previously described. On the first day of adulthood, nematodes were administrated with vehicle (DMSO) or different doses of 3,3′-diindolylmethane in S-complete media for 3 days with 0.1% final concentration of DMSO for all treatment.
Lipid quantification of C. elegans
Total fat was quantified as previously reported. 13 After treatment with 3,3′-diindolylmethane, nematodes were collected and washed thrice with M9 buffer to get rid of E. coli OP50. Worm pellets were redispersed in 0.05% Tween 20 and sonicated for 3 min before measurement. Triglycerides and protein content were qualified using commercial kits (Infinity™ Triglycerides Reagent, Thermo Scientific, and Bio-Rad DC Protein Assay Kit), according to the manufacturer's instructions. Triglyceride content was normalized with protein concentration.
Determination of worm size and locomotion assay
Worm size and locomotive activity were measured using WormLab Tracking System as previously described. 13 After treatment with 3,3′-diindolylmethane, worms were transferred to tracking plate freshly seeded with E. coli OP50. Nematodes were allowed to adjust to the new environment for 10 min before recording started. Worms were videoed and analyzed for the data of worm length, width, and locomotive activity.
Statistical analyses
The statistical analyses were performed using one-way ANOVA, and multiple comparisons were analyzed by a Tukey's test, where a P < .05 was considered significant. Data are presented as mean ± SD.
Results
3,3′-diindolylmethane inhibited fat accumulation in 3T3-L1 adipocytes
First, we determined the effects of 3,3′-diindolylmethane on fat accumulation. As shown in Figure 1B, Oil Red O staining as representative of fat accumulation was significantly reduced when cells were treated with 50–100 μM 3,3′-diindolylmethane compared to control. Next, we used the same concentration of 3,3′-diindolylmethane to determine any effect of those doses on cell viability using MTT-based assay in 3T3-L1 adipocytes. The results indicate that 3,3′-diindolylmethane, up to 100 μM, did not affect cell viability (Fig. 1C).
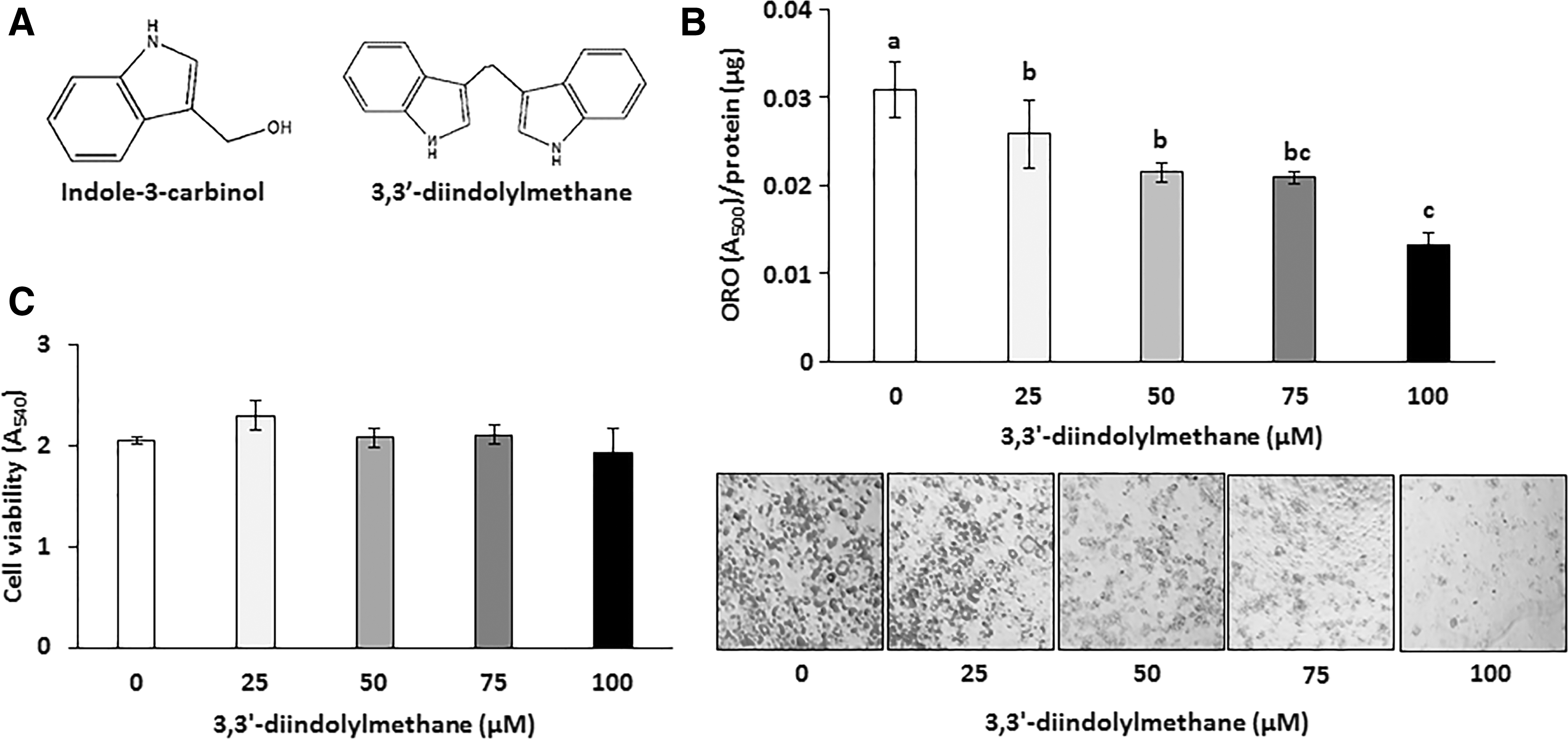
Effects of 3,3′-diindolylmethane on accumulation of triglyceride and cytotoxicity in differentiated 3T3-L1 adipocytes.
3,3′-diindolylmethane suppressed adipogenic transcription factors and their target proteins
Differentiation of preadipocytes into adipocytes is tightly regulated by a sequential activation of several transcriptional factors, including PPARγ and C/EBPα. 16 Moreover, FABP4 and perilipin are downstream targets of PPARγ and C/EBPα and also known to be involved in triglyceride biosynthesis in adipocytes. 17,18 Thus, we further tested 3,3′-diindolylmethane on those factors in 3T3-L1 cells (Fig. 2A–D). Treatment with 3,3′-diindolylmethane significantly decreased the expression of PPARγ and C/EBPα, as well as FABP4 and perilipin, particularly at 75 μM or higher concentrations, compared to respective controls.
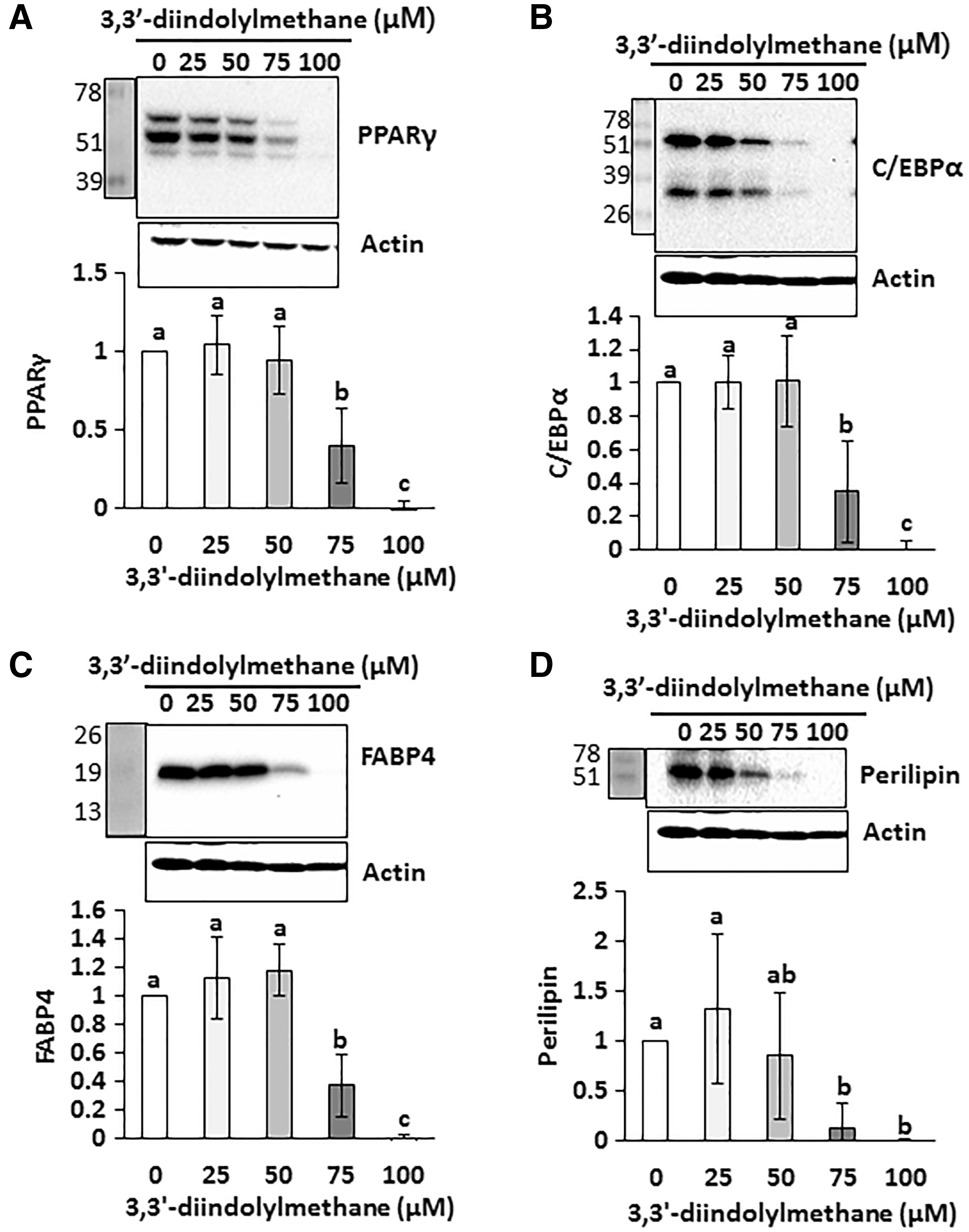
Effects of 3,3′-diindolylmethane on adipogenic transcription factors and their lipogenic target proteins in differentiated 3T3-L1 adipocytes. 3T3-L1 preadipocytes were differentiated in the presence of different concentrations of 3,3′-diindolylmethane for 6 days. PPARγ
3,3′-diindolylmethane increased phosphorylation of AMPKα and ACC
Since AMPKα is an important energy sensor, as well as a regulator of fat cell differentiation, we tested if 3,3′-diindolylmethane affects active phosphorylation of AMPKα in differentiated 3T3-L1 cells. The result indicates that the amount of phosphorylated AMPKα increased in the cells treated with 75 and 100 μM of 3,3′-diindolylmethane (Fig. 3A). Next, we tested phospho-ACC (inactive form), which is a downstream target of AMPKα. As shown in Figure 3B, phosphorylation of ACC was significantly increased in the presence of 75 and 100 μM 3,3′-diindolylmethane compared to control.
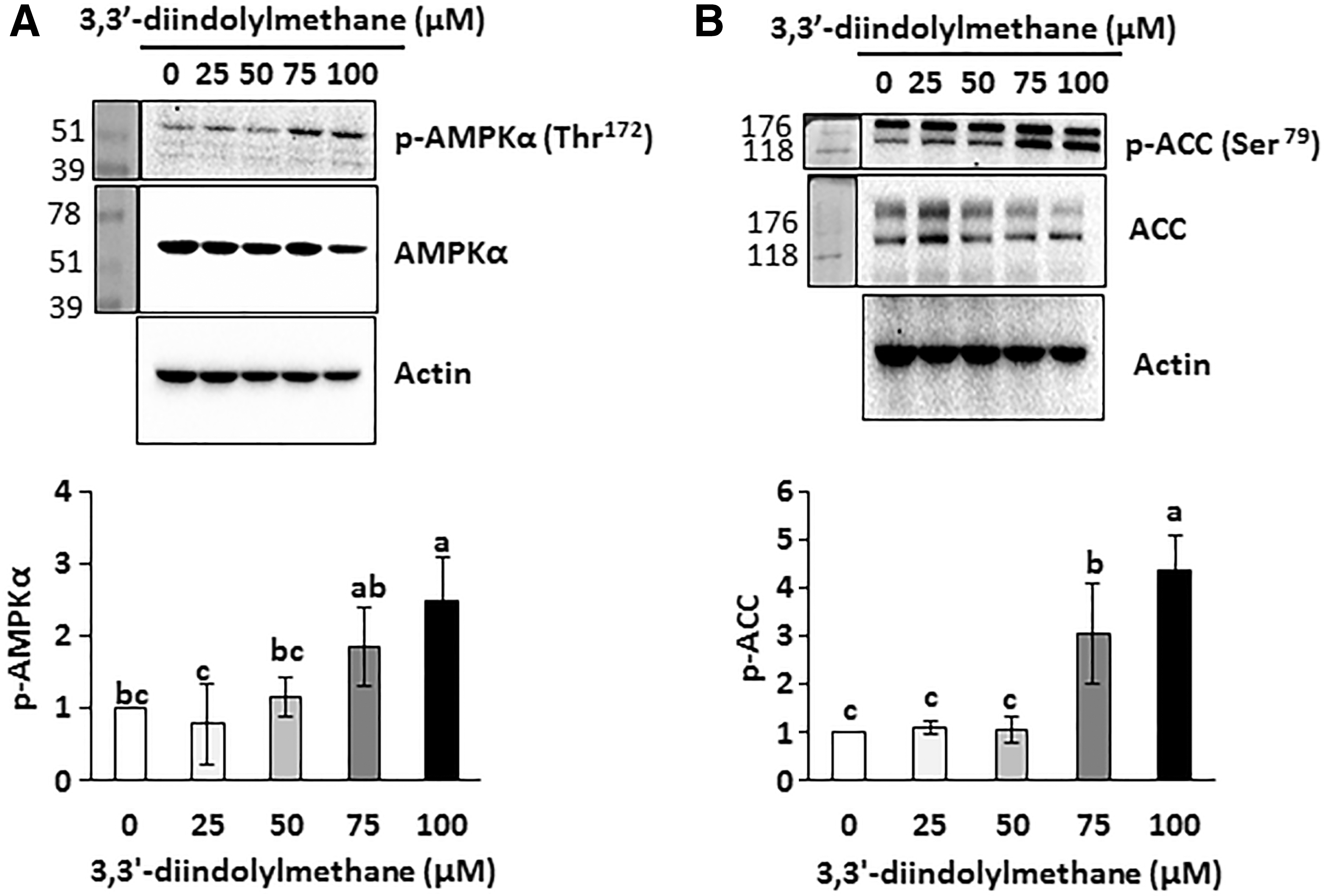
Effects of 3,3′-diindolylmethane on phosphorylation of AMPKα and ACC in differentiated 3T3-L1 adipocytes. 3T3-L1 preadipocytes were differentiated in the presence of different concentrations of 3,3′-diindolylmethane for 6 days. Phospho- and total AMPKα
3,3′-diindolylmethane decreased accumulation of triglycerides in C. elegans
Based on the above in vitro results, we further evaluated the effects of 3,3′-diindolylmethane on accumulation of triglycerides using in vivo C. elegans. As shown in Figure 4A, 3,3′-diindolylmethane treatment at 100 μM significantly inhibited accumulation of triglycerides compared to control in wild-type C. elegans. We further confirmed that treatment with 3,3′-diindolylmethane led to 2.7% reduction in worm length and 13% reduction in 100 μM over control, respectively (Table 1), which may be due to reduced overall triglyceride accumulation. However, no effects of 3,3′-diindolylmethane were observed on locomotive activity over control, suggesting that behaviors were not altered.
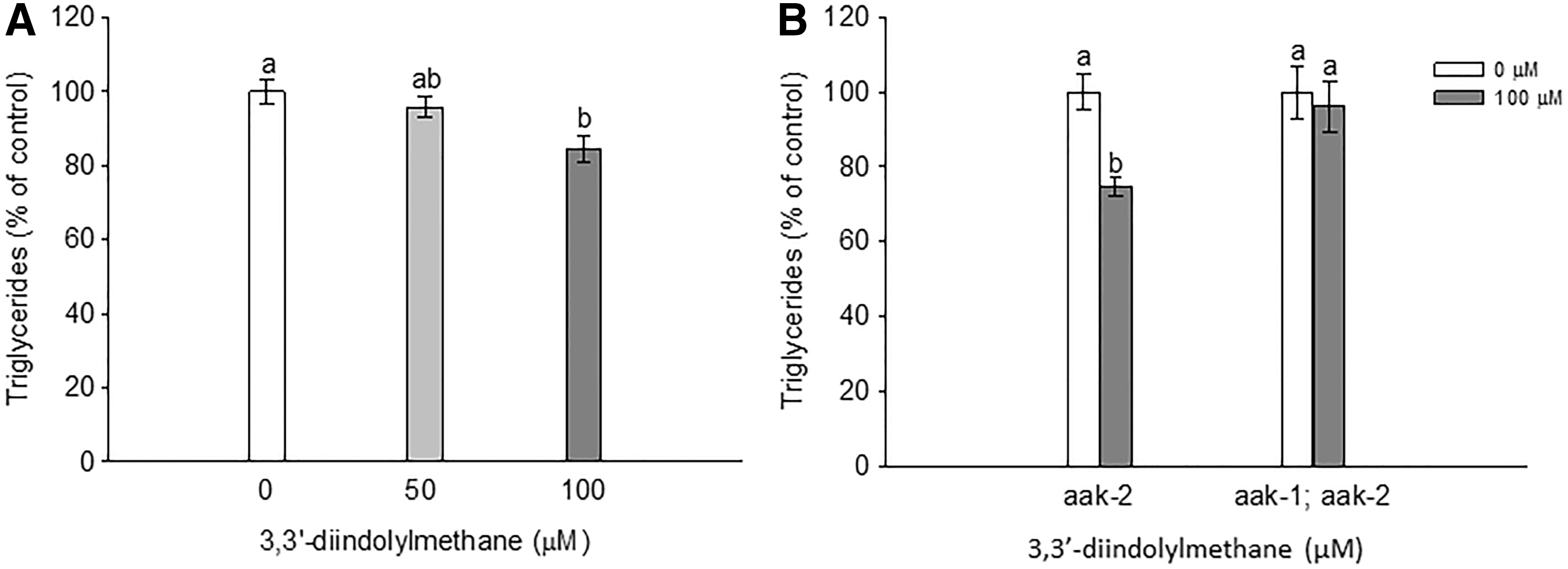
Effects of 3,3′-diindolylmethane on accumulation of triglyceride in Caenorhabditis elegans. Total triacylglyceride contents in
Caenorhabditis elegans were treated with 3,3′-diindolylmethane on the first day of adulthood for 3 days. Results are mean ± SD (n = 63–109). Means with different letters are significantly different at P < .05.
Based on the observation that 3,3′-diindolylmethane activated AMPKα in 3T3-L1 cells, the role of AMPKα in 3,3′-diindolylmethane-induced fat reduction in C. elegans was investigated. There are two C. elegans orthologs encoding the catalytic alpha subunit of AMPK: aak-1 and aak-2. In this study, aak-2 single mutant and aak-1; aak-2 double mutant were used to evaluate the effect of 3,3′-diindolylmethane on accumulation of triglycerides. 3,3′-diindolylmethane had no effects on triglyceride content in aak1; aak-2 double mutant, but had significant reduction on triglyceride accumulation in aak-2 single mutants (Fig. 4B). These results suggest that reduced triglyceride accumulation caused by 3,3′-diindolylmethane was dependent on AMPKα, particularly aak-1, in this model.
Discussion
Cruciferous vegetables contain a number of bioactive compounds, including polyphenols and flavonoids. Indole-3-carbinol is one of the most abundant polyphenol compounds and possesses diverse and beneficial health-related activities, such as anti-inflammatory and anticancer properties. 19 Previously, our research group reported that 3,3′-diindolylmethane, a major in vivo metabolite of indole-3-carbinol, increased apoptosis and growth arrest of colon cancer cells, 20,21 and 3,3′-diindolylmethane shows synergistic anticancer activity with other dietary compounds. 15 Recently there were several publications on indole-3-carbinol and prevention of obesity. 4 –6 However, no study has examined the association between 3,3′-diindolylmethane and adipogenesis. Thus, the current study was designed to investigate the effects of 3,3′-diindolylmethane on lipid accumulation and this is the first report that 3,3′-diindolylmethane possesses significant antiadipogenic activity in differentiated 3T3-L1 adipocytes and C. elegans.
The current results indicate that 3,3′-diindolylmethane suppressed expression of differentiation transcription factors, including PPARγ and C/EBPα, in differentiated 3T3-L1 adipocytes. The transcription factor PPARγ is expressed at early stages of adipocyte differentiation and regulates adipogenesis. The differentiation of exponentially growing 3T3-L1 cells into adipocytes is potentiated by insulin, 22 and PI3K/Akt is a target of 3,3′-diindolylmethane. 23 Thus, we speculate that antiadipogenic activity of 3,3′-diindolylmethane may, in part, be due to inhibition of the signaling pathway from insulin to PI3K/Akt during differentiation of preadipocytes, although this may need to be determined further.
FABP4 is the major form of lipid carrier protein in adipocytes 24 and its expression is tightly regulated by PPARγ. 25,26 Perilipin is highly expressed in adipocytes and controls lipid accumulation. 27 Our data indicate that 3,3′-diindolylmethane may prevent fat accumulation through regulation of lipid carrier and droplet associated proteins.
AMPKα is a serine/threonine protein kinase that plays a significant role in maintaining energy metabolism and regulation of body weight. Aberrant expression and pharmacological inhibition of AMPKα are associated with development of adipogenesis/lipogenesis, obesity, and related metabolic diseases. 28 On the contrary, treatment of AICAR (5-aminoimidazole-4-carboxamide-1-beta-D-ribofuranoside), AMPK activator, reduces obesity and insulin resistance. 29 Therefore, this kinase has received considerable interest as a promising preventive and therapeutic target against obesity and obesity-related morbidities, including insulin resistance and atherosclerosis. In this study, we observed that 3,3′-diindolylmethane activates AMPKα, which subsequently phosphorylates ACC (pACC is inactive form of enzyme), at Ser79 on ACC1, the dominant isoform in white adipose tissues, 30 or the orthologous site Ser212 on ACC2. Thus, the current results suggest that 3,3′-diindolylmethane reduces lipogenesis by inactivation of ACC through AMPKα-dependent mechanism. 31 This was further confirmed in in vivo study, which suggests the antiobesity activity of 3,3′-diindolylmethane in C. elegans. There are two AMPKα (aak-1 and aak-2) catalytic subunits in C. elegans with functional differences. To investigate if antiobesity activity of 3,3′-diindolylmethane is isomer specific, we used C. elegans mutants having a single aak-2 mutation or aak-1/aak-2 double mutations. The results indicate that aak-1 is involved in the antiobesity effect of 3,3′-diindolylmethane (Fig. 4B). It has been known that aak-1 shows broader expression pattern than aak-2, which is more specific for energy metabolism, insulin signaling, and life span regulation. 32,33 We do not know if aak-1 also mediates antiadipogenic activity of 3,3′-diindolylmethane in fully differentiated adipocytes. Further studies using specific inhibition of each isoform in adipocytes are required to address this question.
AMPK-dependent decrease of triglyceride accumulation in 3,3′-diindolylmethane-treated C. elegans supports a significant role of anabolic pathway in lipid metabolism of C. elegans. However, with limited data we do not exclude the possibility that decreased fat accumulation is associated with increased lipolysis and subsequent fatty acid oxidation, or mechanism may be different between C. elegans and 3T3-L1 adipocytes. Thus, further research is required to address this speculation.
We do not know the exact physiological level of 3,3′-diindolylmethane after consumption of dietary sources. Recently, Rajoria et al. measured serum concentration of 3,3′-diindolylmethane from subjects who took 3,3′-diindolylmethane tablets at a dose of 300 mg/day for 14 days, and the concentration was normalized per milligram of creatinine. 34 Although the effective dose (50–100 μM) we observed in the present study seems to be higher than physiological level, the relevance of in vitro doses to in vivo environment should be considered with a couple of factors such as local concentration, bioavailability, and potential further metabolites.
In conclusion, the current study demonstrated that 3,3′-diindolylmethane, a major metabolite of indole-3-carbinol, suppresses adipogenesis, in part, through AMPKα-dependent mechanisms in 3T3-L1 adipocytes and C. elegans.
Footnotes
Acknowledgments
In vitro studies were supported by Maryland Agricultural Experiment Station grant to S-H Lee from the University of Maryland. C. elegans work was supported by the National Institute of Food and Agriculture, U.S. Department of Agriculture, the Massachusetts Agricultural Experiment Station, and the Department of Food Science, the University of Massachusetts Amherst, under project number MAS00450. All the strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440). The authors thank Ms. Jayne M. Storkson for help preparing this article.
Author Disclosure Statement
No competing financial interests exist.