
Abstract
Our previous results suggest that the Rosa rugosa Thunb. (family Rosaceae) alleviates endurance exercise-induced stress by decreasing oxidative stress levels. This study aimed to screen and identify the physiological antistress effects of an extract of R. rugosa (RO) on sleep deprivation–induced anxiety-like behavior and cognitive tests (in vivo) and tested for hippocampal CORT and monoamine levels (ex vivo), corticosterone (CORT)-induced injury, N-methyl-
Introduction
S
The monoamine neurotransmitter 5-hydroxytryptamine (5-HT, serotonin) is derived from tryptophan and regulates a multitude of behavior and physiologies, including depression, anxiety, obesity, social phobia, schizophrenia, sleep and wakefulness, cognition, sexual behavior, eating disorders, aggression, impulsivity, neurodevelopment, circadian rhythms, body temperature, and neuroendocrine function. 14 –16 The 5-HT receptors are categorized into seven classes, 5-HT1–7. Of these, 5-HT6, a G-protein coupled receptor, is located in postsynaptic neurons and has been localized in the terminal regions involved in the regulation of sleep and wakefulness, including the cerebral cortex, basal forebrain, limbic system (hippocampus, amygdala), striatum, nucleus accumbens, thalamus, hypothalamus (anterior and lateral area), and brain stem (raphe nuclei) of the rat. 17 –22 The primary signal transduction pathway of 5-HT6 involves the stimulation of adenylate cyclase activity and production of cAMP. 23
Rosa rugosa Thunb. (Rosaceae; common name Haedanghwa in Korea) is a fragrant medicinal plant found in East Asia. Previous investigations have isolated hydrolyzable tannins, catechin derivatives, flavonoids, monoterpenes, sesquiterpenes, and triterpenes from this species. 24 The R. rugosa plant has traditionally been used as a treatment for several disorders, including diabetes, chronic inflammatory diseases, pain, HIV-1 infection, and hypertension, and for its anticancer and antioxidant effects. 25 –32 Recent studies have reported the efficacy of R. rugosa for the treatment of exercise-induced stress by decreasing oxidative stress levels. 33 However, few studies have investigated its antistress effects, especially those induced by sleep deprivation. Therefore, the present study was designed to study the in vivo antistress effect of an R. rugosa extract in a sleep-deprived (SD) animal model and to investigate the underlying mechanisms in vitro.
Materials and Methods
Chemicals and reagents
The stems and leaves of R. rugosa Thunb. (Yeonggwang, South Korea) were used for this experiment. H2O2, CORT, and caffeine were purchased from Sigma-Aldrich, and GABA, NMDA, serotonin, and antagonists were purchased from Tocris. In vitro reagents, including media, etc., were obtained from Gibco. All other reagents were of analytical grade.
Preparation of the R. rugosa extract (RO)
The stems and leaves of R. rugosa were extracted with distilled water at 100°C, 20% ethanol at 95°C, and 80% ethanol at 75°C for 4 h and concentrated by freeze-drying. The resulting powder extracts (6.4%, water extract; 4.1%, 20% ethanol extract; and 4.9%, 80% ethanol extract) were dissolved in saline (0.9% NaCl) for in vivo experiments and in distilled water for in vitro experiments.
The RO extracts were fractionated on HP-20 resin columns using the following H2O:MeOH (v/v) elution series: 100:0 (32.53%), 20:80 (3.18%), 50:50 (19.32%), 80:20 (10.13%), and 0:100 (10.13%). Each fraction was evaporated to dryness and dissolved as above for experiments.
Animals
Sprague-Dawley rats (male, 150–170 g) were obtained from Samtako Co. Ltd., Osan, Korea. All animals were housed in a light-controlled room (lights on from 8:00 AM to 8:00 PM) at a temperature of 22 ± 2°C and a relative humidity of 50% ± 5% and provided with food ad libitum.
All experimental procedures were conducted in accordance with the guidelines for the care of experimental animals, as approved by the Institutional Animal Care and Use Committee of Jeollanamdo Institute for Natural Resources Research.
Rats were randomly assigned to 1 of 10 groups (n = 5–7 per group): control (non-SD rats), SD group, SD group treated with RO 50, 100, and 200 mg/kg of B.W., SD group treated with PCTL (caffeine 10 mg/kg), non-SD group treated with RO 50, 100, and 200 mg/kg, and non-SD group treated with PCTL (caffeine 10 mg/kg). For these experiments, all rats were placed individually in a plastic chamber with one small or large platform surrounded by water, as described previously. 34 The RO and caffeine treatments were administered orally.
Sleep deprivation procedure
The platform method was used for the SD animal model. Briefly, a cage (21 × 25 × 27 cm) was filled with water and a small flower pot–shaped platform (6 Φ × 10 cm high) was placed within 1 cm of the platform where the rat stands and was changed every day for drinking. Rats of the non-SD groups were placed in a chamber with large platforms (15 cm diameter), which were large enough for the rats to sleep on, but not large enough for them to walk around on, and were also surrounded by water. Physiological factors, including body weight and survival, were checked every day, and the drowsiness score (5 min) was analyzed 72 h after an SD day.
The effect of RO on physiological and behavioral changes caused sleep deprivation stress, which was evaluated in separate experiments.
Cortisol content analysis
Serum was isolated following centrifugation at 3000 g for 20 min. Serum samples were stored at −20°C until assayed. Cortisol levels were determined by a competitive ELISA Kit (Calbiotech) following the manufacturer's protocol. The cortisol MAb–coated microplate was read at 450 nm, and the cortisol concentrations were calculated from standard curves.
Serotonin content analysis
Serotonin in serum was measured by ELISA Kit (reference range: 10.2–2500 ng/mL) provided by Abnova. The intra- and interassay coefficient of variation was 12.6% and 10.4%, respectively. Concentration of serotonin was calculated from the standard curves. The sensitivity of the serotonin was 6.2 ng/mL.
Dopamine content analysis
Dopamine in serum was measured by ELISA Kit (reference range: 0.5–80 ng/mL) provided by Abnova. The intra-assay coefficient of variation was 21.5%. Concentration of dopamine was calculated from the standard curves. The sensitivity of the dopamine was 3.3 pg/mL.
Morris water maze test
The water maze test was performed according to standard methods with some modifications. 35 A circular pool (diameter: 180 cm, height: 75 cm) was filled with water to a depth of 30 ± 1 cm and was made opaque by addition of nonfat milk. The maze was divided into four equal quadrants on the monitoring screen of a computer. The transparent escape platform was placed in one of the four quadrants (target quadrant) and submerged ∼1 cm below the surface of the water.
The first day of the experiment was a swim training day, during which the rats swam for 60 sec in the absence of the platform. On each day following the first training day, the rats were given four sessions, with an intertrial interval of 5 min. The starting point changed for each session, but the location of the platform was fixed during the entire test period. The time from being placed in the pool to reaching the platform was measured.
On the day after the last training session, each rat was subjected to a probe trial (90 sec), in which no platform was present. The time spent in the target quadrant (the quadrant in which the platform had previously been located) was taken as a measure of spatial memory retention. The maze performance was recorded by a video camera suspended above the maze and analyzed using Smart video tracking system (Panlab).
Rotarod test
This experiment was performed on a rotarod task (JD-A-07; Jeung Do Bio & Plant) for measurement of motor balance and coordination. Rats were placed on a horizontal rotating rod (diameter, 8 cm; rotation speed, 15 rpm) and were left on the rod for 5 min or until they fell off. Falling off the rod activated a switch that automatically stopped a timer. Five rats separated by large disks were tested simultaneously. On the testing day, each rat was submitted to three trials with an intertrial interval of 10 min.
Passive avoidance test
The passive avoidance test was performed as previously described, 36 with minor modifications. The rats were tested for memory retention deficits using a passive avoidance apparatus (Iwoo Scientific Co) consisting of a two-compartment dark/light shuttle box with a guillotine door separating the compartments. The dark compartment had a stainless steel shock grid floor. During the acquisition trial, each rat was placed in the light chamber. After a 60-sec habituation period, the guillotine door was opened, and the latency for the animal to enter the dark chamber was recorded. This was termed the initial latency; rats with an initial latency of greater than 60 sec were excluded from further experiments. Immediately after the rat had entered the dark chamber, the guillotine door was closed and an electric foot shock (75 V, 0.2 mA, 50 Hz) was delivered through the floor grid with a stimulator for 3 sec. Five seconds later, the rat was removed from the dark chamber and returned to its home cage. Twenty-four hours later, this test was repeated and the latency to enter the dark chamber was recorded. The latency in this test was termed the retention latency.
Neuroprotection effects
Human neuroblastoma SH-SY5Y cells were used to confirm the neuroprotective effect against H2O2 or corticosterone (CORT). The cells were maintained in MEM medium supplemented with 10% FBS, 1% penicillin, and streptomycin in a humid atmosphere of 5% CO2 and 95% air at 37°C. SH-SY5Y cells were plated in 96-well plates and then pretreated with various nontoxic concentrations of RO extract (1, 3, 10, 30, and 100 μg/mL) for 24 h. This was followed by exposure to H2O2 (100 μM) or CORT (1 mM) in the presence of the same concentrations of RO extract for another 24 h. Cell viability was confirmed by the MTT assay. 37
GABAB/NMDA-induced intracellular Ca2+ imaging
GABAB/Gqi5-transfected CHO cells were prepared according to Wood et al. as previously described. 38 Cultured rat hippocampal neurons were prepared for the NMDA-related assay using a technique modified method as previously reported. 39
The acetoxymethyl-ester form of fura-2 (fura-2/AM; Molecular Probes) was used as the fluorescent Ca2+ indicator. Cells were incubated for 40–60 min at room temperature with 5 μM fura-2/AM and 0.001% Pluronic F-127 in Mg-free HEPES-buffered solution composed of (in mM): 150 NaCl, 5 KCl, 2 CaCl2, 10 HEPES, 10 glucose, and pH adjusted to 7.4 with NaOH. Cells were illuminated using a xenon arc lamp, and excitation wavelengths (340 and 380 nm) were selected by a computer-controlled filter wheel (Sutter Instruments).
Data were acquired every 2 sec and a shutter was interposed in the light path between exposures to protect cells from phototoxicity. Emitter fluorescence light was reflected through a 515 nm long-pass filter to a frame transfer cooled CCD camera and ratios of emitted fluorescence were calculated using a digital fluorescence analyzer. All imaging data were collected and analyzed using MetaMorph software (Molecular Devices).
cAMP inhibition
The γ-irradiated recombinant 1321N1 cells (human brain astrocytoma) expressing the human 5-HT6 receptor (Perkin Elmer) were cultured in DMEM containing 10% FBS and plated on 24-well plates overnight. Cells were washed in PBS buffer containing 0.5 mM phosphodiesterase inhibitors IBMS (3-isobutyl-1-methylxanthine) and Ro 20–1724 (0.1 mM) and underwent three different stimulation treatments (pretreatment with RO; treatment with RO and 5-HT simultaneously; and treatment with only RO) for 15 or 30 min. Intracellular cAMP levels were measured using a parameter™ Cyclic AMP Assay Kit (R&D Systems) according to the manufacturer's instructions.
Western blot analysis
The 5-HT6R-transfected 1321N1 cells were kept in serum-free DMEM for 24 h, treated with 5-HT or pretreated for 15 min with RO, and then lyzed in lysis buffer (PRO-PREP™ Protein Extraction Solution, iNtRON Biotechnology). After sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), the proteins were transferred to a polyvinylidene difluoride membrane (Millipore), and the membrane was blocked with Tris-buffered saline containing 5% skim milk and 0.1% Tween 20 for 1 h at room temperature. The membranes were then incubated with the primary antibody anti-ERK1/2 (1:1000; Cell Signaling), anti-phospho ERK1/2 (1:1000; Cell Signaling), anti-MEK1/2 (1:1000; Cell Signaling), anti-phospho-MEK 1/2 (1:1000; Cell Signaling), or β-actin (Sigma-Aldrich) overnight at 4°C. After three washes, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (anti-mouse or anti-rabbit 1:1000; Cell Signaling) for 1.5 h at room temperature and visualized using the Immobilon Western Chemiluminescent HRP Substrate (Millipore).
Statistical analyses
The results are expressed as the means ± standard errors of the means (SEM) for the number of animals in each group. The differences between the data obtained from “test” animal groups and the data obtained from untreated animal groups were subjected to one-way analysis of variance (ANOVA; 95% confidence interval), followed by Dunnett's test. We evaluated the performance of the preliminary F-test to confirm the homogeneity of within-group variance. Differences were considered significant at error probabilities smaller than 0.05. All graphing and statistical analyses were performed using GraphPad Prism 5 for Windows (GraphPad Software).
Results
Improvement of sleep deprivation-induced physiological and biochemical deficits by RO extract
We investigated the protective effect of the RO extract against sleep deprivation stress by measuring physiological (survival, bodyweight, and drowsiness scores) and biochemical (cortisol, serotonin, and dopamine contents) factors in the rats.
The survival rate of the SD group decreased from day 6, with the deaths of almost all rats occurring by day 15–16 (Fig. 1A). The RO-treated groups showed higher survival rates compared to the SD group. The RO200/SD group had an 85% survival (n = 6) at day 15. Caffeine had no effect on survival rate. The body weight of the CTL group rats gradually increased as time progressed, but sleep deprivation-induced stress caused a significant weight loss regardless of treatment (Fig. 1B).
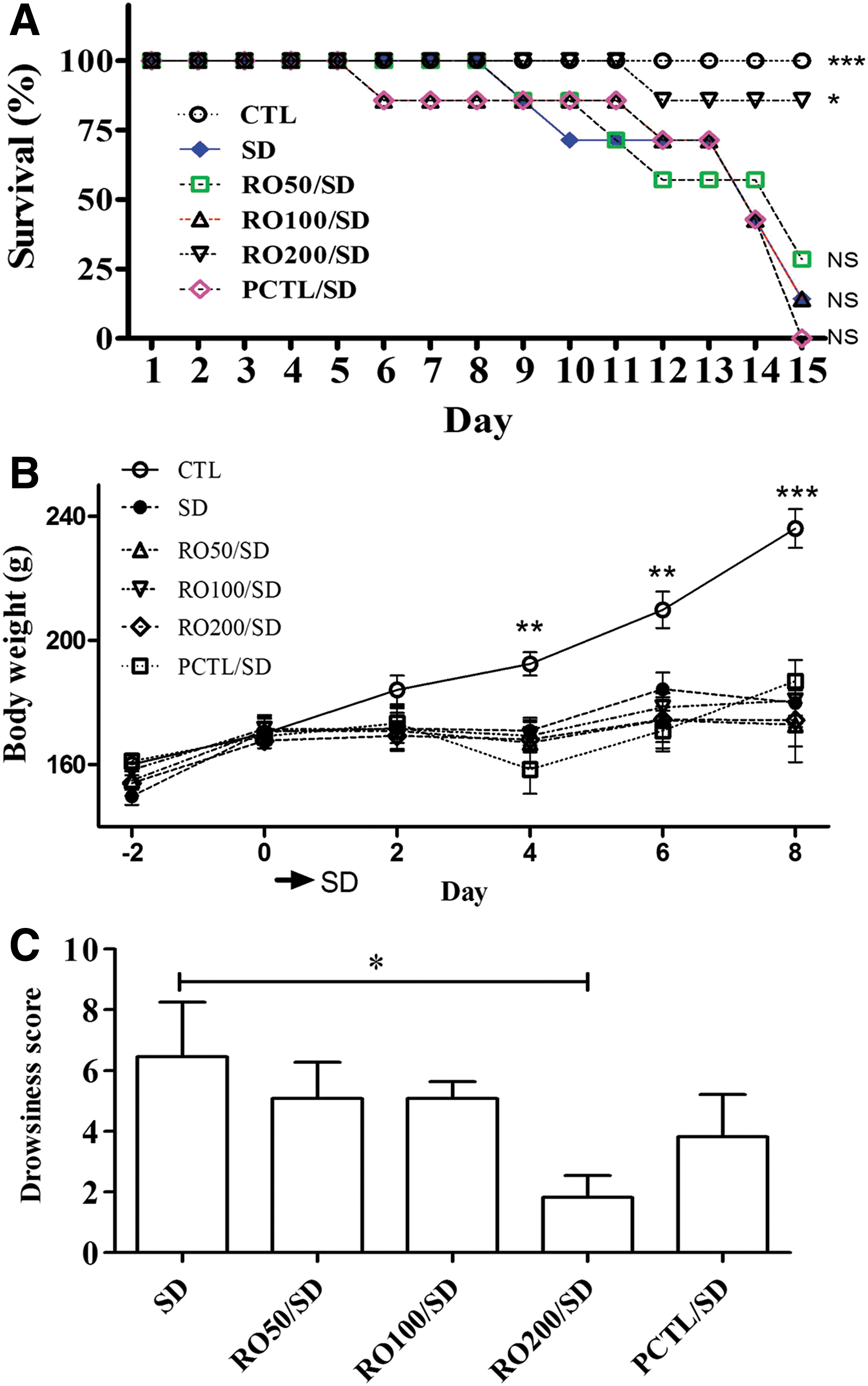
Effects of a Rosa rugosa (RO) extract on sleep deprivation-induced impairment of physiological factors in rats.
The drowsiness score of the SD groups was decreased in a concentration-dependent manner by administration of RO. The group receiving RO 200 mg/kg showed a significant decrease (P < .05) compared to the SD group.
Rat blood sera were used to confirm the cortisol contents. Figure 2A shows that the SD group had the highest serum cortisol concentration (113.6 ± 30.59 μg/mL) and it significantly differed from that of the RO200/SD group (35.16 ± 5.52 μg/mL). The levels for the other treatments were also lower than the SD group (naive, 75.89 ± 15.93 μg/mL; CTL, 56.03 ± 17.55 μg/mL; RO50/SD, 73.54 ± 54.26 μg/mL; RO100/SD, 48.20 ± 38.34 μg/mL; and PCTL/SD, 51.12 ± 19.73 μg/mL).
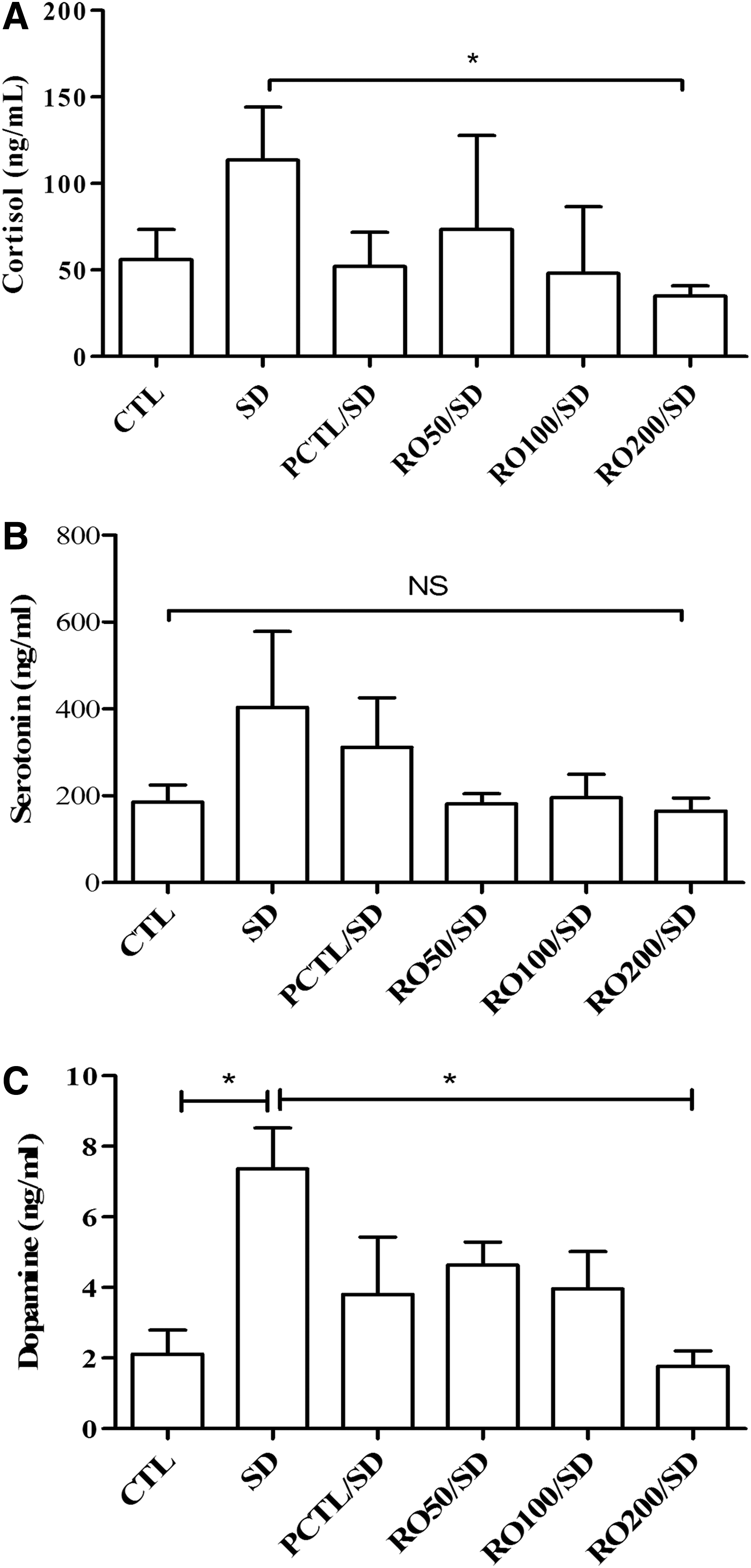
Effects of an R. rugosa (RO) extract on sleep deprivation-induced impairment of biochemical factors in rats.
Serotonin levels, shown in Figure 2B, were increased by sleep deprivation stress and diminished by the PCTL and RO treatments, but did not differ significantly among groups (CTL, 185.9 ± 38.83 μg/mL; SD, 403.7 ± 174.9 μg/mL; RO50/SD, 181.4 ± 23.34 μg/mL; RO100/SD, 195.6 ± 53.96 μg/mL; RO200/SD, 165.0 ± 29.96 μg/mL; and PCTL/SD, 311.3 ± 114.2 μg/mL).
Dopamine concentrations (Fig. 2C) were also increased by sleep deprivation stress (CTL, 2.12 ± 0.68 μg/mL; SD, 7.36 ± 1.16 μg/mL; P < .05), and the PCTL/SD and RO treatment ameliorated the increased dopamine level, especially the RO200/SD group decreased dopamine significantly (P < .05).
Caffeine treatment, as the positive control (PCTL), tended to decrease the drowsiness score and biochemical contents in response to sleep deprivation-induced stress, but had no effect on survival rate.
Effect of RO on impairment of memory acquisition and retention determined by the Morris water maze test
The effects of RO on spatial memory were investigated using the Morris water maze test (Fig. 3). Figure 3A shows the escape latencies for each group in trials 1–3 on day 3. The CTL group rapidly learned the location of the platform, as demonstrated by the short escape latencies. The escape latencies decreased from the first trial to third trial and from the first day to the third day, due to normal learning. However, despite learning for 3 days, the escape latencies of the SD group did not decrease (77.49 ± 24.92 sec) compared to the level observed in the CTL group (32.74 ± 7.47 sec). In contrast, the escape latencies of the RO50/SD (26.73 ± 6.52 sec), RO100/SD (18.34 ± 2.62 sec), RO200/SD (22.96 ± 5.17 sec), and PCTL/SD (20.10 ± 7.32 sec) groups showed decreases over the 3 days of testing.

Effects of an R. rugosa (RO) extract on sleep deprivation-induced impairment of memory acquisition and retention in rats performing the Morris water maze test.
The results of the spatial probe trials are shown in Figure 3B–D, which shows a comparison of the first and third latencies of each day, and confirmed the statistical significance. In the first trial, the CTL, RO200/SD, and PCTL/SD groups showed significantly decreased escape latencies compared to the SD group on day 2, and the latencies in all groups, except for the SD group, were decreased on day 3.
Effect of the RO extract on sleep deprivation-induced behavioral deficits
In the rotarod test, the SD group showed a significant reduction in time spent on the rod (65.67 ± 30.79 sec) compared to the CTL group (228.50 ± 48.19 sec) and the RO-treated group (RO200/SD, 227.44 ± 23.96 sec), indicating better locomotor activity adaptation and coordination in the control and RO-treated groups (Fig. 4A).
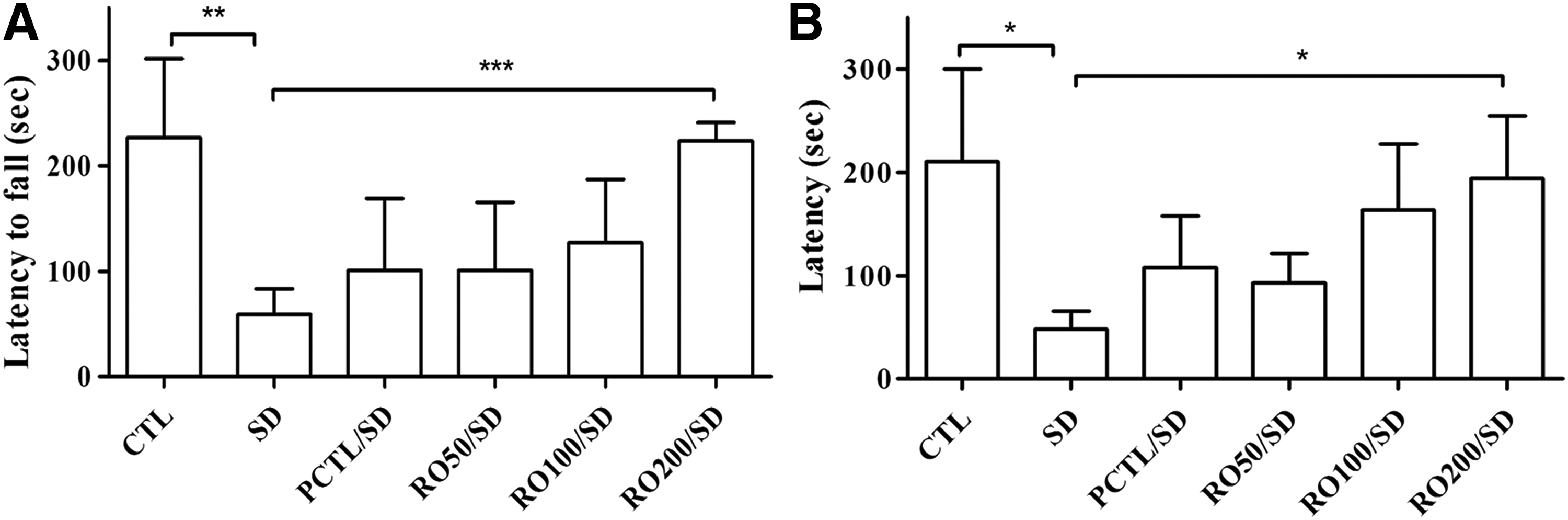
Effects of an R. rugosa (RO) extract on sleep deprivation-induced behavioral deficits in rats.
The effects of the RO extract treatment on the retention latencies, as revealed by the passive avoidance test, are shown in Figure 4B. The SD group showed a decreased retention latency (48.60 ± 16.74 sec) compared to the CTL group (163.50 ± 78.83 sec). The reduced latency induced by sleep deprivation was significantly ameliorated by RO extract administration (RO200/SD mg/kg, 168.20 ± 53.78 sec). The retention latency reduction induced by sleep deprivation was also alleviated by caffeine administration (94.60 ± 40.96 sec). The non-SD groups that administered RO extract (168.20 ± 53.78 sec) or caffeine (190.40 ± 67.13 sec) showed equal escape latencies that were significantly different from the control group values.
Neuroprotective effect of the RO extract against H2O2- and corticosterone-induced damage
SH-SY5Y cells were exposed to H2O2 (100 μM) or CORT (1 mM) for 24 h and cell survival was then assessed by the MTT assay. As shown in Figure 5A, 100 μM of H2O2 induced significant decreases in cell survival to 71.57% ± 0.87% viability (mean ± SEM, n = 5) compared to CTL cells. The RO extract treatment (1, 3, 10, 30, and 100 μg/mL) suppressed the H2O2-induced damage to the cells and resulted in 74.70% ± 2.03%, 73.52% ± 1.96%, 76.76% ± 2.24%, 75.99% ± 0.66%, and 80.10% ± 0.54% viability, respectively.

Neuroprotective effects of R. rugosa (RO) extract on H2O2- or CORT-induced apoptosis in SH-SY5Y cells. Cells were pretreated with various concentrations of the RO extract, followed by 100 μM H2O2 or 1 mM CORT challenge for 24 h.
As shown in Figure 5B, 1 mM CORT treatment decreased the cell viability to 71.86% ± 1.20%. Viability was restored by treatment with RO extract (1, 3, 10, 30, 100 μg/mL) to 72.50% ± 1.61%, 77.11% ± 3.33%, 78.96% ± 1.18%, 83.39% ± 3.10% and 82.06% ± 0.89%, respectively.
The H2O2 and CORT-induced cell stress was significantly decreased by pretreatment with RO extract at 10, 30, and 100 μg/mL. Treatment with RO alone at each of these concentrations did not cause any significant cytotoxicity (data not shown).
Suppression of the NMDA/GABA-induced [Ca2+]i increase by the RO extract
The mechanism underlying the stress-relieving effect of RO was examined by recording the NMDA or GABA-induced [Ca2+] i increase using fura-2-based digital imaging techniques.
Most of the cultured hippocampal cells showed a rapid increase in [Ca2+] i in the Mg2+-free and 1 μM glycine-containing recording solution following the acute application of NMDA (100 μM, for a 10 sec duration). This NMDA-induced [Ca2+] i increase could be reproduced by repeated applications of NMDA at 4–5 min intervals up to 1 h and was not significantly diminished by multiple applications of NMDA. We examined the effects of RO extract on NMDA-induced [Ca2+] i increase under these conditions. The cells were pretreated with various concentrations of RO extract for 1 min, followed by NMDA. The increase in [Ca2+] i was calculated from the difference between the peak and basal values of [Ca2+] i .
The effect of the RO extract on the GABA receptor was confirmed using GABA and a Gqi5-transfected CHO cell (Fig. 6A, B). The influx of Ca2+ into the cells was strongly inhibited by the GABA receptor antagonist, saclofen (10 μM). The RO extract showed weak inhibitory effects on intracellular Ca2+: 10.97% ± 2.35% (1 μg/mL), 5.88% (3 μg/mL), 7.33% ± 6.36% (10 μg/mL), 15.23% ± 5.86% (30 μg/mL), and 40.00% ± 5.65% (100 μg/mL).
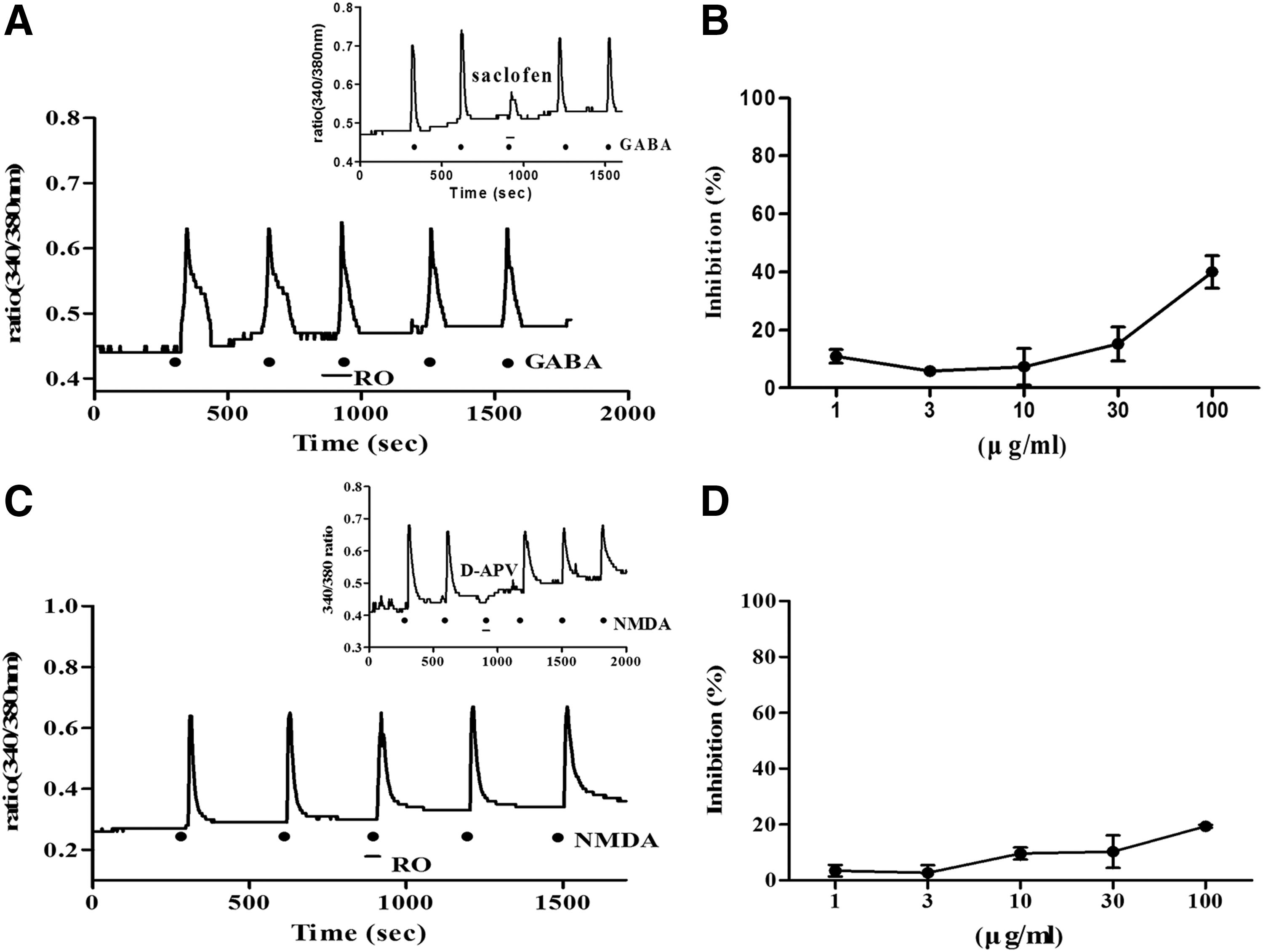
Effect of an R. rugosa (RO) extract on GABAB receptor activity in Gqi5-transfected CHO cells and NMDA receptor activity in hippocampal cells.
Hippocampal cells were used to analyze the changes in intracellular Ca2+ caused by the action of the RO extract on the NMDA receptor (Fig. 6C, D). The NMDA receptor antagonist D-APV (250 μM) completely inhibited [Ca2+] i . changes, whereas the inhibitory effects of RO were 3.46% ± 2.06% (1 μg/mL), 2.72% ± 2.72% (3 μg/mL), 9.64% ± 2.16% (10 μg/mL), 10.28% ± 5.86% (30 μg/mL), and 19.40% ± 0.50% (100 μg/mL).
cAMP inhibition of RO extract effects on the serotonin receptor
We also compared the efficacy of the distilled water, 20% EtOH, and 80% EtOH RO extracts. We first identified the cAMP inhibition by pretreatment of the RO extract (30 min), followed by 5-HT (15 min), as an agonist for the serotonin receptor, as shown in Figure 7A–C. The effects of treatment with RO extract with 5-HT (15 min) together or the RO extract alone are shown in Figures 7D–F and G–I.
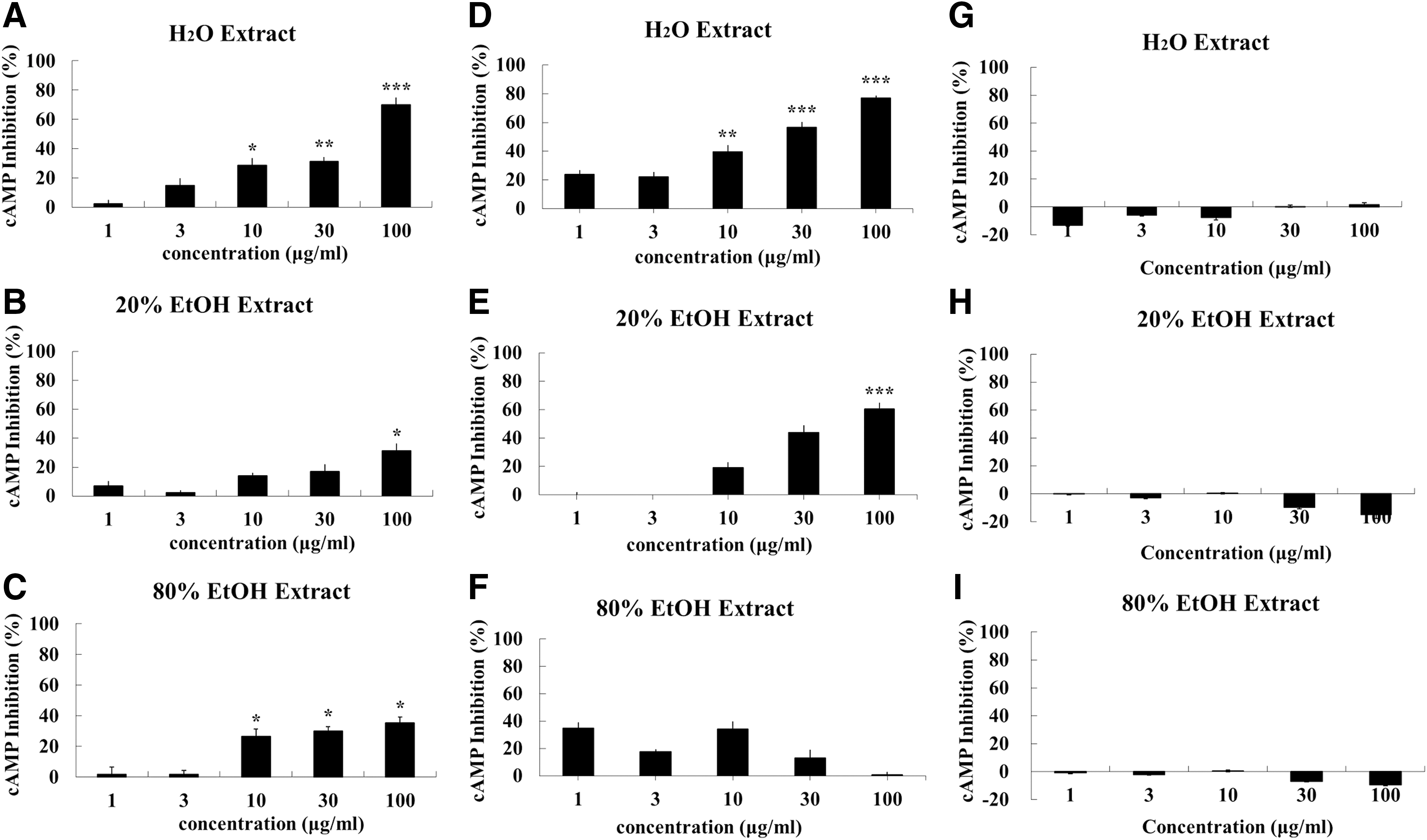
Inhibitory effect of the various fractions of an R. rugosa (RO) extract on 5-HT6 receptor-related cAMP activity in 5-HT6 overexpressing 1321N1 cells.
The RO water extract inhibited the 5-HT6 receptor-related cAMP activity by 2.24% ± 5.24% (1 μg/mL), 14.67% ± 10.11% (3 μg/mL), 28.63% ± 9.92% (10 μg/mL, P < .05), 31.11% ± 5.95% (30 μg/mL, P < .01), and 70.09% ± 10.14% (100 μg/mL, P < .005), as shown in Figure 7A. The 20% EtOH RO extract inhibited this activity by 2.66% ± 6.71% (1 μg/mL), 0.43% ± 3.50% (3 μg/mL), 11.12% ± 3.88% (10 μg/mL), 16.98% ± 10.19% (30 μg/mL), and 31.22% ± 10.22% (100 μg/mL, P < .05), whereas the inhibition by the 80% EtOH RO extract was 1.78% ± 9.17% (1 μg/mL), 1.44% ± 5.42% (3 μg/mL), 33.50% ± 9.49% (10 μg/mL, P < .05), 25.38% ± 5.43% (30 μg/mL, P < .05), and 28.48% ± 7.63% (100 μg/mL, P < .05). Pretreatment with the RO extracts, and especially the water extract, resulted in a dose dependent and statistically significant inhibition of the cAMP activity.
Cotreatment with RO extracts and 5-HT inhibited the cAMP activity by 23.74% ± 6.38% (1 μg/mL), 15.05% ± 8.20% (3 μg/mL), 39.45% ± 9.74% (10 μg/mL, P < .01), 56.55% ± 7.39% (30 μg/mL, P < .005), and 77.00% ± 3.82% (100 μg/mL, P < .005) with the RO water extracts; by 4.32% ± 5.58% (1 μg/mL), −6.92% ± 6.31% (3 μg/mL), 19.14% ± 7.74% (10 μg/mL), 44.11% ± 10.20% (30 μg/mL), and 60.49% ± 9.42% (100 μg/mL, P < .005) with the 20% EtOH RO extract; and by 34.94% ± 8.10% (1 μg/mL), 14.34% ± 3.42% (3 μg/mL), 34.04% ± 11.17% (10 μg/mL), 13.14% ± 11.54% (30 μg/mL), and 0.95% ± 3.99% (100 μg/mL) with the 80% EtOH RO extract (Fig 7D–F). A similar tendency was observed following the pretreatment with the RO extract. The RO extracts themselves did not show any inhibitory effects (Fig. 7G–I)
The water:MeOH column fractions from the distilled water extract of RO were also tested in the same way. Pretreatment with the RO extract inhibited the cAMP activity by 3.39% ± 3.17% (100% H20), 5.98% ± 0.65% (20% MeOH), 10.67% ± 5.57% (50% MeOH, P < .005), 16.09% ± 2.03% (80% MeOH, P < .005), and 0.71% ± 2.37% (100% MeOH), as shown in Figure 8A. The effects of treatment with both RO and 5-HT are shown in Figure 8B, where cAMP activity was inhibited by 16.49% ± 6.09% (100% H20), 5.82% ± 8.23% (20% MeOH), 83.61% ± 16.07% (50% MeOH, P < .01), 62.37% ± 6.48% (80% MeOH, P < .01), and 14.94% ± 7.33% (100% MeOH). The RO extract fractions themselves did not show any inhibitory effects (Fig. 8C).
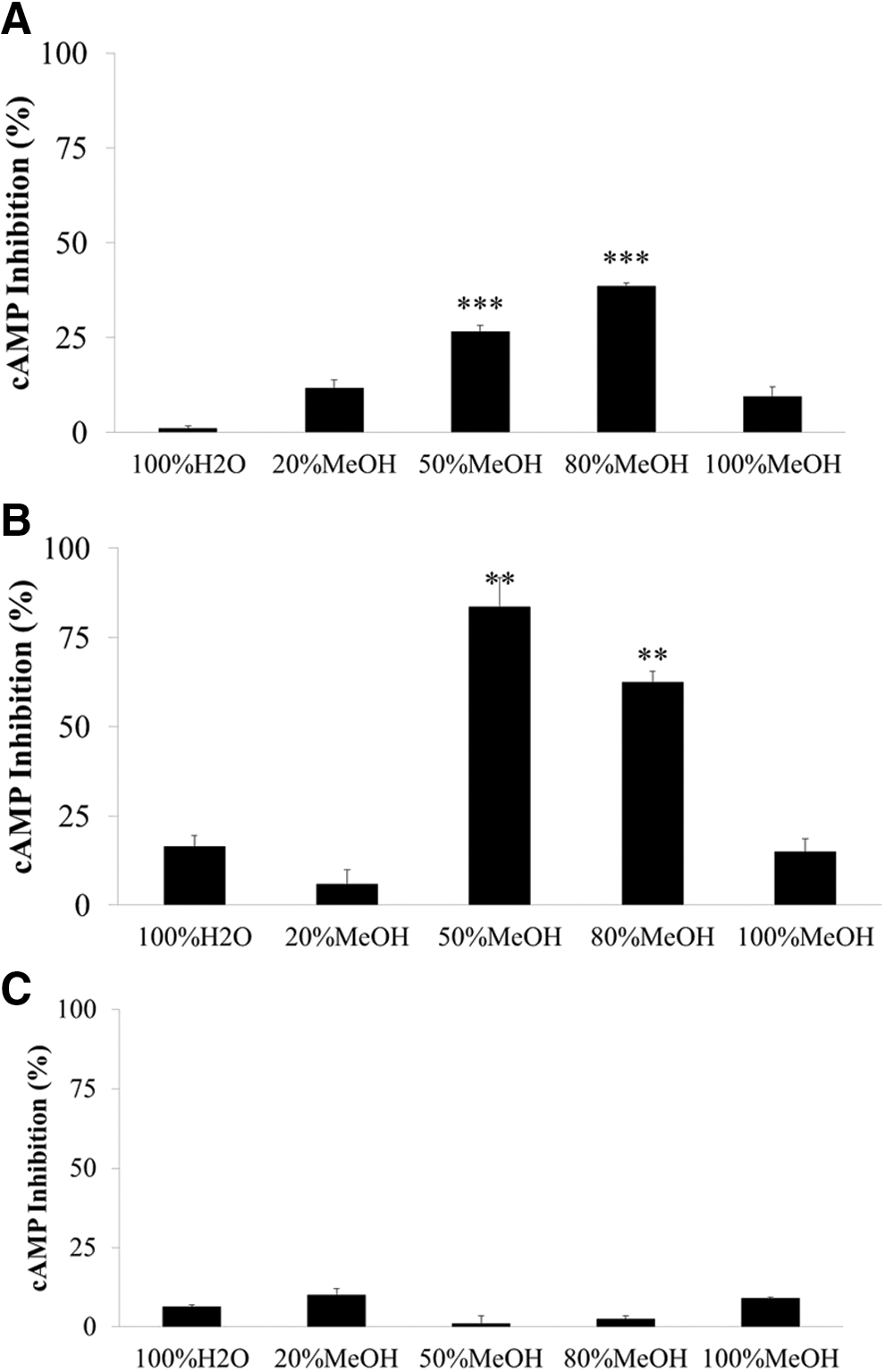
Inhibitory effects of the water:MeOH fractions from an R. rugosa (RO) distilled water extract on 5-HT6 receptor-related cAMP activity in 1321N1 cells.
From these results, we concluded that the 50% and 80% MeOH fractions obtained from the RO distilled water extract have strong inhibitory effects on the cAMP activity.
The activation of MAPK/ERK pathway through serotonin receptor decrease by RO
Based on a related study, 5-HT-induced activation of ERK1/2 is dependent on Ras and MEK in 5-HT6R-transfected HEK293. 40 Therefore, we tried to demonstrate how the RO extract might interact with 5-HT6R through ERK1/2 and MEK. We examined whether 5-HT6R also modulates ERK1/2 activity and, if this is the case, whether RO is involved in 5-HT6R-induced activation of ERK1/2. Figure 9A shows the time course of activation of ERK 1/2 by 5-HT (100 μM) in 5-HT6R-transfected 1321N1 cells. The 5-HT treatment induced ERK 1/2 phosphorylation in a time-dependent manner. The involvement of the RO extract in 5-HT-induced activation of ERK1/2 was further examined by pretreating 5-HT6R-transfected 1321N1 cells with RO extract. A 15-min pretreatment led to a significant decrease in the 5-HT activation of ERK 1/2 (Fig. 9B).

The effects of administration of an R. rugosa (RO) extract after treatment with 5-HT on the activation of ERK 1/2 in 5-HT6 receptor-transfected 132N1 cells.
Mitogen-activated protein kinase/extracellular signal-regulated kinase (MEK) also plays a central role in the classical signaling pathways leading from cell surface receptors to ERK1/2 activation. 41 Figure 10A shows the time-dependent activation of MEK 1/2 by 5-HT in 5-HT6R-transfected 1321N1 cells. A 15-min pretreatment of the 5-HT6R-transfected 1321N1 cells with RO extract significantly reduced the activation of MEK1/2 by 5-HT, as shown in Figure 10B. These results suggested that the 5-HT-induced activation of ERK1/2 was dependent on MEK and that the inhibition of the activation of MEK and ERK1/2 by the RO extract occurred through the 5-HT6 receptor.

Activation of ERK1/2 through 5-HT6receptor depends on MEK.
Discussion
The results of this study demonstrated the novel efficacy of an R. rugosa (RO) extract for improving the stress-related factors caused in vivo by sleep deprivation, and in vitro by H2O2 and CORT, and provided evidence for the involvement of 5-HT6R-mediated signaling pathways. The R. rugosa plant has traditionally been used in folk medicine in East Asian countries, including Korea. In the present study, orally administered RO extract significantly improved physiological (survival, body weight, drowsiness score, and serum cortisol content) and behavioral factors (Morris water maze, rotarod, and passive avoidance test scores) in SD rats.
Caffeine, used in the present study as a positive control (PCTL), is an important pharmacological research tool for studying the involvement of adenosine receptors in cognitive function, 42,43 and it enhances cognitive function in both humans and experimental animals. 44 Caffeine administration alleviates memory impairment in a variety of brain disorders in animal models, including sleep loss. 45 The present results for the Morris water maze test also confirmed that oral administration of caffeine restored the impairment of memory acquisition and retention. The antistress effect of the RO extract was confirmed by the recovery of the physiological, biochemical, and behavioral factors that had been impaired by sleep deprivation stress in vivo and by the concentration-dependent prevention of cell death caused by H2O2 or CORT in vitro. The RO extract was confirmed to act as a 5-HT6 antagonist that inhibited cyclic AMP activity.
Previous studies in various models of learning and memory have shown that 5-HT6 receptor antagonists improve cognitive performance and may be useful in the treatment of cognitive dysfunctions. 46 –48 Serotonergic neurons are more active during waking than during sleep, and extracellular serotonin levels also follow this pattern; therefore, serotonin is thought to stimulate waking. 49 –52 Systemic administration of the selective 5-HT6 receptor antagonists SB-399885 and RO-4368554 during the light phase were reported to increase the awake condition and to reduce slow wave sleep (SWS) and REMS in rats. 53 Accordingly, in vivo microdialysis in the freely moving rat has established that blockade of the 5-HT6 receptor with SB-271046 or RO-046790 increases glutamine (GLU) and acetylcholine (Ach) levels in the frontal cortex and hippocampus and norepinephrine (NE) and dopamine (DA) concentrations in the prelimbic/infralimbic subregions of the medial prefrontal cortex. 54 It remains to be established whether the enhancement of the awake condition and decreases in SWS and REMS after administration of 5-HT6 receptor antagonists are consequences of the increase in ACh, GLU, NE, and DA levels at critical sites in the CNS or if they are independent events.
The 5-HT6 receptors are expressed in the hippocampus and basal ganglia, located on GABAergic neurons. When activated, these receptors increase the inhibitory GABA function and, consequently, decrease cholinergic and/or glutamatergic neuronal activity. Acute treatment with 5-HT6 receptor antagonists improves the consolidation and retention of information by preventing 5-HT6-mediated inhibition of cholinergic function. However, longer term 5-HT6 receptor antagonist treatment also improves acquisition, which could involve dendrite proliferation. 55
We identified the active fractions of the RO extract by column fractionation of a distilled water RO extract and elution by water:MeOH mixtures. We determined the activity of fractions eluted by 50% and 80% MeOH, but further studies are needed.
We demonstrated an interaction between 5-HT6R and the RO extract and the related signaling pathway involving cAMP, ERK1/2, and MEK. Stimulation of Gs-coupled receptors leads to activation of adenylyl cyclase and a rapid increase in cAMP levels. The main target of cAMP is PKA, which plays an essential role in the ERK1/2 cascade. 56 The5-HT4, 5-HT6, and 5-HT7 receptors, which are Gs-coupled receptors that activate ERK1/2, were reported in mammalian cell lines and showed a dependence on Ras and MEK. 40 These observations raise the possibility of an involvement of RO in 5-HT6 function, as well as 5-HT4 and 5-HT7 receptor-mediated activation of ERK1/2.
In conclusion, our results demonstrate that the RO extract prepared in this study exerts an antistress effect by functioning as an antagonist of 5-HT6 receptor by suppressing cyclic AMP activation and subsequent inhibition of MEK and ERK1/2. The decreases in survival and in memory and cognitive function caused by sleep deprivation stress are improved in a concentration-dependent manner by oral administration of the RO extract. This finding suggests that R. rugosa extracts should be considered as a potential treatment for stress.
Footnotes
Acknowledgments
This research was supported by the Research and Development of Regional Industry and funded by the Ministry of Knowledge Economy (MKE) and Korea Institute for Advancement of Technology (KIAT) (70011030), and by the Jeollanamdo Institute for Natural Resources (JINR) Core-Competence Program grants.
Author Disclosure Statement
No competing financial interests exist.