
Abstract
Obesity is viewed as a serious public health problem. This study aimed to investigate the antiobesity effects of fermented garlic extract by lactic acid bacteria (LAFGE) on obesity. Male C57BL/6J mice were fed with high-fat diet (HFD) to induce obesity. The HFD-induced obese mice were orally administrated with 250 or 500 mg/kg LAFGE for 8 weeks. Feeding HFD-fed mice with 250 or 500 mg/kg LAFGE reduced body weight by 14% and 18%, respectively, compared to HFD. HFD-fed mice with 500 mg/kg LAFGE administration had lower epididymal, retroperitoneal, and mesenteric adipose tissue mass by 36%, 44%, and 63%, respectively, compared to HFD. The concentration of plasma triacylglyceride and total cholesterol was significantly lower in the HFD-fed mice with LAFGE administration. Moreover, LAFGE supplementation suppressed adipogenesis by downregulation in mRNA and protein expression of PPARγ, C/EBPα, and lipogenic proteins, including SREBP-1c, FAS, and SCD-1. Based on these findings, LAFGE may ameliorate diet-induced obesity by inhibiting adipose tissue hypertrophy by suppressing adipogenesis.
Introduction
O
Garlic has numerous beneficial effects, including antioxidant, anticancer, antibacterial, cholesterol-lowering, anti-inflammatory, and antidiabetic propterties. 3 –6 Aged black garlic (ABG), one of the well-known processed garlic products, is produced by an aging process at a condition of high temperature and humidity. Although there are numerous advantageous effects of ABG against metabolic diseases, 7 –9 it has the serious disadvantage that the manufacturing process takes more than 30 days. 10 Thus, we wished to develop products with enhanced functional properties compared to raw garlic, using a short-term manufacturing process. One of them is fermented garlic extract with lactic acid bacteria (LAB), however, it was difficult to accomplish since garlic has strong antimicrobial activity from allicin transformed from allin by allinase, when garlic is cut or crushed. 11 Recently, Kim et al. 12 demonstrated that inactivating of allinase by heat processing allows garlic to be fermented with LAB.
Previously, we prepared fermented garlic extract by Lactobacillus plantarum (LAFGE) and it showed an improved garlic-specific pungent taste and aroma. In particular, the manufacturing process of LAFGE requires only 3 days. Previously, we reported that garlic-derived organosulfur compounds (OSC) from LAFGE, including cycloalliin, S-allyl cysteine (SAC), S-methyl cysteine (SMC), and S-ethyl cysteine (SEC), were increased during the fermentation compared to nonfermented garlic extract. Furthermore, consistent with the increases of such OSC, the antioxidant activity of LAFGE was enhanced compared to nonfermented garlic extract. 13 Sumioka et al. 14 demonstrated that lipid-lowering effects of fermented garlic extract were enhanced when using a mold, Monascus pilosus, suggesting that fermented garlic extract with the microorganisms could be a promising functional food supplement for obesity-related disorders such as hyperlipidemia and hypercholesterolemia. However, the antiobesity effects of fermented garlic extract with LAB remain unclear.
In this study, we investigated the ameliorative effect of OSC-enriched LAFGE on a high-fat diet-induced obesity mouse model.
Materials and Methods
Chemicals and reagents
Acrylamide, ammonium persulfate, protease, and phosphatase inhibitor cocktails were purchased from Sigma-Aldrich, Co. (St Louis, MO). SREBP-1c, FAS, SCD-1, C/EBPα, and PPARγ antibodies were purchased from Cell Signaling Technology, Inc. (Boston, MA). Garlic was obtained from Gaweol Agricultural Association in Korea.
Preparation of lactic acid-fermented garlic extract (LIVERTECT™)
Five kilograms of garlic were added to 10 kg of water and homogenized at 100°C for 3 h in the fermenter (MJS U3; Marubishi, Tokyo, Japan). L. Plantarum BL2 (International Depositary Authority: KCCM11019P) was inoculated at 2% (v/v) into homogenized garlic after cooling down to 37°C and then additionally incubated for 36 h. The extract was sterilized for 1 h at 121°C, and then filtered, concentrated, and pulverized.
Animal experiment
C57BL6/N mice (6 weeks old) were obtained from OrientBio (Seongnam, South Korea). Animals were maintained in a controlled environment at 25°C ± 2°C under a 12-h dark/12-h light cycle and acclimated for a week before the experiments. The animals were randomly divided into four groups (n = 8): low-fat diet (LFD) feeding group, high-fat diet (HFD) feeding group, HFD with LAFGE-administered groups (dosages of 250 and 500 mg/kg, respectively). LAFGE suspended in saline was administered orally once a day for 8 weeks. At the end of the experimental period, the animals were sacrificed and the blood was collected by cardiac puncture, and visceral fat pads and liver were removed and weighed. Portions of the epididymal fat pad were immersed in 10% formaldehyde for histological analysis, and the other portions were frozen immediately in liquid nitrogen and stored at −70°C for reverse transcription polymerase chain reaction (RT-PCR) and immunoblot analysis. The animal experiment was approved by the Institutional Animal Care and Use Committee of Korea University (KUIACUC-2016-102).
Serum biochemical assays
The levels of serum alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), triglyceride, and total cholesterol were determined using an automatic biochemical analyzer (Cobas c-111; Roche, Switzerland) according to the instructions supplied with the commercial assay kits (Roche).
Histological analysis
Pieces of fixed epididymal adipose tissue were embedded in paraffin for histological analysis. The embedded tissues were sectioned at 5 μm thickness, stained with hematoxylin and eosin, and examined under a light microscope (Olympus, Tokyo, Japan).
Semiquantitative RT-PCR
The total RNA was isolated using the easy-BLUE™ Total RNA Extraction Kit (Intron Biotechnology, Inc., Seongnam, South Korea) and reverse transcribed to cDNA using a cDNA synthesis kit (Thermo Fisher Scientific, Inc., Waltham, MA) as described in the manufacturer's protocol. For semiquantitative RT-PCR, polymerase chain reaction (PCR) products were separated by electrophoresis on 1.8% agarose gel stained with ethidium bromide. PCR products were visualized under UV light and photographed using the gel documentation system. The intensities of the bands were quantified using ImageJ software (National Institute of Health, NIH Version v1.32j). Relative expression levels of target genes were normalized to β-actin, and then, data were expressed as a ratio compared to control.
Immunoblot analysis
The epididymal tissues were lysed with RIPA buffer (Cell Signaling Technology, Danvers, MA). Tissue lysates were centrifuged at 15,000 g for 20 min. Total protein concentration was determined by Bradford assay. Equal amounts of protein were separated on SDS/PAGE and the proteins were transferred to polyvinylidene difluoride (PVDF) membranes. The membranes were then blocked for 30 min in a PBS solution containing 0.5% BSA and 0.1% Tween-20. After washing, membranes were incubated for 1 h with horseradish peroxidase (HRP)-linked secondary antibodies in PBS containing 0.5% BSA and 0.1% Tween-20 in PBS. Finally, after washing three times for 10 min with PBST, proteins were visualized by ImageQuant LAS 4000 (General Electric, Pittsburgh, PA). Band intensities were quantified with ImageJ software.
Statistical analysis
Statistical significances were analyzed by one-way analysis of variance (ANOVA) followed by Duncan's multiple range test using SAS version 8.2 (SAS Institute, Inc., Cary, NC).
Results
To determine if LAFGE administration can ameliorate HFD-induced obesity, body weight changes were measured. The effects of LAFGE on body weight changes, weight gain, daily food and caloric intake, and food efficiency ratio (FER) during the experimental period are shown in Figure 1. The body weight change for 8 weeks in the HFD-fed group was significantly higher than that of the LFD-fed group (P < .05). LAFGE-supplemented groups lowered the body weight compared to HFD-fed group (Fig. 1A). There were no significant differences in the daily food intake among the mice of all groups (Fig. 1B); however, daily caloric intakes were significantly increased by about two folds in all HFD-fed groups compared to LFD-fed groups (P < .05) (Fig. 1C). Furthermore, the body weight gains during experimental periods were significantly higher in the HFD-fed group (25.6 g) than that in the LFD-fed group (8.3 g) (P < .05); In contrast, the body weight gain in metformin- and LAFGE-supplemented groups at doses of 250 and 500 mg/kg fell to 13.2, 16.5, and 13.8 g, respectively (Fig. 1D). Furthermore, the FER in LAFGE-administered groups at doses of 250 and 500 mg/kg was lower compared to those in HFD (Fig. 1E).
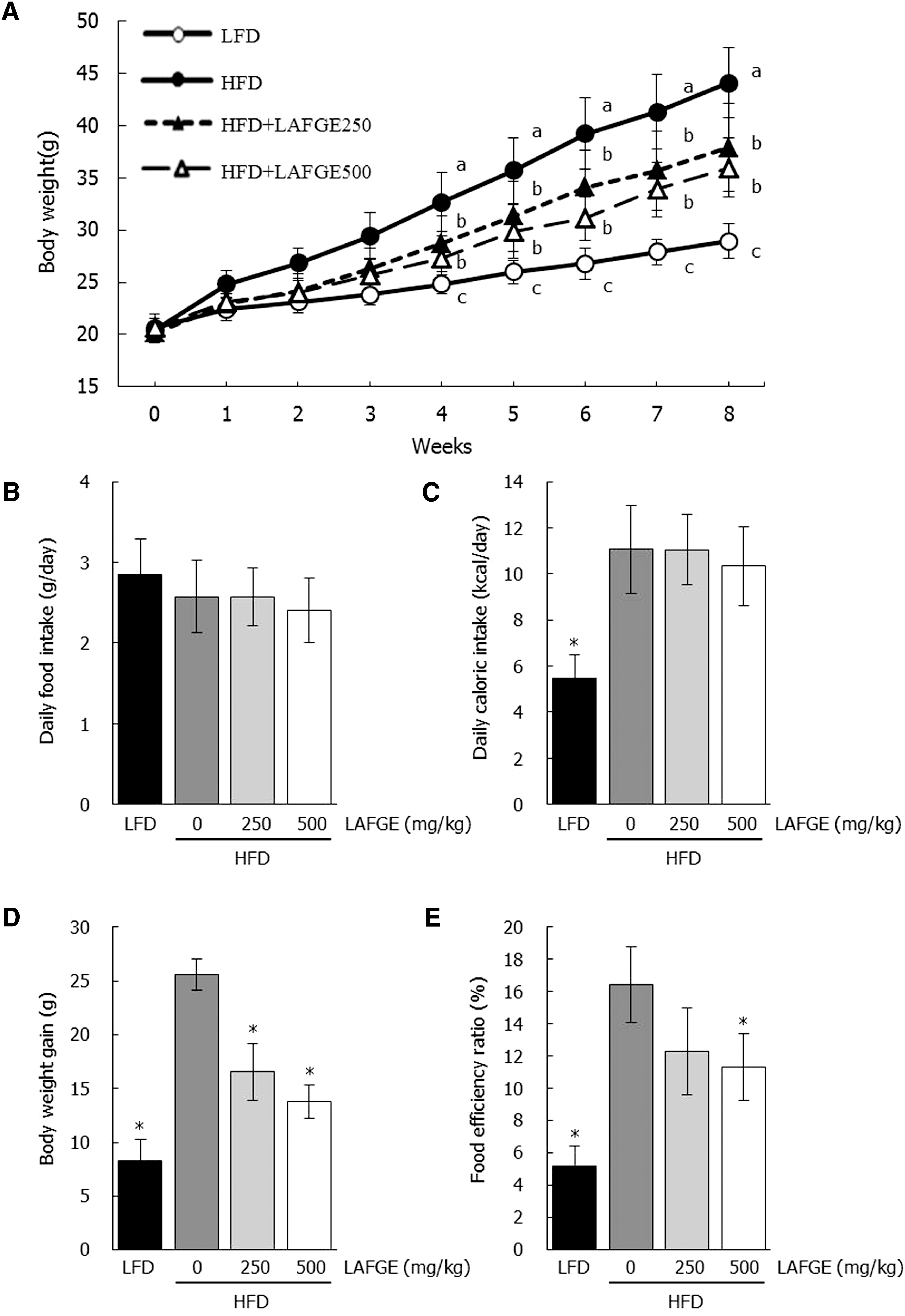
Effects of LAFGE on body weight change
As shown in Figure 2A, the serum triglyceride (TG) levels were significantly increased in the HFD-fed group (72 mg/dL) than in the LFD-fed group (27 mg/dL) (P < .05). However, the serum total TG levels in HFD with LAFGE-administered groups at doses of 250 and 500 mg/kg were significantly decreased to 44 and 37, respectively (P < .05). Serum total cholesterol levels in the HFD-fed group (213 mg/dL) was significantly higher than that of LFD-fed group (115 mg/day). In contrast, serum total cholesterol levels in HFD-administered LAFGE groups at doses of 250 and 500 mg/kg were decreased to 171 and 152 mg/dL, respectively (Fig. 2B).
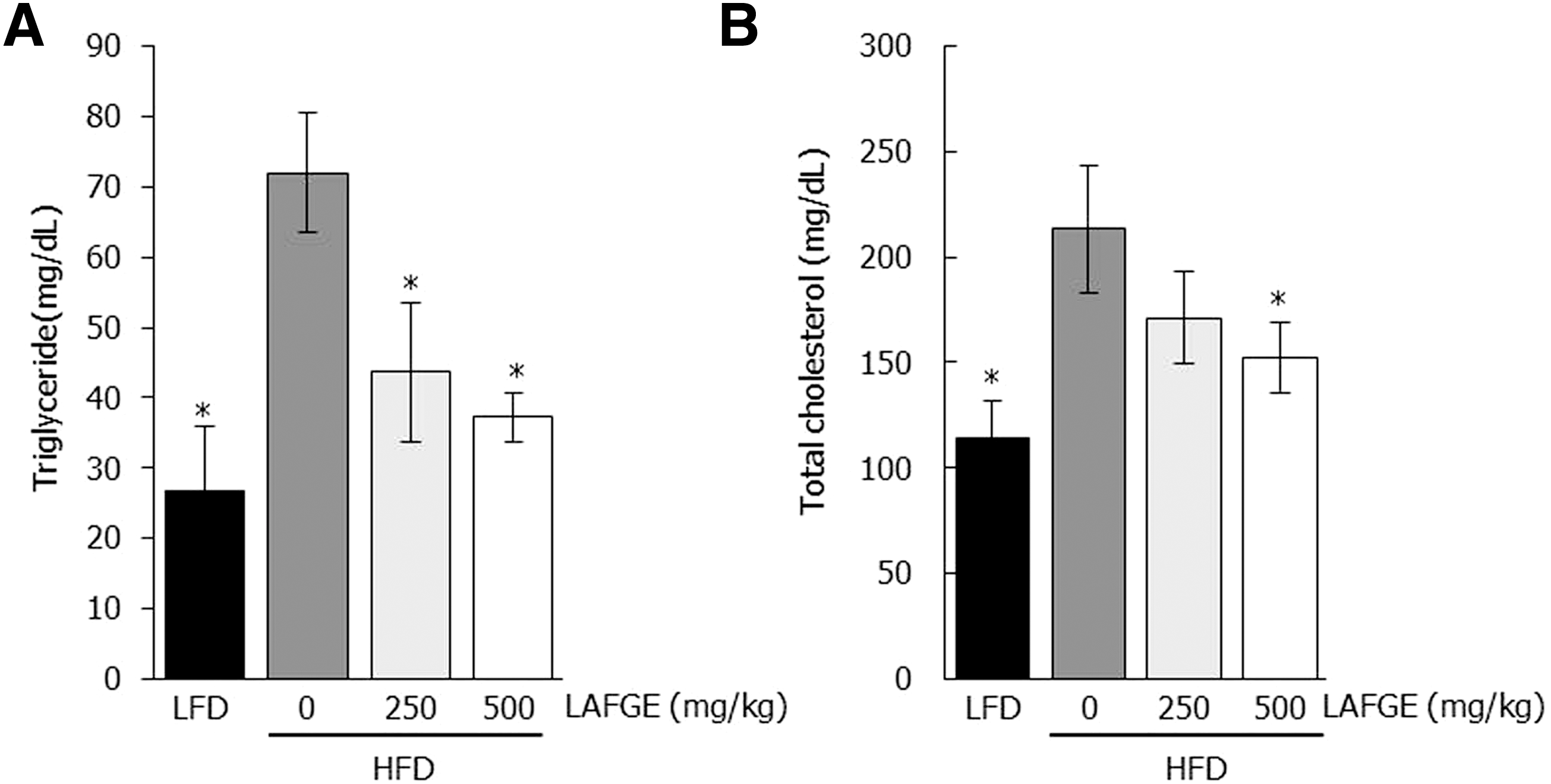
Effects of LAFGE on serum total triglyceride
Next, we measured abdominal fat mass such as epididymal (EAT), retroperitoneal (RAT), and mesenteric adipose tissues (MAT). Such abdominal adipose tissue weights were significantly increased in the HFD group (P < .05), while those of LAFGE-administered groups at doses of 250 and 500 mg/kg were decreased in comparison with the HFD group (Fig. 3A–C). Also the area of epididymal adipocytes in the HFD group was much higher in comparison with that of the LFD group. However, LAFGE-administered groups at a dose of 500 mg/kg had significantly reduced the HFD-induced hypertrophy in epididymal fat cells (P < .05) (Fig. 3D, E).

Effect of LAFGE on adipose tissue of HFD-induced obese mice.
We performed RT-PCR and immunoblot analysis to investigate if the reduction of abdominal adipose tissues in the HFD-fed mice with LAFGE administration related to changes of gene associated with adipogenesis and lipogenesis. As shown in Figure 4, adipogenesis-related factors of epididymal adipose tissue such as peroxisome proliferator-activated receptor γ (PPARγ) and CCAAT-enhancer-binding proteins α (C/EBPα) in HFD-fed group showed significantly higher levels of mRNA and proteins compared to those of LFD-fed group (P < .05). However, LAFGE-administered groups at doses of 250 and 500 mg/kg significantly downregulated both the mRNA and protein levels of PPARγ and C/EBPα compared to those of HFD groups (P < .05) (Fig. 4). Furthermore, the expression of lipogenic proteins such as sterol regulatory element-binding protein 1c (SREBP-1c), fatty acid synthase (FAS), and stearoyl-CoA desaturase 1 (SCD-1) in epididymal adipose tissue revealed higher levels in HFD group than those of LFD group; however, these lipogenic proteins in the HFD-fed group were significantly downregulated compared to those of LAFGE-administered groups at doses of 250 and 500 mg/kg (P < .05) (Fig. 5).
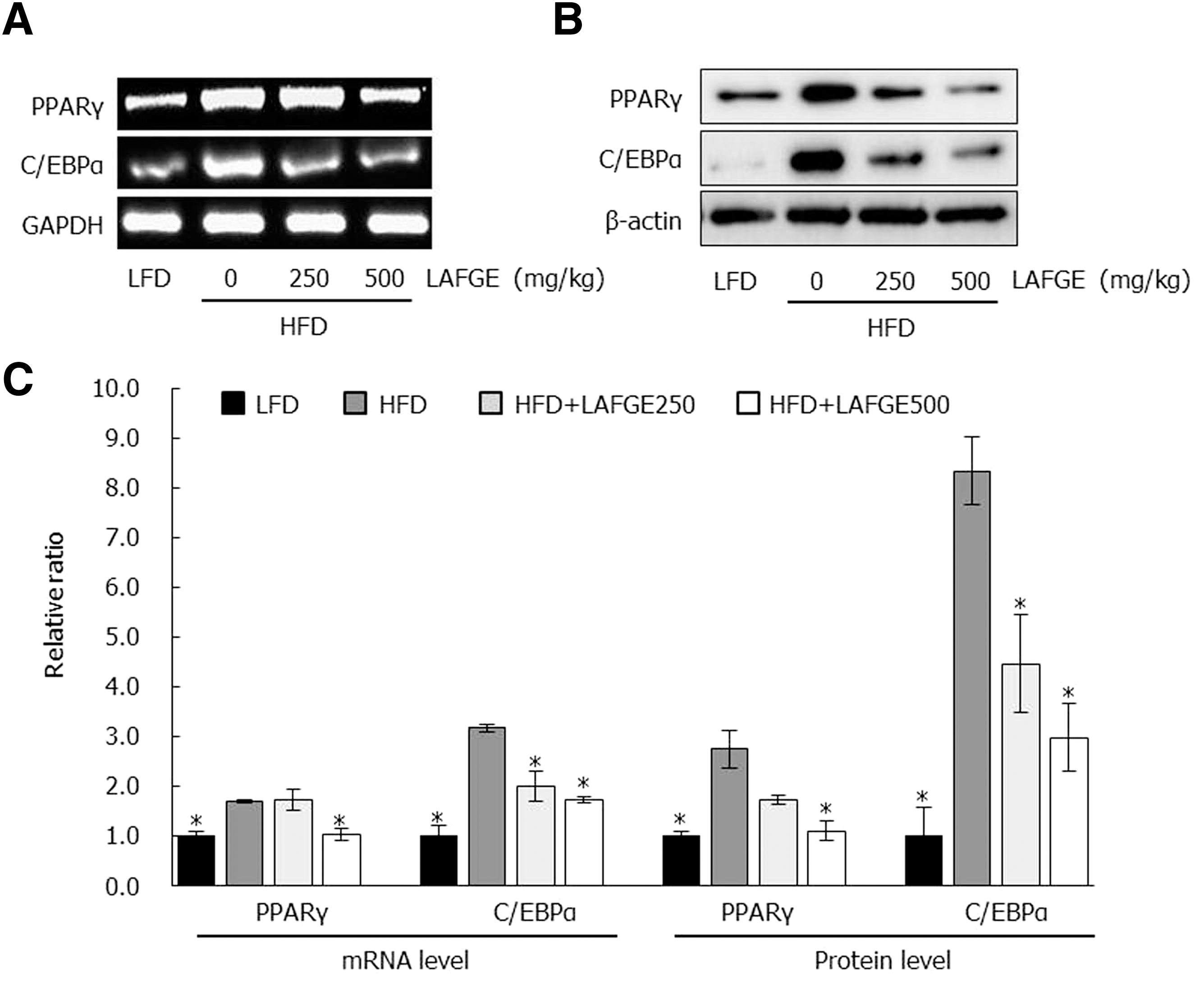
Effect of LAFGE on adipogenic genes and proteins in epididymal adipose tissues. Data are expressed as mean ± SD.
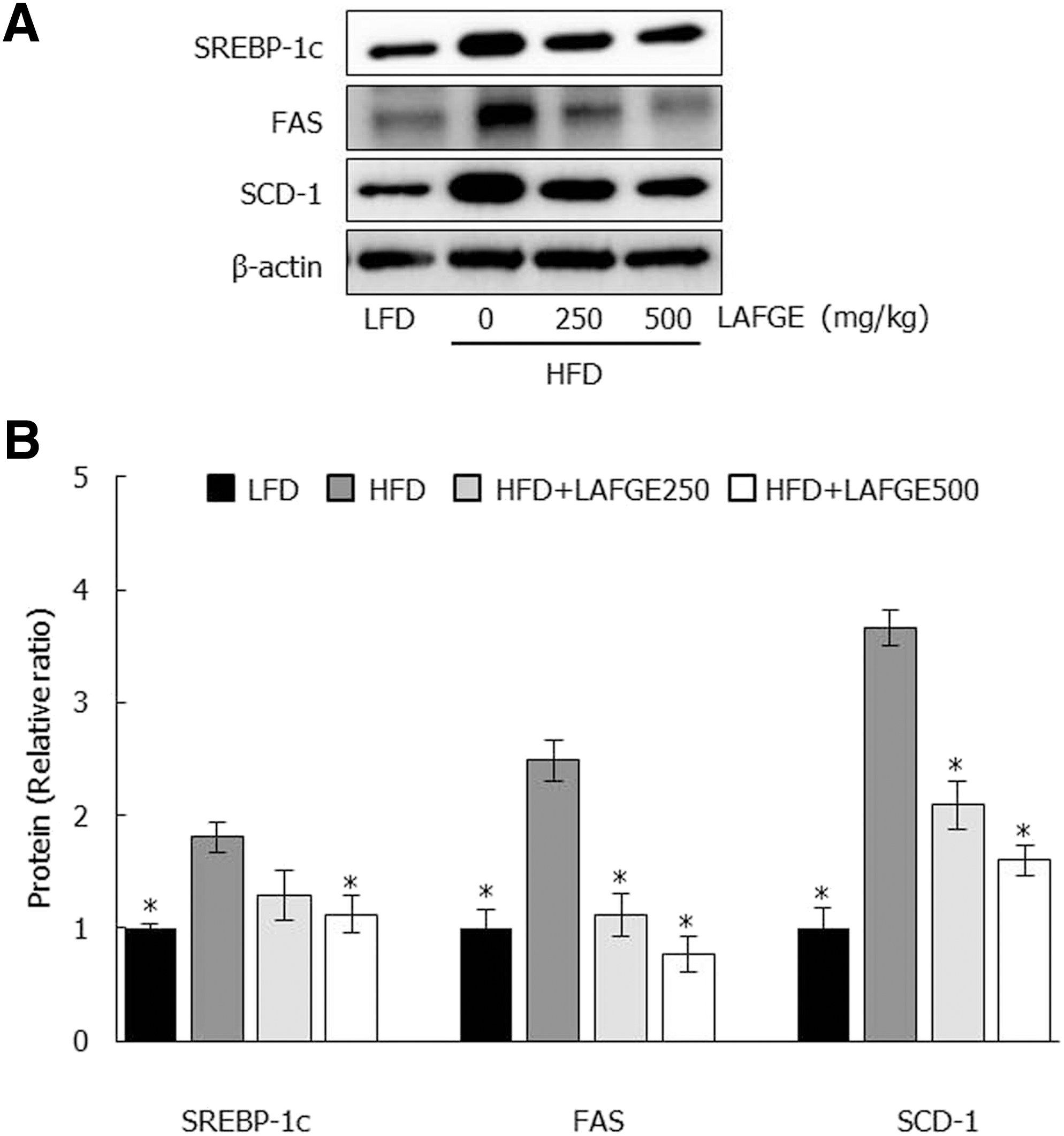
Effect of LAFGE on lipogenic proteins in epididymal adipose tissues. Data are expressed as mean ± SD.
Discussion
Obesity is closely related to excessive caloric intake that exceeds the metabolic needs. Fat accumulation in adipocytes following excess caloric intake is the most representative feature of obesity. 15 In the present study, obese manifestations, including increases of body weight, hyperlipidemia, and adipocyte hypertrophy in HFD-fed mice, were clearly observed. These observations, however, were remarkably attenuated by OSC-enriched LAFGE administration. The body weight gain during the entire experimental period was significantly decreased by LAFGE supplement with no differences in daily food or caloric intake among all the HFD-fed groups, indicating that LAFGE intake effectively reduced body weight gain without anorectic effects.
OSCs, including SAC, SEC, SMC, and s-propyl cysteine (SPC), have shown serum TG- and TC-lowering effects through inhibition of enzymatic activities such as FAS and malic enzyme (ME). 16 SMC and SAC have been revealed to exert a lipid-lowering effect by inhibiting fatty acid and cholesterol synthesis in vitro. 17,18 In the present study, serum TG and total cholesterol levels were markedly increased in HFD-fed mice, while those of LAFGE-administered groups were significantly decreased. These observations could be assumed to be due to the lipid-lowering effects of OSCs, including SAC, SEC, and SMC, which were increased after fermentation with L. plantarum.
Excessive energy is stored in WAT as a form of triglyceride, and adipocyte hypertrophy and hyperplasia are general characteristics in obesity. 19 The increase of visceral fat is a major factor for metabolic diseases, including diabetes, hypertension, and arteriosclerosis. 20 Garlic-derived sulfur-containing compounds such as ajoene and thiacremonone have shown antiobesity capacity through inhibition of adipocyte differentiation and lipid catabolism via AMP-activated protein kinase (AMPK) activation. 21,22 It is unclear whether these compounds were increased by fermentation with L. plantarum; however, HFD-induced hypertrophy of epididymal adipocyte in histological observation and increases of abdominal adipose tissue weights were effectively attenuated by LAFGE supplement, indicating that the antiobesity effect of LAFGE may be related to inhibitory effects of garlic-derived sulfur-containing compounds on adipocyte differentiation.
PPARγ and C/EBPα are considered the important transcription factors in adipocyte differentiation. 23,24 In the late stage of adipogenesis, C/EBPα contributes to promoting the lipogenic process through maintaining PPARγ expression. 25 Activation of PPARγ promotes expression of lipogenic proteins, including SREBP-1c, FAS, and SCD-1, which mediate lipogenesis and lipid accumulation in adipose tissue. 26,27 Thus, PPARγ and C/EBPα were considered critical factors for adipogenesis. Diallyl trisulfide (DATS) and 1,2-vinyldithiin (1,2-DT), which are oil soluble OSCs, have been reported to inhibit differentiation of 3T3-L1 preadipocyte to mature adipocyte by suppression of adipogenesis, including C/EBPs and PPARγ. 28,29 Chen et al. 30 demonstrated that black garlic extract ameliorated diet-induced obesity by downregulation of adipogenesis- and lipogenesis-related genes expression such as PPARγ, SREBP-1c, and FAS in adipose tissues. In the present study, LAFGE effectively suppressed adipogenesis and lipogenesis by downregulation of the abovementioned adipogenic and lipogenic factors. Based on these evidences, it is clear that antiobesity effects of LAFGE in HFD-induced obese mice result from inhibitory activities of garlic-derived OSCs on adipogenesis and lipogenesis. It is considered that these effects of LAFGE play important roles in the prevention of obesity progression.
There are clinical trials in hyperlipidemic subjects regarding fermented garlic extract with Monascus pilosus yeast, which exhibited a lipid-lowering effect, providing availability of fermented garlic extract for preventing obesity-related diseases. 14,31 Moreover, there are several reports regarding the antiobesity effect of intragastrical intake of probiotics such as Lactobacillus gasseri, Lactobacillus acidophilus, Lactobacillus reuteri, and L. plantarum. 32 –35 This body of evidence supports that fermented garlic extract with L. plantarum BL2 has the potential as a safe and effective supplement or functional food to prevent obesity-related disorders.
Taken together, in HFD-induced obese mice, supplementation of LAFGE, which is enriched OSCs by lactic acid fermentation with L. plantarum BL2, suppressed body weight gain, reduced abdominal fat pad, and plasma lipid level via downregulation of adipogenic proteins, including PPARγ, C/EBPα, SREBP-1c, FAS, and SCD-1. These results provide evidence to support its potential for preventing obesity-associated disorders.
Footnotes
Acknowledgment
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry & Fisheries (IPET) through High Value-Added Food Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (no. 112104-03-2-HD020).
Author Disclosure Statement
No competing financial interests exist.